Mycotoxin Fumonisin B1 Interferes Sphingolipid Metabolisms and Neural Tube Closure during Early Embryogenesis in Brown Tsaiya Ducks

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Developmental Toxicity of FB1 in Brown Tsaiya Duck (BTD) Embryos
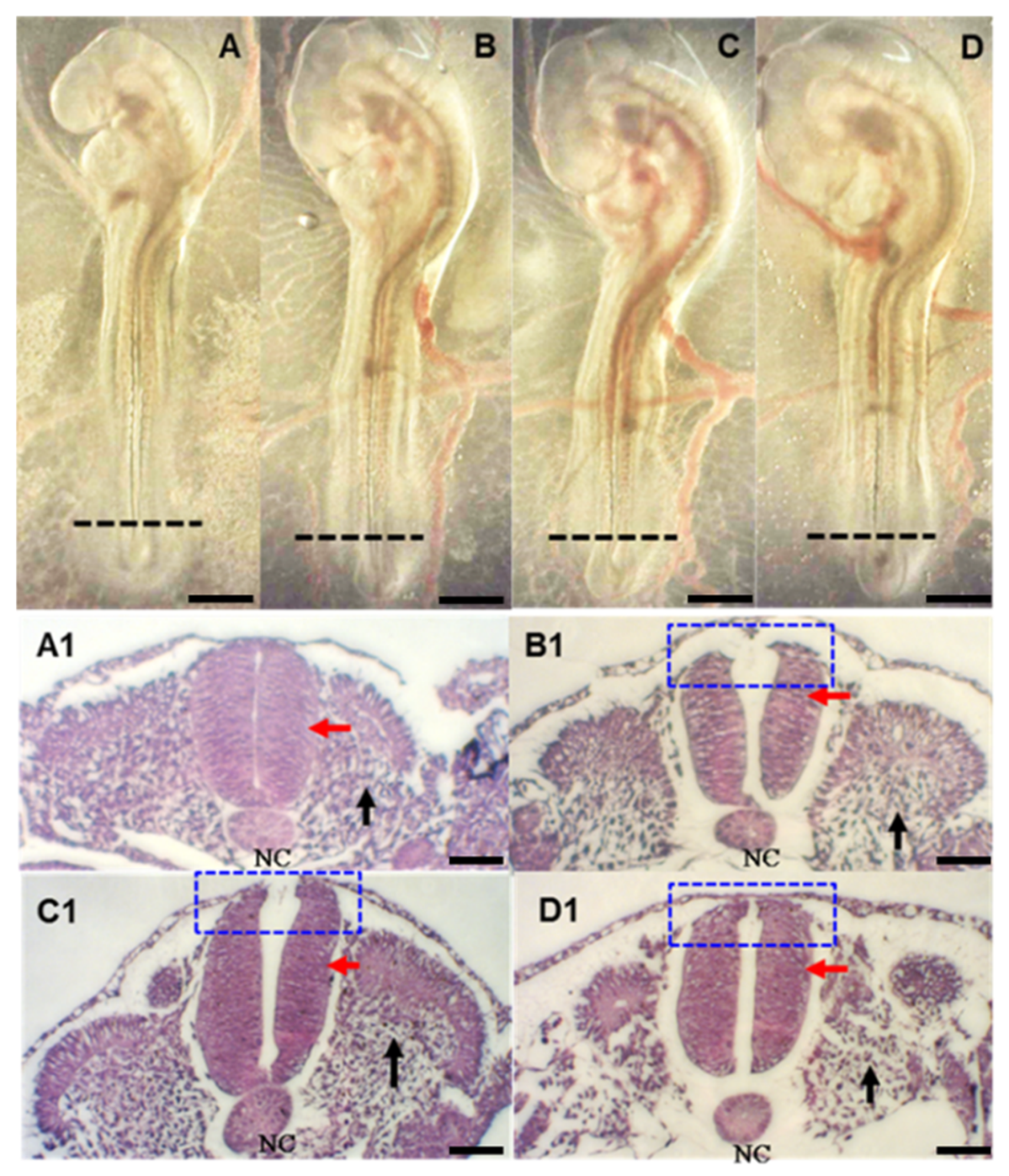
2.2. FB1 Induces Abnormal Neurulation and Somitogenesis during Early Embryogenesis
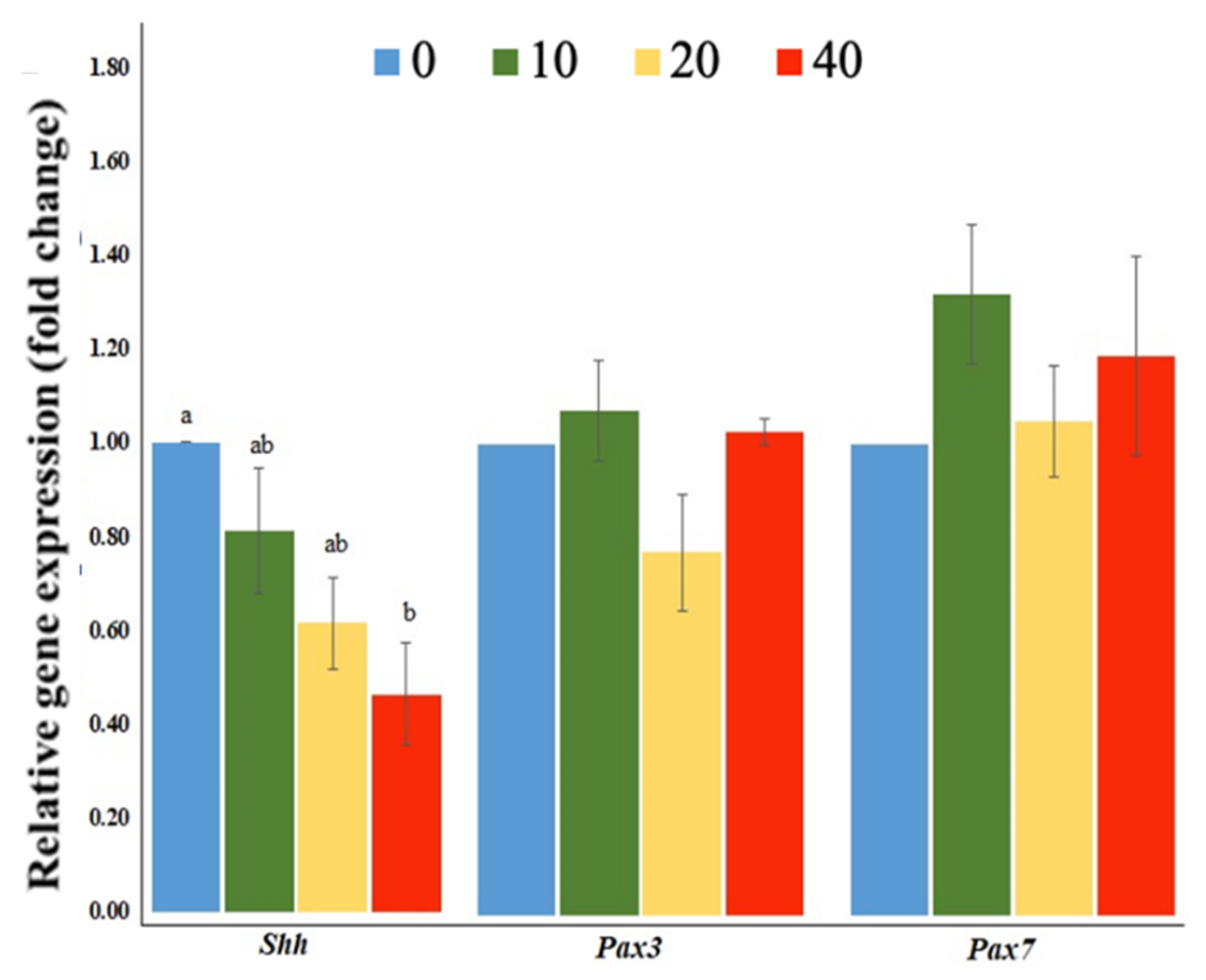
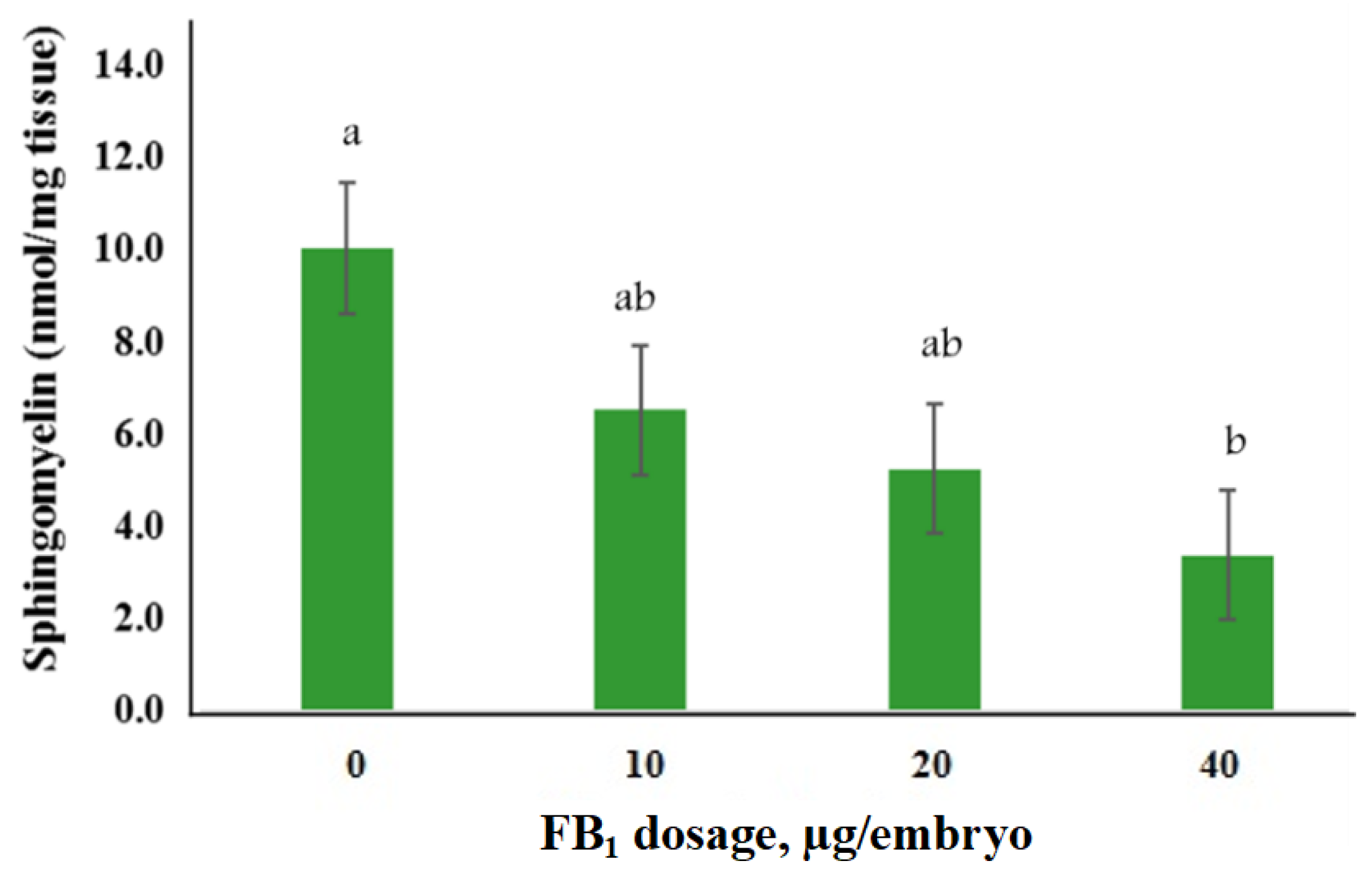
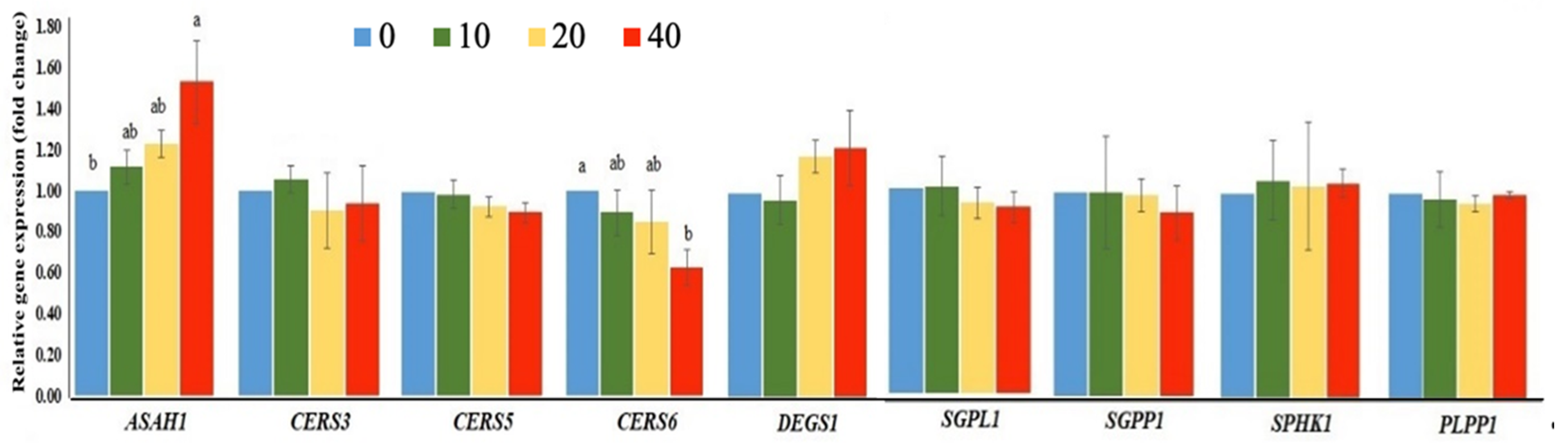
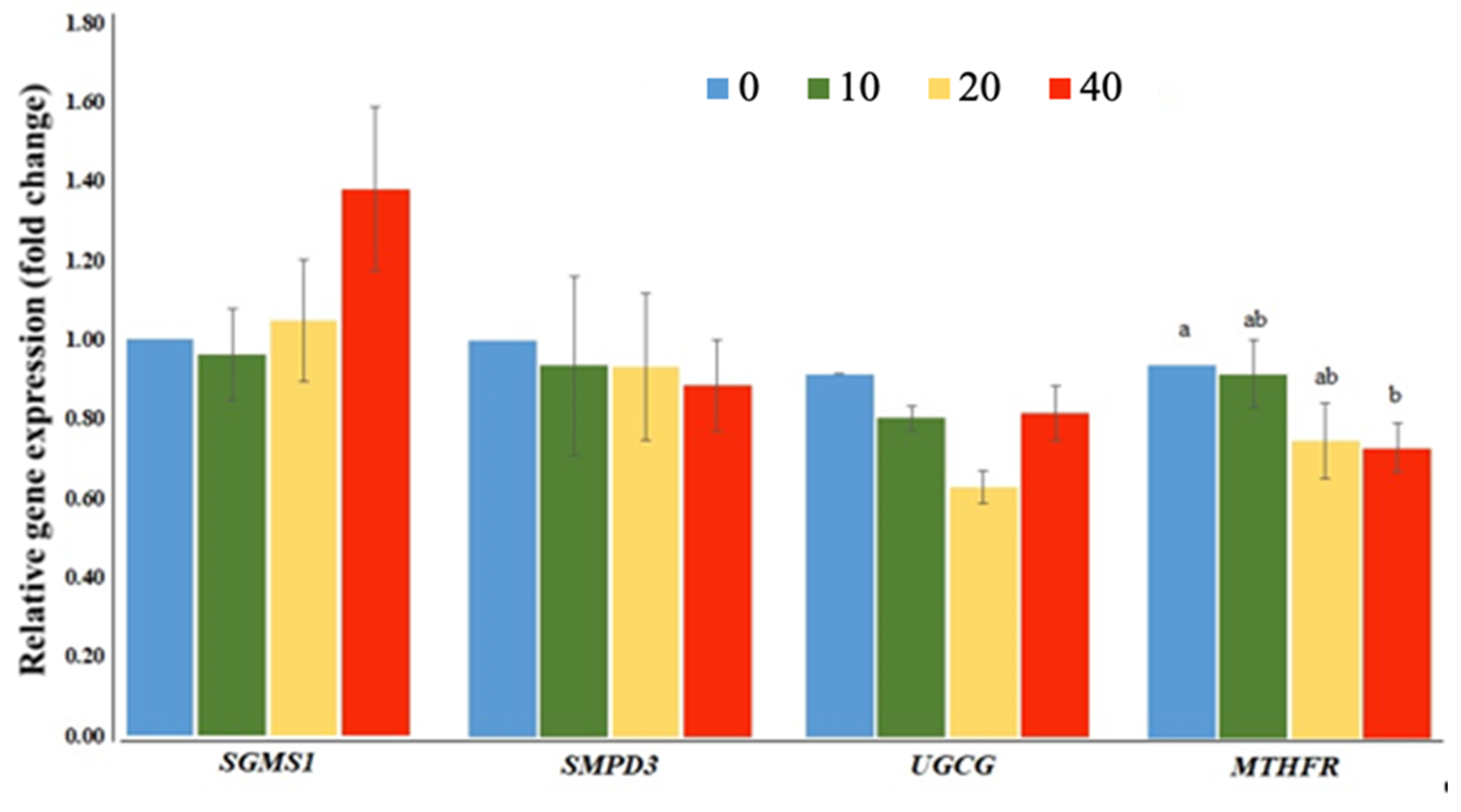
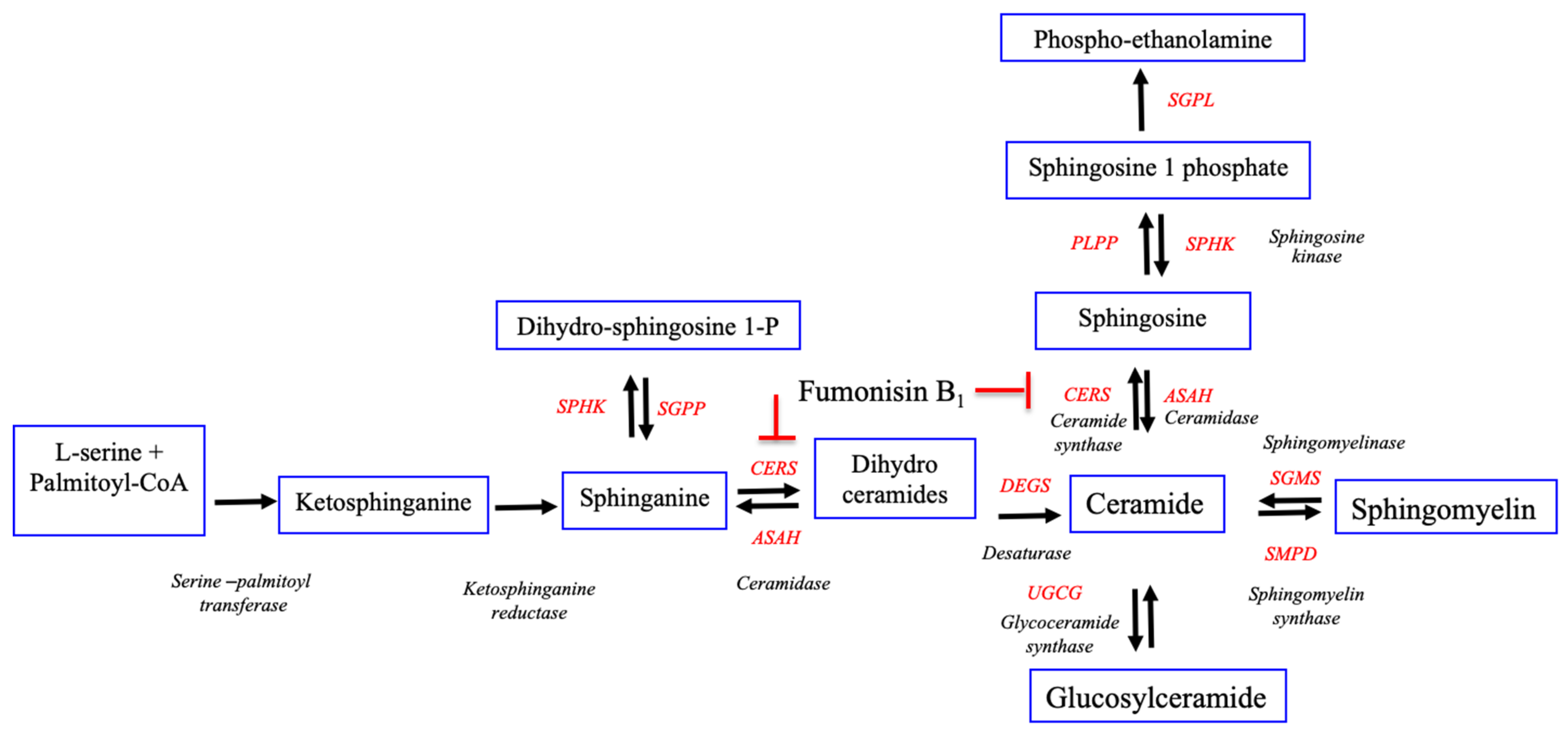
2.3. Exposure to FB1 Alters the Sphingolipid Metabolism and the Related-Gene Expression
3. Discussion
4. Materials and Methods
4.1. Eggs Used and Conditions of Incubation
4.2. Treatment Concentrations of Fumonisin B1
4.3. Embryo Viability and Development
4.4. Histological Assessments
4.5. Sphingomyelin Quantification by Colorimetric Assay
4.6. Revers Transcription and Quantitative Real-Time PCR (qPCR) Analyses
4.7. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bezuidenhout, S.C.; Gelderblom, W.C.; Gorst-Allman, C.P.; Horak, R.M.; Marasas, W.F.; Spiteller, G.; Vleggaar, R. Structure elucidation of the fumonisins, mycotoxins from Fusarium moniliforme. J. Chem. Soc. Chem. Commun. 1988, 11, 743–745. [Google Scholar] [CrossRef]
- Gelderblom, W.; Jaskiewicz, K.; Marasas, W.; Thiel, P.; Horak, R.; Vleggaar, R.; Kriek, N. Fumonisins—Novel mycotoxins with cancer-promoting activity produced by Fusarium moniliforme. Appl. Environ. Microbiol. 1988, 54, 1806–1811. [Google Scholar] [CrossRef] [Green Version]
- Anfossi, L.; Giovannoli, C.; Baggiani, C. Mycotoxin detection. Curr. Opin. Biotechnol. 2016, 37, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Commission Regulation. Commission Regulation (EC) No. 1126/2007 of 28 September 2007 Amending Regulation (EC) No. 1881/2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs as Regards Fusarium Toxins in Maize and Maize Products. Off. J. Eur. Union 2007, 255, 14–17. [Google Scholar]
- El Darra, N.; Gambacorta, L.; Solfrizzo, M. Multimycotoxins occurrence in spices and herbs commercialized in Lebanon. Food Control 2019, 95, 63–70. [Google Scholar] [CrossRef]
- Mendes, G.D.R.L.; Reis, T.A.D.; Corrêa, B.; Badiale-Furlong, E. Mycobiota and occurrence of Fumonisin B1 in wheat harvested in Southern Brazil. Ciência. Rural. 2015, 45, 1050–1057. [Google Scholar] [CrossRef]
- Thiel, P.G.; Shephard, G.S.; Sydenham, E.W.; Marasas, W.F.; Nelson, P.E.; Wilson, T.M. Levels of fumonisins B1 and B2 in feeds associated with confirmed cases of equine leukoencephalomalacia. J. Agric. Food Chem. 1991, 39, 109–111. [Google Scholar] [CrossRef]
- Shephard, G.S.; Thiel, P.G.; Stockenström, S.; Sydenham, E.W. Worldwide survey of fumonisin contamination of corn and corn-based products. J. AOAC Int. 1996, 79, 671–687. [Google Scholar] [CrossRef] [Green Version]
- Howard, P.C.; Couch, L.H.; Patton, R.E.; Eppley, R.M.; Doerge, D.R.; Churchwell, M.I.; Marques, M.M.; Okerberg, C.V. Comparison of the toxicity of several fumonisin derivatives in a 28-day feeding study with female B6C3F1 mice. Toxicol. Appl. Pharmacol. 2002, 185, 153–165. [Google Scholar] [CrossRef]
- Buszewska-Forajta, M. Mycotoxins, invisible danger of feedstuff with toxic effect on animals. Toxicon 2020, 182, 34–53. [Google Scholar] [CrossRef]
- Marasas, W.F. Fumonisins: History, world-wide occurrence and impact. Fumonisins Food 1996, 392, 1–17. [Google Scholar]
- Rheeder, J.P.; Marasas, W.F.; Thiel, P.G.; Sydenham, E.W.; Shephard, G.S.; Van Schalkwyk, D.J. Fusarium moniliforme and fumonisins in corn in relation to human esophageal cancer in Transkei. Phytopathology. Postharvest Pathol. Mycotoxins 1992, 82, 353–357. [Google Scholar]
- Van der Westhuizen, L.; Shephard, G.S.; Rheeder, J.; Burger, H.-M. Individual fumonisin exposure and sphingoid base levels in rural populations consuming maize in South Africa. Food Chem. Toxicol. 2010, 48, 1698–1703. [Google Scholar] [CrossRef]
- Marasas, W.F.O.; Kellerman, T.S.; Gelderblom, W.C.; Thiel, P.; Van der Lugt, J.J.; Coetzer, J.A. Leukoencephalomalacia in a horse induced by fumonisin B1 isolated from Fusarium moniliforme. Onderstepoort J. Vet. Res. 1988, 55, 197–203. [Google Scholar]
- Colvin, B.M.; Harrison, L.R. Fumonisin-induced pulmonary edema and hydrothorax in swine. Mycopathologia 1992, 117, 79–82. [Google Scholar] [CrossRef]
- Voss, K.; Smith, G.; Haschek, W. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Voss, K.A.; Chamberlain, W.J.; Bacon, C.W.; Norred, W.P. A preliminary investigation on renal and hepatic toxicity in rats fed purified fumonisin B1. Nat. Toxins. 1993, 1, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.A.; Riley, R.T.; Norred, W.; Bacon, C.W.; Meredith, F.I.; Howard, P.C.; Plattner, R.D.; Collins, T.; Hansen, D.K.; Porter, J.K. An overview of rodent toxicities: Liver and kidney effects of fumonisins and Fusarium moniliforme. Environ. Health Perspect. 2001, 109, 259–266. [Google Scholar] [PubMed] [Green Version]
- Bouhet, S.; Oswald, I.P. The intestine as a possible target for fumonisin toxicity. Mol. Nutr. Food Res. 2007, 51, 925–931. [Google Scholar] [CrossRef]
- Bouhet, S.; Hourcade, E.; Loiseau, N.; Fikry, A.; Martinez, S.; Roselli, M.; Galtier, P.; Mengheri, E.; Oswald, I.P. The mycotoxin fumonisin B1 alters the proliferation and the barrier function of porcine intestinal epithelial cells. Toxicol. Sci. 2004, 77, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Loiseau, N.; Polizzi, A.; Dupuy, A.; Therville, N.; Rakotonirainy, M.; Loy, J.; Viadere, J.-L.; Cossalter, A.-M.; Bailly, J.-D.; Puel, O. New insights into the organ-specific adverse effects of fumonisin B1: Comparison between lung and liver. Arch. Toxicol. 2015, 89, 1619–1629. [Google Scholar] [CrossRef]
- Smith, G.W.; Constable, P.D.; Eppley, R.M.; Tumbleson, M.E.; Gumprecht, L.A.; Haschek-Hock, W.M. Purified fumonisin B1 decreases cardiovascular function but does not alter pulmonary capillary permeability in swine. Toxicol. Sci. 2000, 56, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.; Norred, W.; Bacon, C.; Riley, R.; Merrill, A.H., Jr. Inhibition of sphingolipid biosynthesis by fumonisins. Implications for diseases associated with Fusarium moniliforme. J. Biol. Chem. 1991, 266, 14486–14490. [Google Scholar] [CrossRef]
- Wang, E.; Ross, P.F.; Wilson, T.M.; Riley, R.T.; Merrill, A.H., Jr. Increases in serum sphingosine and sphinganine and decreases in complex sphingolipids in ponies given feed containing fumonisins, mycotoxins produced by Fusarium moniliforme. J. Nutr. 1992, 122, 1706–1716. [Google Scholar] [CrossRef]
- Merrill Jr, A.H.; Sullards, M.C.; Wang, E.; Voss, K.A.; Riley, R.T. Sphingolipid metabolism: Roles in signal transduction and disruption by fumonisins. Environ. Health Perspect. 2001, 109, 283–289. [Google Scholar]
- Riley, R.T.; An, N.-H.; Showker, J.L.; Yoo, H.-S.; Norred, W.P.; Chamberlain, W.J.; Wang, E.; Merrill, A.H.; Motelin, G.; Beasley, V.R. Alteration of tissue and serum sphinganine to sphingosine ratio: An early biomarker of exposure to fumonisin-containing feeds in pigs. Toxicol. Appl. pharmacol. 1993, 118, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Loiseau, N.; Debrauwer, L.; Sambou, T.; Bouhet, S.; Miller, J.D.; Martin, P.G.; Viadère, J.-L.; Pinton, P.; Puel, O.; Pineau, T. Fumonisin B1 exposure and its selective effect on porcine jejunal segment: Sphingolipids, glycolipids and trans-epithelial passage disturbance. Biochem. Pharmacol. 2007, 74, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.T.; Enongene, E.; Voss, K.A.; Norred, W.P.; Meredith, F.I.; Sharma, R.P.; Spitsbergen, J.; Williams, D.E.; Carlson, D.B.; Merrill, A.H., Jr. Sphingolipid perturbations as mechanisms for fumonisin carcinogenesis. Environ. Health Perspect. 2001, 109, 301–308. [Google Scholar]
- Marasas, W.F.; Riley, R.T.; Hendricks, K.A.; Stevens, V.L.; Sadler, T.W.; Gelineau-van Waes, J.; Missmer, S.A.; Cabrera, J.; Torres, O.; Gelderblom, W.C. Fumonisins disrupt sphingolipid metabolism, folate transport, and neural tube development in embryo culture and in vivo: A potential risk factor for human neural tube defects among populations consuming fumonisin-contaminated maize. J. Nutr. 2004, 134, 711–716. [Google Scholar] [CrossRef]
- Tidhar, R.; Futerman, A.H. The complexity of sphingolipid biosynthesis in the endoplasmic reticulum. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 2511–2518. [Google Scholar] [CrossRef] [Green Version]
- Jan, J.-T.; Chatterjee, S.; Griffin, D.E. Sindbis virus entry into cells triggers apoptosis by activating sphingomyelinase, leading to the release of ceramide. J. Virol. 2000, 74, 6425–6432. [Google Scholar] [CrossRef] [Green Version]
- Lumsangkul, C.; Fan, Y.-K.; Chang, S.-C.; Ju, J.-C.; Chiang, H.-I. Characterizing early embryonic development of Brown Tsaiya Ducks (Anas platyrhynchos) in comparison with Taiwan Country Chicken (Gallus gallus domestics). PLoS ONE 2018, 13, e0196973. [Google Scholar] [CrossRef] [Green Version]
- Lumsangkul, C.; Chiang, H.-I.; Lo, N.-W.; Fan, Y.-K.; Ju, J.-C. Developmental toxicity of mycotoxin Fumonisin B1 in animal embryogenesis: An overview. Toxins 2019, 11, 114. [Google Scholar] [CrossRef] [Green Version]
- Tufan, A.C.; Akdogan, I.; Adiguzel, E. Shell-less culture of the chick embryo as a model system in the study of developmental neurobiology. Neuroanatomy 2004, 3, 8–11. [Google Scholar]
- Song, G.; Cui, Y.; Han, Z.-J.; Xia, H.-F.; Ma, X. Effects of choline on sodium arsenite-induced neural tube defects in chick embryos. Food Chem. Toxicol. 2012, 50, 4364–4374. [Google Scholar] [CrossRef]
- Ma, Z.-L.; Qin, Y.; Wang, G.; Li, X.-D.; He, R.-R.; Chuai, M.; Kurihara, H.; Yang, X. Exploring the caffeine-induced teratogenicity on neurodevelopment using early chick embryo. PLoS ONE 2012, 7, e34278. [Google Scholar]
- Chen, Y.; Fan, J.-X.; Zhang, Z.-L.; Wang, G.; Cheng, X.; Chuai, M.; Lee, K.K.H.; Yang, X. The negative influence of high-glucose ambience on neurogenesis in developing quail embryos. PLoS ONE 2013, 8, e66646. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, G.; Ma, Z.-L.; Li, Y.; Wang, X.-Y.; Cheng, X.; Chuai, M.; Tang, S.-Z.; Lee, K.K.H.; Yang, X. Adverse effects of high glucose levels on somite and limb development in avian embryos. Food Chem. Toxicol. 2014, 71, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wang, G.; Ma, Z.-L.; Chen, Y.-Y.; Fan, J.-J.; Zhang, Z.-L.; Lee, K.K.H.; Luo, H.-M.; Yang, X. Exposure to 2, 5-hexanedione can induce neural malformations in chick embryos. Neurotoxicology 2012, 33, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.-J.; Yang, J.-R.; Chen, S.-E.; Wu, S.-J.; Huang, S.-Y.; Lin, J.-J.; Chen, L.-R.; Tang, P.-C. Inhibition of fumonisin B1 cytotoxicity by nanosilicate platelets during mouse embryo development. PLoS ONE 2014, 9, e112290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolopoulou, E.; Galea, G.L.; Rolo, A.; Greene, N.D.; Copp, A.J. Neural tube closure: Cellular, molecular and biomechanical mechanisms. Development 2017, 144, 552–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, N.D.; Copp, A.J. Neural tube defects. Annu. Rev. Neurosci. 2014, 37, 221–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dequéant, M.-L.; Pourquié, O. Segmental patterning of the vertebrate embryonic axis. Nat. Rev. Genet. 2008, 9, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, Y.; Wang, X.-Y.; Chuai, M.; Yeuk-Hon Chan, J.; Lei, J.; Münsterberg, A.; Lee, K.K.H.; Yang, X. Misexpression of BRE gene in the developing chick neural tube affects neurulation and somitogenesis. Mol. Biol. Cell. 2015, 26, 978–992. [Google Scholar] [CrossRef]
- Sela-Donenfeld, D.; Kalcheim, C. Inhibition of noggin expression in the dorsal neural tube by somitogenesis: A mechanism for coordinating the timing of neural crest emigration. Development 2000, 127, 4845–4854. [Google Scholar] [CrossRef]
- Resende, T.P.; Ferreira, M.; Teillet, M.-A.; Tavares, A.T.; Andrade, R.P.; Palmeirim, I. Sonic hedgehog in temporal control of somite formation. Proc. Nati. Acad. Sci. USA 2010, 107, 12907–12912. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.; Litingtung, Y.; Lee, E.; Young, K.E.; Corden, J.L.; Westphal, H.; Beachy, P.A. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 1996, 383, 407–413. [Google Scholar] [CrossRef]
- Otto, A.; Schmidt, C.; Patel, K. Pax3 and Pax7 expression and regulation in the avian embryo. Anat. Embryol. 2006, 211, 293–310. [Google Scholar] [CrossRef]
- Galli, L.M.; Knight, S.R.; Barnes, T.L.; Doak, A.K.; Kadzik, R.S.; Burrus, L.W. Identification and characterization of subpopulations of Pax3 and Pax7 expressing cells in developing chick somites and limb buds. Dev. Dyn. 2008, 237, 1862–1874. [Google Scholar] [CrossRef] [Green Version]
- Relaix, F.; Rocancourt, D.; Mansouri, A.; Buckingham, M. Divergent functions of murine Pax3 and Pax7 in limb muscle development. Genes Dev. 2004, 18, 1088–1105. [Google Scholar] [CrossRef] [Green Version]
- Eliyahu, E.; Park, J.-H.; Shtraizent, N.; He, X.; Schuchman, E.H. Acid ceramidase is a novel factor required for early embryo survival. FASEB J. 2007, 21, 1403–1409. [Google Scholar] [CrossRef]
- Saad, A.F.; Meacham, W.D.; Bai, A.; Anelli, V.; Anelli, V.; Mahdy, A.E.; Turner, L.S.; Cheng, J.; Bielawska, A.; Bielawski, J. The functional effects of acid ceramidase over-expression in prostate cancer progression and resistance to chemotherapy. Cancer Biol. Ther. 2007, 6, 1451–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, S.T.; Tardieu, D.; Auvergne, A.; Bailly, J.D.; Babilé, R.; Durand, S.; Benard, G.; Guerre, P. Serum sphinganine and the sphinganine to sphingosine ratio as a biomarker of dietary fumonisins during chronic exposure in ducks. Chem. Biol. Interact. 2006, 160, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Bailly, J.-D.; Benlashehr, I.; Auby, A.; Jouglar, J.-Y.; Guerre, P. Tissue persistence of fumonisin B1 in ducks and after exposure to a diet containing the maximum European tolerance for fumonisins in avian feeds. Chem. Biol. Interact. 2009, 182, 239–244. [Google Scholar] [CrossRef]
- Ben-David, O.; Futerman, A.H. The role of the ceramide acyl chain length in neurodegeneration: Involvement of ceramide synthases. Neuromol. Med. 2010, 12, 341–350. [Google Scholar] [CrossRef]
- Riley, R.T.; Merrill, A.H. Ceramide synthase inhibition by fumonisins: A perfect storm of perturbed sphingolipid metabolism, signaling, and disease. J. Lipid Res. 2019, 60, 1183–1189. [Google Scholar] [CrossRef] [Green Version]
- Spassieva, S.D.; Ji, X.; Liu, Y.; Gable, K.; Bielawski, J.; Dunn, T.M.; Bieberich, E.; Zhao, L. Ectopic expression of ceramide synthase 2 in neurons suppresses neurodegeneration induced by ceramide synthase 1 deficiency. Proc. Nat. Acad. Sci. USA 2016, 113, 5928–5933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahfoud, R.; Maresca, M.; Santelli, M.; Pfohl-Leszkowicz, A.; Puigserver, A.; Fantini, J. pH-dependent interaction of fumonisin B1 with cholesterol: Physicochemical and molecular modeling studies at the air− water interface. J. Agric. Food Chem. 2002, 50, 327–331. [Google Scholar] [CrossRef]
- Rog, T.; Vattulainen, I. Cholesterol, sphingolipids, and glycolipids: What do we know about their role in raft-like membranes? Chem. Phys. Lipids. 2014, 184, 82–104. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | FB1 Dosage, µg/Embryo | p Value | |||
|---|---|---|---|---|---|
| 0 1 | 10 | 20 | 40 | ||
| No. of embryo | 18 | 19 | 16 | 18 | |
| Viability, % (n) 2 | 100 (18) | 100 (19) | 100 (16) | 100 (18) | |
| Embryonic stage 3 | 16–17 | 16–17 | 10–17 | 12–17 | |
| ECTL 4, mm | 4.43–7.41 | 5.28–6.54 | 3.02–8.35 | 3.08–6.59 | |
| No. of somite | 30.3 ± 1.70 | 29.9 ± 1.73 | 26.5 ± 6.74 | 27.9 ± 4.14 | |
| Malformations or delay, % (n) 5 | 0 b | 73.7 (14) a | 81.3 (13) a | 88.9 (16) a | <0.0001 |
| Parameter | FB1 Dosage, µg/Embryo | p Value | |||
|---|---|---|---|---|---|
| 0 1 | 10 | 20 | 40 | ||
| No. of Embryos | 10 | 10 | 9 | 8 | |
| Failure of neural tube closure, % (n) | 0 b | 50 (5) a | 55.6(5)a | 37.5 (3) ab | 0.0417 |
| Abnormal neural tube, % (n) | 0 (0) c | 50 (5) b | 100 (9) a | 100 (8) a | <0.0001 |
| Abnormal somites, % (n) | 10 (1) b | 100 (10) a | 100 (9) a | 100 (8) a | <0.0001 |
| Neural tube width, mm | 0.51 ± 0.09 | 0.51 ± 0.13 | 0.59 ± 0.20 | 0.52 ± 0.08 | 0.4510 |
| Target Genes | Primers | Sequences (5′/3′) | Product Size | Accession No. |
|---|---|---|---|---|
| Sphingolipid metabolism pathway | ||||
| CERS3 | Forward | GTGCCACGTTGTATCAACCT | 172 | XM_005011097 |
| Reverse | TCGCTTCGGTCGTCTTTCAA | |||
| CERS6 | Forward | CCTTCTGTTCCTTACGTTTGCC | 153 | XM_038182293 |
| Reverse | TGAAGAACCACAAGCAACACA | |||
| CERS5 | Forward | ATCATTCGCACCGCCTACAA | 202 | XM_021273744 |
| Reverse | ACCCACAGCCTTACTCCTCT | |||
| SPHK1 | Forward | ACTGCACCTTCATCCTCTGC | 234 | XM_027471697 |
| Reverse | AAAACAGGTAGGAGAGGCGG | |||
| DEGS1 | Forward | GCACCACGACTTCCCCAATA | 155 | XM_027453068 |
| Reverse | TCATGCGTGAATATGGGCTGA | |||
| SGPL1 | Forward | TTCCCTTCCACGTTGATGCC | 212 | XM_013107606 |
| Reverse | GGGTGCCACAAAGAACTGGT | |||
| SGPP1 | Forward | TGCTGGTGTTTATTGGTTTGCT | 205 | XM_005022680 |
| Reverse | TGAAAGAGAAGATGCCCAGGG | |||
| ASAH1 | Forward | AGGATGCAAAAGACAAACTGGC | 158 | XM_038178632 |
| Reverse | CACATACCACGTGCCCTTCT | |||
| PLPP1 | Forward | TGTAGTGACGAATCCATCCAGT | 248 | XM_038169951 |
| Reverse | GCATACTTGGCAATGTCCGTC | |||
| Sphingomyelin synthase | ||||
| SMPD3 | Forward | GGTCTACAGTTGCCATGCCT | 182 | XM_021276493 |
| Reverse | GGTCCTGAGGTGTACTTCCC | |||
| SGMS1 | Forward | ATCACTGGCTTTGCTGGACA | 194 | XM_005017312 |
| Reverse | GTACCACCAGACCCTTGCAA | |||
| Glucosylceramide synthase | ||||
| UGCG | Forward | ACAGACAGGGATTTGCTGCT | 152 | XM_038170717 |
| Reverse | CAGTCCTCCAGCTTGATCCA | |||
| Folate metabolism pathway | ||||
| MTHFR | Forward | CCTGGCATCTTCCCCATACA | 226 | XM_013101210 |
| Reverse | TAGCCACTTCCCGATTGAGG | |||
| Development of neural tube and somite | ||||
| Pax3 | Forward | GTCAATCAGCTCGGAGGAGT | 166 | XM_038183383 |
| Reverse | TCTCCTGGTACCTGCAGAGA | |||
| Pax7 | Forward | GAGTTCAGGTGTGGTTCAGCA | 169 | NM_001310395 |
| Reverse | GAAATGGTGGTGGTTGGGTAG | |||
| Shh | Forward | AGGAGTCGCTGCATTACGAG | 250 | XM_038175308 |
| Reverse | CTCAGGTCCTTCACCAGCTT | |||
| Housekeeping gene | ||||
| GAPDH | Forward | CTGGCATTGCACTGAACGAC | 165 | XM_038180584 |
| Reverse | CTCCAACAAAGGGTCCTGCT | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lumsangkul, C.; Tso, K.-H.; Fan, Y.-K.; Chiang, H.-I.; Ju, J.-C. Mycotoxin Fumonisin B1 Interferes Sphingolipid Metabolisms and Neural Tube Closure during Early Embryogenesis in Brown Tsaiya Ducks. Toxins 2021, 13, 743. https://doi.org/10.3390/toxins13110743
Lumsangkul C, Tso K-H, Fan Y-K, Chiang H-I, Ju J-C. Mycotoxin Fumonisin B1 Interferes Sphingolipid Metabolisms and Neural Tube Closure during Early Embryogenesis in Brown Tsaiya Ducks. Toxins. 2021; 13(11):743. https://doi.org/10.3390/toxins13110743
Chicago/Turabian StyleLumsangkul, Chompunut, Ko-Hua Tso, Yang-Kwang Fan, Hsin-I Chiang, and Jyh-Cherng Ju. 2021. "Mycotoxin Fumonisin B1 Interferes Sphingolipid Metabolisms and Neural Tube Closure during Early Embryogenesis in Brown Tsaiya Ducks" Toxins 13, no. 11: 743. https://doi.org/10.3390/toxins13110743
APA StyleLumsangkul, C., Tso, K.-H., Fan, Y.-K., Chiang, H.-I., & Ju, J.-C. (2021). Mycotoxin Fumonisin B1 Interferes Sphingolipid Metabolisms and Neural Tube Closure during Early Embryogenesis in Brown Tsaiya Ducks. Toxins, 13(11), 743. https://doi.org/10.3390/toxins13110743