The Proposed Neurotoxin β-N-Methylamino-l-Alanine (BMAA) Is Taken up through Amino-Acid Transport Systems in the Cyanobacterium Anabaena PCC 7120

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Anabaena Is a Suitable Model for Genetic Studies of BMAA Metabolism
2.2. Isolation of BMAA Resistant Mutants
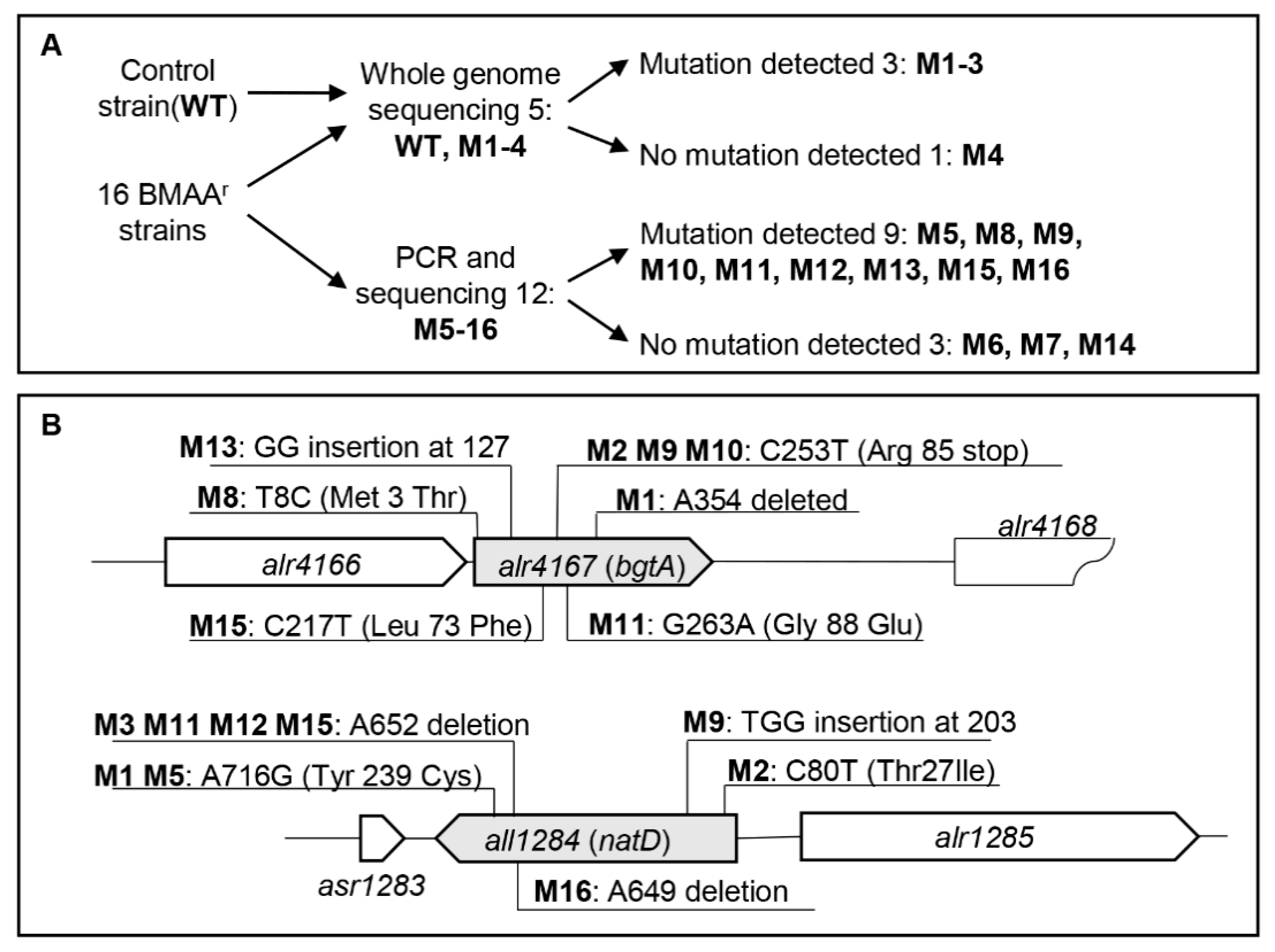
2.3. Spontaneous Mutations Conferring Resistance to BMAA in Anabaena Were Mapped on Genes Involved in Amino Acid Transport
2.4. Exogenously Supplied Amino Acids in the Growth Medium Relieve the Toxic Effect of BMAA in Anabaena
2.5. Inactivation of the Three Major Amino Acid Transport Tystems Conferred Full Resistance to BMAA
2.6. BMAA Uptake Assay in the Wild Type and the Mutants
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. BMAA, Other Chemicals and Molecular Biology Tools
4.3. Screening of Spontaneous BMAA Resistance Mutants
4.4. Construction of Plasmids, Mutants and Genomic Sequencing
4.5. Sample Preparation for BMAA Detection
4.6. BMAA Derivatization
4.7. UPLC-MS/MS
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vega, A. α-Amino-β-methylaminopropionic acid, a new amino acid from seeds of Cycas circinalis. Phytochemistry 1967, 6, 759–762. [Google Scholar] [CrossRef]
- Nunn, P.B.; Seelig, M.; Zagoren, J.C.; Spencer, P.S. Stereospecific acute neuronotoxicity of ‘uncommon’ plant amino acids linked to human motor-system diseases. Brain Res. 1987, 410, 375–379. [Google Scholar] [CrossRef]
- Delcourt, N.; Claudepierre, T.; Maignien, T.; Arnich, N.; Mattei, C. Cellular and molecular aspects of the β-N-Methylamino-l-alanine (BMAA) mode of action within the neurodegenerative pathway: Facts and controversy. Toxins 2017, 10, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, P.A.; Davis, D.A.; Mash, D.C.; Metcalf, J.S.; Banack, S.A. Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain. Proc. R. Soc. B Boil. Sci. 2016, 283, 20152397. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Mondo, K.; Stern, E.; Annor, A.K.; Murch, S.J.; Coyne, T.M.; Brand, L.E.; Niemeyer, M.E.; Sharp, S.; Bradley, W.G.; et al. Cyanobacterial neurotoxin BMAA and brain pathology in stranded dolphins. PLoS ONE 2019, 14, e0213346. [Google Scholar] [CrossRef]
- Eiser, A.R. Why does Finland have the highest dementia mortality rate? Environmental factors may be generalizable. Brain Res. 2017, 1671, 14–17. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β-N-methylamino-l-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, T.; Kato, N.; Hosoya, K.; Kaya, K. Effective determination method for a cyanobacterial neurotoxin, β-N-methylamino-l-alanine. Toxicon 2008, 51, 1264–1268. [Google Scholar] [CrossRef]
- Moura, S.; Ultramari, M.D.A.; De Paula, D.M.L.; Yonamine, M.; Pinto, E. 1H NMR determination of β-N-methylamino-l-alanine (l-BMAA) in environmental and biological samples. Toxicon 2009, 53, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Krüger, T.; Mönch, B.; Oppenhäuser, S.; Luckas, B. LC–MS/MS determination of the isomeric neurotoxins BMAA (β-N-methylamino-l-alanine) and DAB (2,4-diaminobutyric acid) in cyanobacteria and seeds of Cycas revoluta and Lathyrus latifolius. Toxicon 2010, 55, 547–557. [Google Scholar] [CrossRef]
- Li, A.; Fan, H.; Ma, F.; McCarron, P.; Thomas, K.; Tang, X.; Quilliam, M. Elucidation of matrix effects and performance of solid-phase extraction for LC-MS/MS analysis of β-N-methylamino-l-alanine (BMAA) and 2,4-diaminobutyric acid (DAB) neurotoxins in cyanobacteria. Analyst 2012, 137, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Réveillon, D.; Abadie, E.; Sechet, V.; Brient, L.; Savar, V.; Bardouil, M.; Hess, P.; Amzil, Z. Beta-N-methylamino-l-alanine: LC-MS/MS optimization, screening of cyanobacterial strains and occurrence in shellfish from Thau, a French Mediterranean lagoon. Mar. Drugs 2014, 12, 5441–5467. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Qiu, J.; Fan, L.; Li, A. Effects of growth conditions on the production of neurotoxin 2,4-diaminobutyric acid (DAB) in Microcystis aeruginosa and its universal presence in diverse cyanobacteria isolated from freshwater in China. Environ. Sci. Pollut. Res. 2014, 22, 5943–5951. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef]
- Quiblier, C.; Susanna, W.; Isidora, E.-S.; Mark, H.; Aurélie, V.; Jean-François, H. A review of current knowledge on toxic benthic freshwater cyanobacteria—Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J. Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the Chamorro people of Guam. Proc. Natl. Acad. Sci. USA 2003, 100, 13380–13383. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, K.J.; Main, B.J.; Samardzic, K. Cyanobacterial neurotoxins: Their occurrence and mechanisms of toxicity. Neurotox. Res. 2017, 33, 168–177. [Google Scholar] [CrossRef]
- Downing, S.; Downing, T.G. The metabolism of the non-proteinogenic amino acid β-N-methylamino-l-alanine (BMAA) in the cyanobacterium Synechocystis PCC6803. Toxicon 2016, 115, 41–48. [Google Scholar] [CrossRef]
- Nunn, P.B.; Codd, G.A. Metabolic solutions to the biosynthesis of some diaminomonocarboxylic acids in nature: Formation in cyanobacteria of the neurotoxins 3-N-methyl-2,3-diaminopropanoic acid (BMAA) and 2,4-diaminobutanoic acid (2,4-DAB). Phytochemistry 2017, 144, 253–270. [Google Scholar] [CrossRef]
- Downing, S.; Banack, S.; Metcalf, J.S.; Cox, P.A.; Downing, T. Nitrogen starvation of cyanobacteria results in the production of β-N-methylamino-l-alanine. Toxicon 2011, 58, 187–194. [Google Scholar] [CrossRef]
- Yan, B.; Liu, Z.; Huang, R.; Xu, Y.; Liu, D.; Wang, W.; Zhao, Z.; Cui, F.; Shi, W. Impact factors on the production of β-methylamino-l-alanine (BMAA) by cyanobacteria. Chemosphere 2020, 243, 125355. [Google Scholar] [CrossRef] [PubMed]
- Downing, S.; Van De Venter, M.; Downing, T.G. The effect of exogenous β-N-Methylamino-l-alanine on the growth of Synechocystis PCC6803. Microb. Ecol. 2011, 63, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Berntzon, L.; Erasmie, S.; Celepli, N.A.; Eriksson, J.; Rasmussen, U.; Bergman, B. BMAA inhibits nitrogen fixation in the cyanobacterium Nostoc sp. PCC 7120. Mar. Drugs 2013, 11, 3091–3108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolk, C.P. Genetic analysis of cyanobacterial development. Curr. Opin. Genet. Dev. 1991, 1, 336–341. [Google Scholar] [CrossRef]
- Grigorieva, G.; Shestakov, S. Transformation in the cyanobacterium Synechocystis sp. 6803. FEMS Microbiol. Lett. 1982, 13, 367–370. [Google Scholar] [CrossRef]
- Pernil, R.; Picossi, S.; Herrero, A.; Flores, E.; Mariscal, V. Amino acid transporters and release of hydrophobic amino acids in the heterocyst-forming cyanobacterium Anabaena sp. strain PCC 7120. Life 2015, 5, 1282–1300. [Google Scholar] [CrossRef] [Green Version]
- Pernil, R.; Picossi, S.; Mariscal, V.; Herrero, A.; Flores, E. ABC-type amino acid uptake transporters Bgt and N-II of Anabaena sp. strain PCC 7120 share an ATPase subunit and are expressed in vegetative cells and heterocysts. Mol. Microbiol. 2008, 67, 1067–1080. [Google Scholar] [CrossRef]
- Zhang, C.-C.; Laurent, S.; Sakr, S.; Peng, L.; Bédu, S. Heterocyst differentiation and pattern formation in cyanobacteria: A chorus of signals. Mol. Microbiol. 2005, 59, 367–375. [Google Scholar] [CrossRef]
- Picossi, S.; Montesinos, M.L.; Pernil, R.; Lichtlé, C.; Herrero, A.; Flores, E. ABC-type neutral amino acid permease N-I is required for optimal diazotrophic growth and is repressed in the heterocysts of Anabaena sp. strain PCC 7120. Mol. Microbiol. 2005, 57, 1582–1592. [Google Scholar] [CrossRef]
- Ungerer, J.; Pakrasi, H.B. Cpf1 is a versatile tool for CRISPR genome editing across diverse species of cyanobacteria. Sci. Rep. 2016, 6, 39681. [Google Scholar] [CrossRef]
- Niu, T.-C.; Lin, G.-M.; Xie, L.-R.; Wang, Z.-Q.; Xing, W.-Y.; Zhang, J.-Y.; Zhang, C.-C. Expanding the potential of CRISPR-Cpf1-based genome editing technology in the Cyanobacterium Anabaena PCC 7120. ACS Synth. Boil. 2018, 8, 170–180. [Google Scholar] [CrossRef]
- Bishop, S.L.; Murch, S.J. A systematic review of analytical methods for the detection and quantification of β-N-methylamino-l-alanine (BMAA). Analyst 2020, 145, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Baker, T.C.; Tymm, F.J.; Murch, S.J. Assessing environmental exposure to β-N-Methylamino-l-Alanine (BMAA) in complex sample matrices: A comparison of the three most popular LC-MS/MS methods. Neurotox. Res. 2017, 33, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Manolidi, K.; Triantis, T.M.; Kaloudis, T.; Hiskia, A. Neurotoxin BMAA and its isomeric amino acids in cyanobacteria and cyanobacteria-based food supplements. J. Hazard. Mater. 2019, 365, 346–365. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, Y.; Li, C.; Gao, H.; Zhang, C.-C.; Xu, X. Three substrains of the Cyanobacterium Anabaena sp. strain PCC 7120 display divergence in genomic sequences and hetC function. J. Bacteriol. 2018, 200, e00076-18. [Google Scholar] [CrossRef] [Green Version]
- Popova, A.A.; Semashko, T.A.; Kostina, N.V.; Rasmussen, U.; Govorun, V.; Koksharova, O.A. The cyanotoxin BMAA induces heterocyst specific gene expression in Anabaena sp. PCC 7120 under repressive conditions. Toxins 2018, 10, 478. [Google Scholar] [CrossRef] [Green Version]
- Herrero, A.; Stavans, J.; Flores, E. The multicellular nature of filamentous heterocyst-forming cyanobacteria. FEMS Microbiol. Rev. 2016, 40, 831–854. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.; Karlsson, O.; Bergström, U.; Brittebo, E.B.; Brandt, I. Maternal transfer of the cyanobacterial neurotoxin β-N-Methylamino-l-Alanine (BMAA) via milk to suckling offspring. PLoS ONE 2013, 8, e78133. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.; Ersson, L.; Brandt, I.; Bergström, U. Potential transfer of neurotoxic amino acid β-N-methylamino-alanine (BMAA) from mother to infant during breast-feeding: Predictions from human cell lines. Toxicol. Appl. Pharmacol. 2017, 320, 40–50. [Google Scholar] [CrossRef]
- Zhang, C.C. A gene encoding a protein related to eukaryotic protein kinases from the filamentous heterocystous Cyanobacterium Anabaena PCC 7120. Proc. Natl. Acad. Sci. USA 1993, 90, 11840–11844. [Google Scholar] [CrossRef] [Green Version]
- Rippka, R. Isolation and purification of cyanobacteria. Methods Enzymol. 1988, 167, 3–27. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.J.; Gillissen, F.; Lürling, M. A Comparative study on three analytical methods for the determination of the neurotoxin BMAA in Cyanobacteria. PLoS ONE 2012, 7, e36667. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.-Q.; Wang, S.; Zhang, J.-Y.; Lin, G.-M.; Gan, N.; Song, L.; Zeng, X.; Zhang, C.-C. The Proposed Neurotoxin β-N-Methylamino-l-Alanine (BMAA) Is Taken up through Amino-Acid Transport Systems in the Cyanobacterium Anabaena PCC 7120. Toxins 2020, 12, 518. https://doi.org/10.3390/toxins12080518
Wang Z-Q, Wang S, Zhang J-Y, Lin G-M, Gan N, Song L, Zeng X, Zhang C-C. The Proposed Neurotoxin β-N-Methylamino-l-Alanine (BMAA) Is Taken up through Amino-Acid Transport Systems in the Cyanobacterium Anabaena PCC 7120. Toxins. 2020; 12(8):518. https://doi.org/10.3390/toxins12080518
Chicago/Turabian StyleWang, Zi-Qian, Suqin Wang, Ju-Yuan Zhang, Gui-Ming Lin, Nanqin Gan, Lirong Song, Xiaoli Zeng, and Cheng-Cai Zhang. 2020. "The Proposed Neurotoxin β-N-Methylamino-l-Alanine (BMAA) Is Taken up through Amino-Acid Transport Systems in the Cyanobacterium Anabaena PCC 7120" Toxins 12, no. 8: 518. https://doi.org/10.3390/toxins12080518