Acetamiprid Affects Destruxins Production but Its Accumulation in Metarhizium sp. Spores Increases Infection Ability of Fungi


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
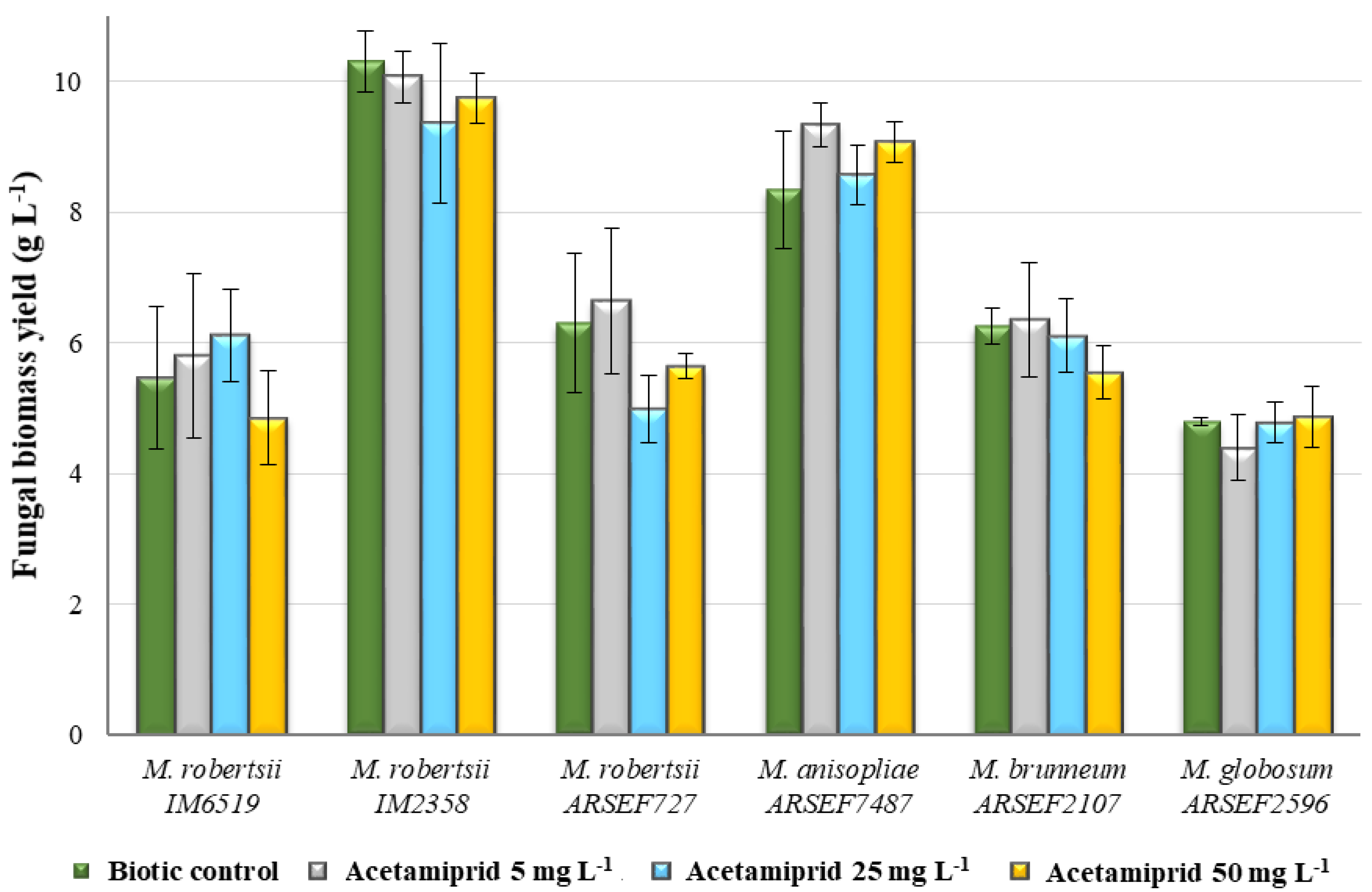
2.1. Fungal Biomass Yield of Metarhizium sp. in the Presence of Acetamiprid
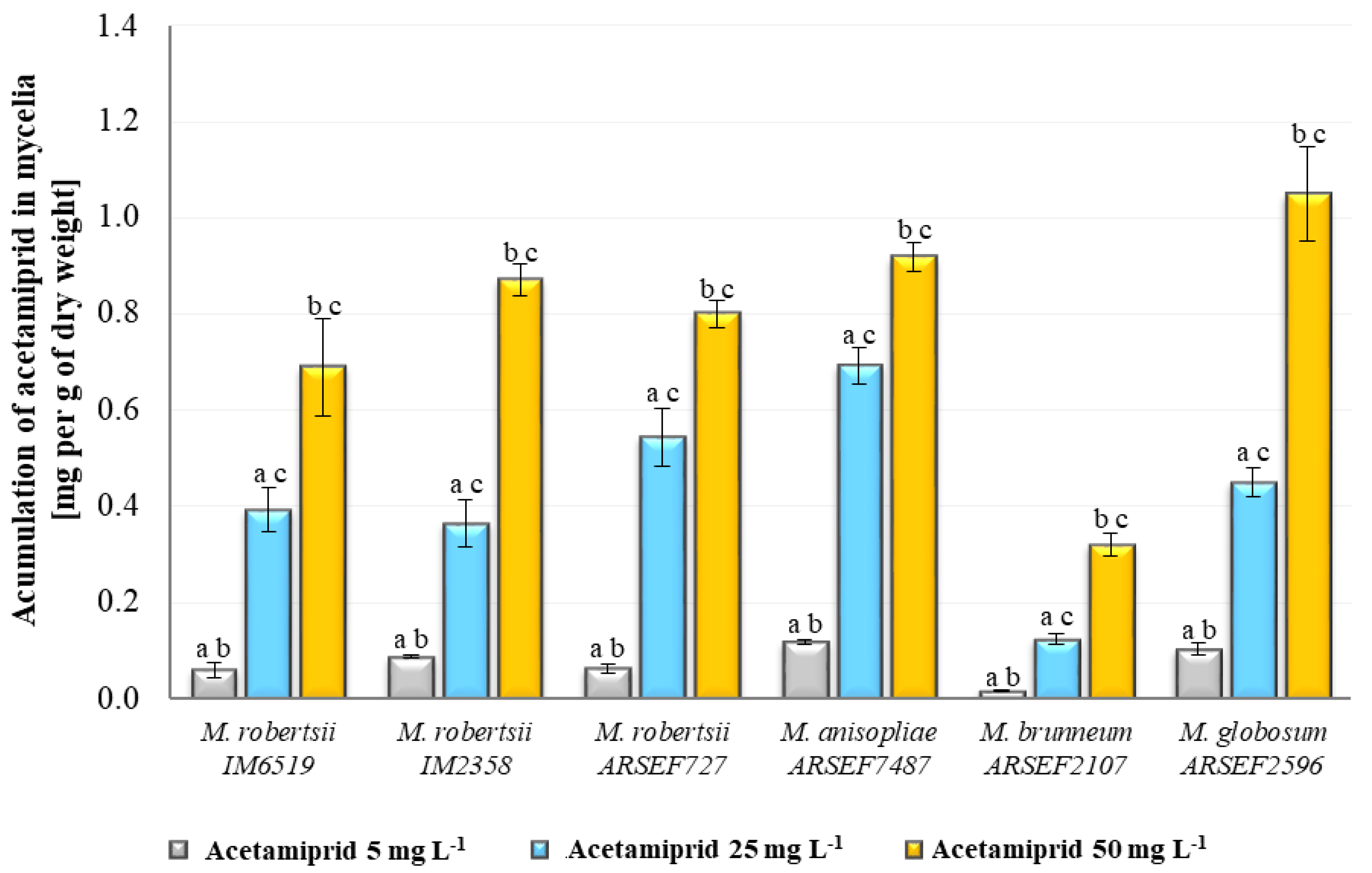
2.2. Quantitative Analyses of the Content of Acetamiprid in Metarhizium Fungal Cultures
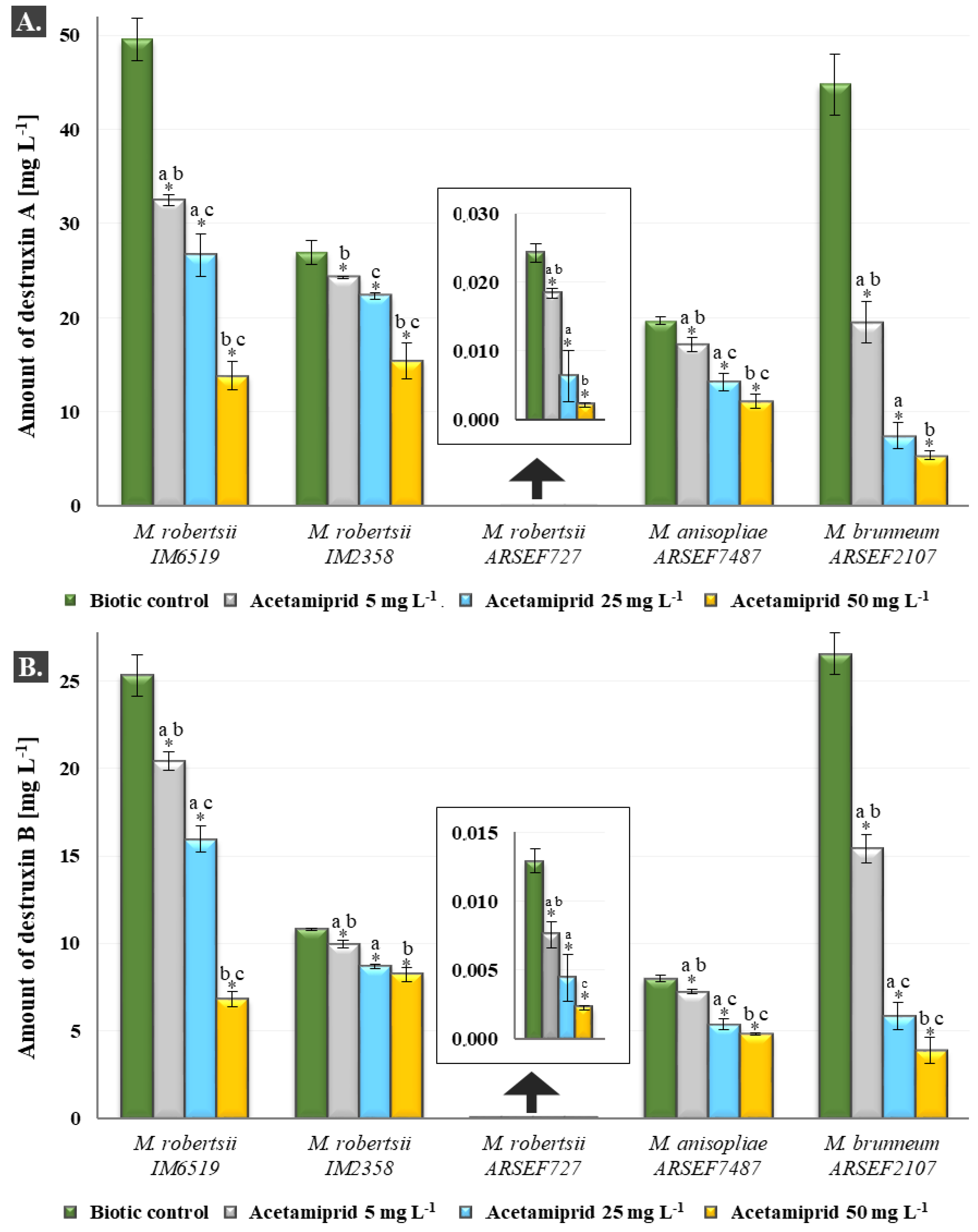
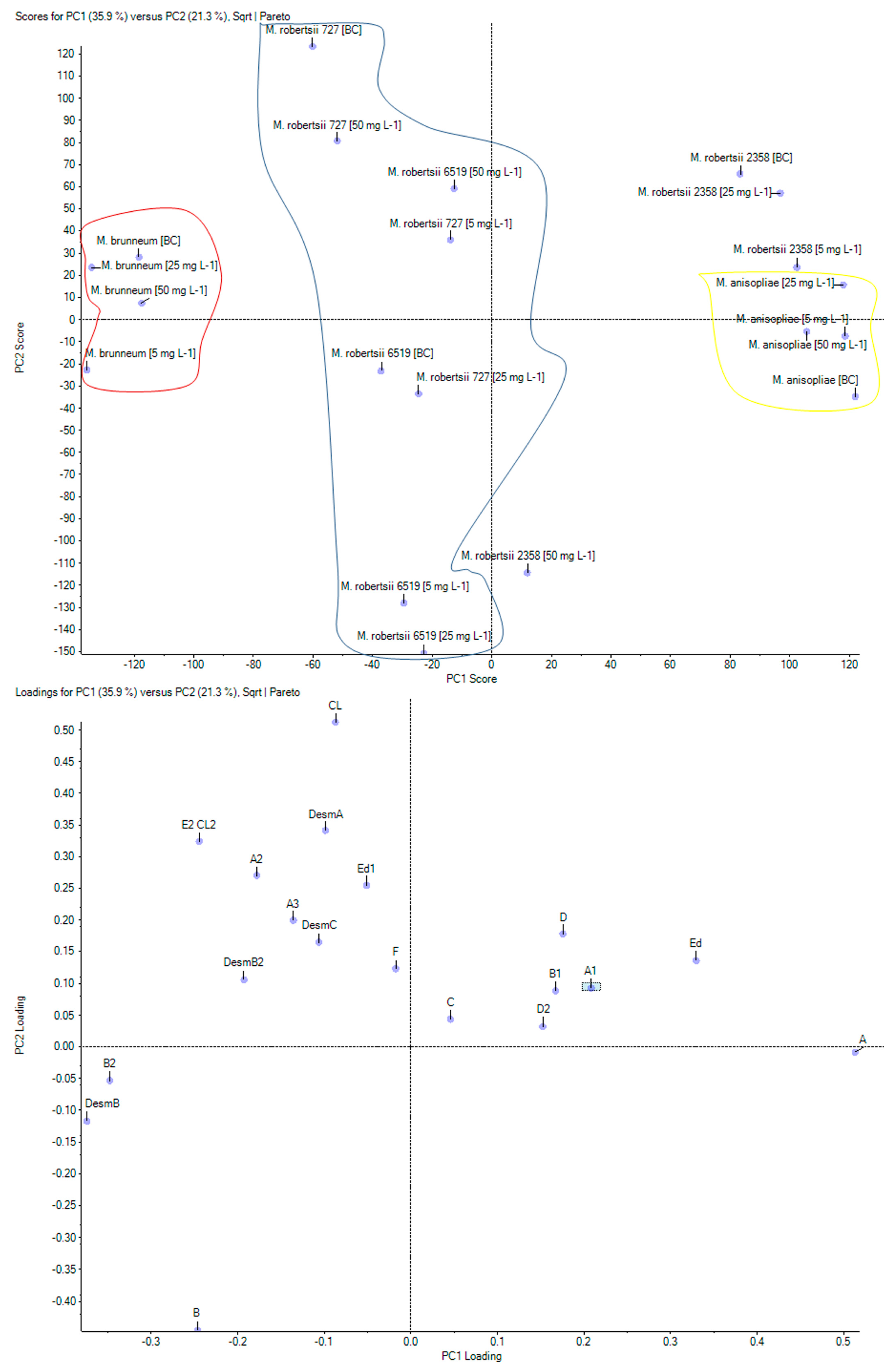
2.3. Analyses of Destruxins in Fungal Cultures of Metarhizium sp.
2.4. Permeability of the Cell Membrane and the Content of Acetamiprid in Spores and Subcellular Fractions of the M. brunneum
2.5. Influence of Acetamiprid, Spores of M. brunneum ARSEF2107 and the Combination of Spores and Acetamiprid on the Mortality of Tenebrio molitor (Mealworm)
3. Conclusions
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Microorganisms and Cultivation Conditions
4.3. Destruxins Extraction by a Modified QuEChERS Method
4.4. Acetamiprid Extraction by a Modified QuEChERS Method
4.5. Acetamiprid Extraction from M. brunneum Spores and Subcellular Fractions
4.6. LC–MS/MS Quantitative and Qualitative Analyses of Destruxins
4.7. LC–MS/MS Quantitative Analyses of Acetamiprid
4.8. Permeability of the Cell Membranes
4.9. Mortality Test of Larvae of Tenebrio molitor (Mealworm)
4.10. Data Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Han, W.; Tian, Y.; Shen, X. Human exposure to neonicotinoid insecticides and the evaluation of their potential toxicity: An overview. Chemosphere 2018, 192, 59–65. [Google Scholar] [CrossRef]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority). Conclusion on the peer review of the pesticide risk assessment of the active substance acetamiprid. EFSA J. 2016, 14, 26. [Google Scholar] [CrossRef]
- Xiong, J.; Wang, Z.; Ma, X.; Li, H.; You, J. Occurrence and risk of neonicotinoid insecticides in surface water in a rapidly developing region: Application of polar organic chemical integrative samplers. Sci. Total Environ. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ihara, M. Neonicotinoids: Molecular mechanisms of action, insights into resistance and impact on pollinators. Curr. Opin. Insect Sci. 2018, 30, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Lovett, B.; St. Leger, R.J. Genetically engineering better fungal biopesticides. Pest Manag. Sci. 2018, 74, 781–789. [Google Scholar] [CrossRef]
- Mondal, S.; Baksi, S.; Koris, A.; Vatai, G. Journey of enzymes in entomopathogenic fungi. Pac. Sci. Rev. A Nat. Sci. Eng. 2016. [Google Scholar] [CrossRef]
- Wang, B.; Kang, Q.; Lu, Y.; Bai, L.; Wang, C. Unveiling the biosynthetic puzzle of destruxins in Metarhizium species. Proc. Natl. Acad. Sci. USA 2012, 109, 1287–1292. [Google Scholar] [CrossRef]
- Donzelli, B.G.G.; Krasnoff, S.B. Molecular Genetics of Secondary Chemistry in Metarhizium Fungi. Adv. Genet. 2016. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Irina Zaharia, L.I.; Ward, D.E. The destruxins: Synthesis, biosynthesis, biotransformation, and biological activity. Phytochemistry 2002, 59, 579–596. [Google Scholar] [CrossRef]
- Wang, X.; Gong, X.; Li, P.; Lai, D.; Zhou, L.; Wang, X.; Gong, X.; Li, P.; Lai, D.; Zhou, L. Structural diversity and biological activities of cyclic depsipeptides from fungi. Molecules 2018, 23, 169. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.L.; Tzeng, Y.M. Development and applications of destruxins: A review. Biotechnol. Adv. 2012, 30, 1242–1254. [Google Scholar] [CrossRef] [PubMed]
- Litwin, A.; Nowak, M.; Różalska, S. Entomopathogenic fungi: Unconventional applications. Rev. Environ. Sci. Biotechnol. 2020, 19, 23–42. [Google Scholar] [CrossRef]
- Ríos-Moreno, A.; Garrido-Jurado, I.; Resquín-Romero, G.; Arroyo-Manzanares, N.; Arce, L.; Quesada-Moraga, E. Destruxin A production by Metarhizium brunneum strains during transient endophytic colonisation of Solanum tuberosum. Biocontrol Sci. Technol. 2016, 26, 1574–1585. [Google Scholar] [CrossRef]
- Taibon, J.; Sturm, S.; Seger, C.; Strasser, H.; Stuppner, H. Quantitative assessment of destruxins from strawberry and maize in the lower parts per billion range: Combination of a QuEChERS-based extraction protocol with a fast and selective UHPLC-QTOF-MS assay. J. Agric. Food Chem. 2015, 63, 5707–5713. [Google Scholar] [CrossRef]
- Neves, P.M.O.J.; Hirose, E.; Tchujo, P.T.; Moino, J.R.A. Compatibility of entomopathogenic fungi with neonicotinoid insecticides. Neotrop. Entomol. 2001, 30, 263–268. [Google Scholar] [CrossRef]
- Li, Y.; Long, L.; Yan, H.; Ge, J.; Cheng, J.; Ren, L.; Yu, X. Comparison of uptake, translocation and accumulation of several neonicotinoids in komatsuna (Brassica rapa var. perviridis) from contaminated soils. Chemosphere 2018, 200, 603–611. [Google Scholar] [CrossRef] [PubMed]
- De Laet, C.; Matringe, T.; Petit, E.; Grison, C. Eichhornia crassipes: A Powerful Bio-indicator for Water Pollution by Emerging Pollutants. Sci. Rep. 2019, 9, 7326. [Google Scholar] [CrossRef]
- Barbieri, M.V.; Postigo, C.; Guillem-Argiles, N.; Monllor-Alcaraz, L.S.; Simionato, J.I.; Stella, E.; Barceló, D.; López de Alda, M. Analysis of 52 pesticides in fresh fish muscle by QuEChERS extraction followed by LC-MS/MS determination. Sci. Total Environ. 2019, 653, 958–967. [Google Scholar] [CrossRef]
- Bartlett, A.J.; Hedges, A.M.; Intini, K.D.; Brown, L.R.; Maisonneuve, F.J.; Robinson, S.A.; Gillis, P.L.; de Solla, S.R. Acute and chronic toxicity of neonicotinoid and butenolide insecticides to the freshwater amphipod, Hyalella azteca. Ecotoxicol. Environ. Saf. 2019, 175, 215–223. [Google Scholar] [CrossRef]
- Dong, T.; Zhang, B.; Weng, Q.; Hu, Q. The production relationship of destruxins and blastospores of Metarhizium anisopliae with virulence against Plutella xylostella. J. Integr. Agric. 2016, 15, 1313–1320. [Google Scholar] [CrossRef]
- Ríos-Moreno, A.; Carpio, A.; Garrido-Jurado, I.; Arroyo-Manzanares, N.; Lozano-Tovar, M.D.; Arce, L.; Gámiz-Gracia, L.; García-Campaña, A.M.; Quesada-Moraga, E. Production of destruxins by Metarhizium strains under different stress conditions and their detection by using UHPLC-MS/MS. Biocontrol Sci. Technol. 2016, 26, 1298–1311. [Google Scholar] [CrossRef]
- Ravindran, K.; Akutse, S.; Sivaramakrishnan, S.; Wang, L. Determination and characterization of destruxin production in Metarhizium anisopliae Tk6 and formulations for Aedes aegypti mosquitoes control at the field level. Toxicon 2016. [Google Scholar] [CrossRef] [PubMed]
- Bernat, P.; Nykiel-Szymańska, J.; Stolarek, P.; Słaba, M.; Szewczyk, R.; Ró, S. 2,4-dichlorophenoxyacetic acid-induced oxidative stress: Metabolome and membrane modifications in Umbelopsis isabellina, a herbicide degrader. PLoS ONE 2018, 13, e0199677. [Google Scholar] [CrossRef]
- Różalska, S.; Pawłowska, J.; Wrzosek, M.; Tkaczuk, C.; Długoński, J. Utilization of 4-n-nonylphenol by Metarhizium sp. isolates. Acta Biochim. Pol. 2013, 60, 677–682. [Google Scholar] [CrossRef]
- Rózalska, S.; Glińska, S.; Długoński, J. Metarhizium robertsii morphological flexibility during nonylphenol removal. Int. Biodeterior. Biodegrad. 2014, 95, 285–293. [Google Scholar] [CrossRef]
- Nowak, M.; Soboń, A.; Litwin, A.; Różalska, S. 4-n-nonylphenol degradation by the genus Metarhizium with cytochrome P450 involvement. Chemosphere 2019, 220, 324–334. [Google Scholar] [CrossRef]
- Słaba, M.; Szewczyk, R.; Bernat, P.; Długoński, J. Simultaneous toxic action of zinc and alachlor resulted in enhancement of zinc uptake by the filamentous fungus Paecilomyces marquandii. Sci. Total Environ. 2009, 407, 4127–4133. [Google Scholar] [CrossRef]
- Arroyo-Manzanares, N.; Diana Di Mavungu, J.; Garrido-Jurado, I.; Arce, L.; Vanhaecke, L.; Quesada-Moraga, E.; De Saeger, S. Analytical strategy for determination of known and unknown destruxins using hybrid quadrupole-Orbitrap high-resolution mass spectrometry. Anal. Bioanal. Chem. 2017, 409, 3347–3357. [Google Scholar] [CrossRef]
- Siewiera, P.; Bernat, P.; Rózalska, S.; Długoński, J. Estradiol improves tributyltin degradation by the filamentous fungus Metarhizium robertsii. Int. Biodeterior. Biodegrad. 2015, 104, 258–263. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowak, M.; Bernat, P.; Mrozińska, J.; Różalska, S. Acetamiprid Affects Destruxins Production but Its Accumulation in Metarhizium sp. Spores Increases Infection Ability of Fungi. Toxins 2020, 12, 587. https://doi.org/10.3390/toxins12090587
Nowak M, Bernat P, Mrozińska J, Różalska S. Acetamiprid Affects Destruxins Production but Its Accumulation in Metarhizium sp. Spores Increases Infection Ability of Fungi. Toxins. 2020; 12(9):587. https://doi.org/10.3390/toxins12090587
Chicago/Turabian StyleNowak, Monika, Przemysław Bernat, Julia Mrozińska, and Sylwia Różalska. 2020. "Acetamiprid Affects Destruxins Production but Its Accumulation in Metarhizium sp. Spores Increases Infection Ability of Fungi" Toxins 12, no. 9: 587. https://doi.org/10.3390/toxins12090587
APA StyleNowak, M., Bernat, P., Mrozińska, J., & Różalska, S. (2020). Acetamiprid Affects Destruxins Production but Its Accumulation in Metarhizium sp. Spores Increases Infection Ability of Fungi. Toxins, 12(9), 587. https://doi.org/10.3390/toxins12090587