Updated Review of the Toxicity of Selected Fusarium Toxins and Their Modified Forms

 , , , and
, , , and
Abstract
:1. Introduction
2. Metabolism
2.1. DON
2.2. T-2 and HT-2 Toxins
2.3. ZEN
3. Modified Forms of DON
3.1. In Vitro Cytotoxicity
3.2. Cytotoxicity in In Vivo Systems
3.3. Immunotoxicity
3.4. Interactions
4. Modified Forms of T-2 and HT-2
4.1. Cytotoxicity
4.2. Studies in In Vivo Systems
4.3. Immunotoxicity
4.4. Interactions
4.5. Bioinformatic Evaluation of Toxicity
4.6. Metabolism vs. Toxicity
5. Modified Forms of ZEN
5.1. Cytotoxicity
5.2. Studies in the In Vivo Systems
5.3. Immunotoxicity
5.4. Interactions
5.5. Estrogenic Activity
5.6. Oxidative Activity
5.7. Induction of Epigenetic Alteration and Modulation of Gene Expression
5.8. Bioinformatic Evaluation of Toxicity
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bennett, J.W. Mycotoxins, mycotoxicoses, mycotoxicology and Mycopathologia. Mycopathologia 1987, 100, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Perincherry, L.; Lalak-Kańczugowska, J.; Stępień, Ł. Fusarium-produced mycotoxins in plant-pathogen interactions. Toxins 2019, 11, 664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bönnighausen, J.; Schauer, N.; Schäfer, W.; Bormann, J. Metabolic profiling of wheat rachis node infection by Fusarium graminearum—Decoding deoxynivalenol-dependent susceptibility. New Phytol. 2019, 221, 459–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whetton, R.L.; Hassall, K.L.; Waine, T.W.; Mouazen, A.M. Hyperspectral measurements of yellow rust and fusarium head blight in cereal crops: Part 1: Laboratory study. Biosyst. Eng. 2018, 166, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Oldenburg, E.; Höppner, F.; Ellner, F.; Weinert, J. Fusarium diseases of maize associated with mycotoxin contamination of agricultural products intended to be used for food and feed. Mycotoxin Res. 2017, 33, 167–182. [Google Scholar] [CrossRef]
- Miedaner, T.; Gwiazdowska, D.; Waśkiewicz, A. Editorial: Management of Fusarium species and their mycotoxins in cereal food and feed. Front. Microbiol. 2017, 8, 1543. [Google Scholar] [CrossRef] [Green Version]
- Commission Regulation (EC) No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32006R1881 (accessed on 28 October 2021).
- Commision Regulation (EC) No 1126/2007 of 28 September 2007 Setting Maximum Levels for Certain Contaminants in Foodstuffs as Regards Fusarium Toxins in Maize and Maize Products. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32007R1126 (accessed on 28 October 2021).
- Commission Recommendation No 2013/165/EU of 27 March 2013 on the Presence of T-2 and HT-2 Toxin in Cereals and Cereal Products. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32013H0165 (accessed on 28 October 2021).
- Köpke, U.; Thiel, B.; Elmholt, S. Handbook of Organic Food Safety and Quality; Woodhead Publishing: Sawston, UK, 2007; pp. 353–391. [Google Scholar]
- Schöneberg, T.; Musa, T.; Forrer, H.R.; Mascher, F.; Bucheli, T.B.; Bertossa, M.; Keller, B.; Vogelgsang, S. Infection conditions of Fusarium graminearum in barley are variety specific and different from those in wheat. Eur. J. Plant Pathol. 2018, 151, 975–989. [Google Scholar] [CrossRef] [Green Version]
- Bernhoft, A.; Torp, M.; Clasen, P.-E.; Løes, A.-K.; Kristoffersen, A. Influence of agronomic and climatic factors on Fusarium infestation and mycotoxin contamination of cereals in Norway. Food Addit. Contam. 2012, 29, 1129–1140. [Google Scholar] [CrossRef] [Green Version]
- Kochiieru, Y.; Mankevičiene, A.; Cesavičiene, J.; Semaškiene, R.; Dabkevičius, Z.; Janavičiene, S. The influence of harvesting time and meterological conditions on the occurrence of Fusarium species and mycotoxin contamination of spring cereals. J. Sci. Food Agric. 2020, 100, 2999–3006. [Google Scholar] [CrossRef]
- Medina, A.; Magan, N. Comparisons of water activity and temperature impacts on growth of Fusarium langsethiae strains from northern Europe on oat based media. Int. J. Food Microbiol. 2010, 142, 365–369. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.M.; Parry, D.W.; Nicholson, P.; Thomsett, M.A.; Simpson, D.; Edwards, S.G.; Cooke, B.M.; Doohan, F.M.; Brennan, J.M.; Moretti, A.; et al. Predominance and association of pathogenic fungi causing Fusarium ear blightin wheat in four European countries. Eur. J. Plant Pathol. 2005, 112, 143–154. [Google Scholar] [CrossRef]
- Schaarschmidt, S.; Fauhl-Hassek, C. The fate of mycotoxins during the primary food processing of maize. Food Control 2021, 121, 107651. [Google Scholar] [CrossRef]
- Kuchenbuch, H.S.; Becker, S.; Schulz, M.; Cramer, B.; Humpf, H.U. Thermal stability of T-2 and HT-2 toxins during biscuit- and crunchy muesli-making and roasting. Food Addit. Contam. Part A 2018, 35, 2158–2167. [Google Scholar] [CrossRef]
- Aitken, A.; Miller, J.D.; McMullin, D.R. Isolation, chemical characterization and hydrolysis of the trichothecene 7α-hydroxy, 15-deacetylcalonectrin (3ANX) from Fusarium graminearum DAOMC 242077. Tetrahedron Lett. 2019, 60, 852–856. [Google Scholar] [CrossRef]
- Kuchenbuch, H.S.; Cramer, B.; Humpf, H.U. Matrix binding of T-2 toxin: Structure elucidation of reaction products and indications on the fate of a relevant food-borne toxin during heating. Mycotoxin Res. 2019, 35, 261–270. [Google Scholar] [CrossRef]
- Metzler, M.; Pfeiffer, E.; Hildebrandm, A. Zearalenone and its metabolites as endocrine disrupting chemicals. World Mycotoxin J. 2010, 3, 385–401. [Google Scholar] [CrossRef]
- Rychlik, M.; Humpf, H.U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Khaneghah, A.M.; Martins, L.M.; von Hertwig, A.M.; Bertoldo, R.; Sant’Ana, A.S. Deoxynivalenol and its masked forms: Characteristics, incidence, control and fate during wheat and wheat based products processing—A review. Trends Food Sci. Technol. 2018, 71, 13–24. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, 4718. [Google Scholar]
- Freire, L.; Sant’Ana, A.S. Modified mycotoxins: An updated review on their formation, detection, occurrence, and toxic effects. Food Chem. Toxicol. 2018, 111, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, N.; Devreese, M.; Demeyere, K.; Berthiller, F.; Michlmayr, H.; Varga, E.; Adam, G.; Meyer, E.; Croubels, S. Comparative in vitro cytotoxicity of modified deoxynivalenol on porcine intestinal epithelial cells. Food Chem. Toxicol. 2016, 95, 103–109. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Puel, O.; Oswald, I.P. Toxicological interactions between the mycotoxins deoxynivalenol, nivalenol and their acetylated derivatives in intestinal epithelial cells. Arch. Toxicol. 2015, 89, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef]
- Knutsen, H.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; DiNovi, M.; Edler, L.; EFSA Panel on Contaminants in the Food Chain (CONTAM); et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, e04851. [Google Scholar]
- Knutsen, H.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Appropriateness to set a group health based guidance value for T2 and HT2 toxin and its modified forms. EFSA J. 2017, 15, e04655. [Google Scholar]
- Juan-García, A.; Juan, C.; Tolosa, J.; Ruiz, M.J. Effects of deoxynivalenol, 3-acetyl-deoxynivalenol and 15-acetyl-deoxynivalenol on parameters associated with oxidative stress in HepG2 cells. Mycotoxin Res. 2019, 35, 197–205. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, S.; Tan, Y.; Liu, N.; Wu, A. Individual and combined cytotoxic effects of co-occurring deoxynivalenol family mycotoxins on human gastric epithelial cells. Toxins 2017, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Taroncher, M.; Rodríguez-Carrasco, Y.; Ruiz, M.J. Interactions between T-2 toxin and its metabolites in HepG2 cells and in silico approach. Food Chem. Toxicol. 2021, 148, 111942. [Google Scholar] [CrossRef]
- Ling, A.; Sun, L.W.; Guo, W.B.; Sun, S.Y.; Yang, J.H.; Zhao, Z.H. Individual and combined cytotoxic effects of T-2 toxin and its four metabolites on porcine Leydig cells. Food Chem. Toxicol. 2020, 139, 111277. [Google Scholar] [CrossRef] [PubMed]
- Tran, V.N.; Viktorova, J.; Augustynkova, K.; Jelenova, N.; Dobiasova, S.; Rehorova, K.; Fenclova, M.; Stranska-Zachariasova, M.; Vitek, L.; Hajslova, J.; et al. In silico and in vitro studies of mycotoxins and their cocktails; Their toxicity and its mitigation by silibinin pre-treatment. Toxins 2020, 12, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, D.E.; Pistol, G.C.; Bulgaru, C.V.; Taranu, I. Cytotoxic and inflammatory effects of individual and combined exposure of HepG2 cells to zearalenone and its metabolites. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Karaman, E.F.; Zeybel, M.; Ozden, S. Evaluation of the epigenetic alterations and gene expression levels of HepG2 cells exposed to zearalenone and α-zearalenol. Toxicol. Lett. 2020, 326, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Bracarense, A.P.F.L.; Cossalter, A.M.; Laffitte, J.; Schwartz-Zimmermann, H.E.; Schatzmayr, G.; Pinton, P.; Moll, W.D.; Oswald, I.P. Deepoxy-deoxynivalenol retains some immune-modulatory properties of the parent molecule deoxynivalenol in piglets. Arch. Toxicol. 2018, 92, 3381–3389. [Google Scholar] [CrossRef]
- Bracarense, A.P.F.L.; Pierron, A.; Pinton, P.; Gerez, J.R.; Schatzmayr, G.; Moll, W.D.; Zhou, T.; Oswald, I.P. Reduced toxicity of 3-epi-deoxynivalenol and de-epoxy-deoxynivalenol through deoxynivalenol bacterial biotransformation: In vivo analysis in piglets. Food Chem. Toxicol. 2020, 140, 111241. [Google Scholar] [CrossRef]
- Juan-García, A.; Taroncher, M.; Font, G.; Ruiz, M.J. Micronucleus induction and cell cycle alterations produced by deoxynivalenol and its acetylated derivatives in individual and combined exposure on HepG2 cells. Food Chem. Toxicol. 2018, 118, 719–725. [Google Scholar] [CrossRef]
- Reinhardt, H.C.; Yaffe, M.B. Kinases that control the cell cycle in response to DNA damage: Chk1, Chk2, and MK2. Curr. Opin. Cell Biol. 2009, 21, 245–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Yu, S.; Jia, B.X.; Liu, N.; Wu, A. Metabolomic profiling reveals similar cytotoxic effects and protective functions of quercetin during deoxynivalenol- and 15-acetyl deoxynivalenol-induced cell apoptosis. Toxicol. Vitro 2020, 66, 104838. [Google Scholar] [CrossRef]
- Wang, X.; Fan, M.; Chu, X.; Zhang, Y.; Rahman, S.U.; Jiang, Y.; Chen, X.; Zhu, D.; Feng, S.; Li, Y. Deoxynivalenol induces toxicity and apoptosis in piglet hippocampal nerve cells via the MAPK signaling pathway. Toxicon 2018, 155, 1–8. [Google Scholar] [CrossRef]
- He, Y.; Yin, X.; Dong, J.; Yang, Q.; Wu, Y.; Gong, Z. Transcriptome analysis of Caco-2 cells upon the exposure of mycotoxin deoxynivalenol and its acetylated derivatives. Toxins 2021, 13, 167. [Google Scholar] [CrossRef] [PubMed]
- Agahi, F.; Juan, C.; Font, G.; Juan-García, A. In silico methods for metabolomic and toxicity prediction of zearalenone, α-zearalenone and β-zearalenone. Food Chem. Toxicol. 2020, 146, 111818. [Google Scholar] [CrossRef]
- Hasan, N.; Bhuiya, N.M.M.A.; Hossain, M.K. In silico molecular docking, PASS prediction, and ADME/T analysis for finding novel COX-2 inhibitor from Heliotropium indicum. J. Comp. Med. Res. 2019, 10, 142–154. [Google Scholar] [CrossRef]
- Bai, Y.; Zheng, J.; Yuan, X.; Jiao, S.; Feng, C.; Du, Y.; Liu, H.; Zheng, L. Chitosan oligosaccharides improve glucolipid metabolism disorder in liver by suppression of obesity-related inflammation and restoration of peroxisome proliferator-activated receptor gamma (PPARγ). Mar. Drugs 2018, 16, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [Green Version]
- Agahi, F.; Font, G.; Juan, C.; Juan-García, A. Individual and combined effect of zearalenone derivates and beauvericin mycotoxins on SH-SY5Y cells. Toxins 2020, 12, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riahi, I.; Ramos, A.J.; Pérez-Vendrell, A.M.; Marquis, V. A toxicokinetic study reflecting the absorption, distribution, metabolism and excretion of deoxynivalenol in broiler chickens. J. Appl. Anim. Res. 2021, 49, 284–288. [Google Scholar] [CrossRef]
- Jurisic, N.; Schwartz-Zimmermann, H.E.; Kunz-Vekiru, E.; Reisinger, N.; Klein, S.; Caldwell, D.; Fruhmann, P.; Schatzmayr, D.; Berthiller, F. Deoxynivalenol-3-sulphate is the major metabolite of dietary deoxynivalenol in eggs of laying hens. World Mycotoxin J. 2019, 12, 245–255. [Google Scholar] [CrossRef]
- De Zutter, N.; Audenaert, K.; Arroyo-Manzanares, N.; De Boevre, M.; Van Poucke, C.; De Saeger, S.; Haesaert, G.; Smagghe, G. Aphids transform and detoxify the mycotoxin deoxynivalenol via a type II biotransformation mechanism yet unknown in animals. Sci. Rep. 2016, 6, 38640. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.; Clear, R.M.; Ward, T.J.; Gaba, D.; Tekauz, A.; Turkington, T.K.; Woods, S.M.; Nowicki, T.; O’Donnell, K. Relative aggressiveness and production of 3- or 15-acetyl deoxynivalenol and deoxynivalenol by Fusarium graminearum in spring wheat. Can. J. Plant Pathol. 2010, 32, 146–152. [Google Scholar] [CrossRef]
- Albuquerque, D.R.; Patriarca, A.; Pinto, V.F. Can discrepancies between Fusarium graminearum trichothecene genotype and chemotype be explained by the influence of temperature in the relative production of 3-ADON and 15-ADON? Fungal Biol. 2021, 125, 153–159. [Google Scholar] [CrossRef]
- Spaggiari, M.; Righetti, L.; Galaverna, G.; Giordano, D.; Scarpino, V.; Blandino, M.; Dall’Asta, C. HR-MS profiling and distribution of native and modified Fusarium mycotoxins in tritordeum, wheat and barley whole grains and corresponding pearled fractions. J. Cereal Sci. 2019, 87, 178–184. [Google Scholar] [CrossRef]
- Kluger, B.; Bueschl, C.; Lemmens, M.; Michlmayr, H.; Malachova, A.; Koutnik, A.; Maloku, I.; Berthiller, F.; Adam, G.; Krska, R.; et al. Biotransformation of the mycotoxin deoxynivalenol in Fusarium resistant and susceptible near isogenic wheat lines. PLoS ONE 2015, 10, e0119656. [Google Scholar] [CrossRef] [PubMed]
- Warth, B.; Fruhmann, P.; Wiesenberger, G.; Kluger, B.; Sarkanj, B.; Lemmens, M.; Hametner, C.; Fröhlich, J.; Adam, G.; Krska, R.; et al. Deoxynivalenol-sulfates: Identification and quantification of novel conjugated (masked) mycotoxins in wheat. Anal. Bioanal. Chem. 2015, 407, 1033–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlovsky, P. Biological detoxification of the mycotoxin deoxynivalenol and its use in genetically engineered crops and feed additives. Appl. Microbiol. Biotechnol. 2011, 91, 491–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maul, R.; Warth, B.; Schebb, N.H.; Krska, R.; Koch, M.; Sulyok, M. In vitro glucuronidation kinetics of deoxynivalenol by human and animal microsomes and recombinant human UGT enzymes. Arch. Toxicol. 2015, 89, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Claeys, L.; Mengelers, M.; Vanhoorne, V.; Vervaet, C.; Huybrechts, B.; De Saeger, S.; De Boevre, M. Humans significantly metabolize and excrete the mycotoxin deoxynivalenol and its modified form deoxynivalenol-3-glucoside within 24 hours. Sci. Rep. 2018, 8, 5255. [Google Scholar] [CrossRef] [Green Version]
- Vidal, A.; Mengelers, M.; Yang, S.; De Saeger, S.; De Boevre, M. Mycotoxin biomarkers of exposure: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1127–1155. [Google Scholar] [CrossRef] [Green Version]
- Gratz, S.W.; Duncan, G.; Richardson, A.J. Microbial metabolism and intestinal absorption of the mycotoxin deoxynivalenol and its metabolites in humans. Ann. Nutr. Metab. 2013, 63, 1806. [Google Scholar]
- Pestka, J.J.; Clark, E.S.; Schwartz-Zimmermann, H.E.; Berthiller, F. Sex is a determinant for deoxynivalenol metabolism and elimination in the mouse. Toxins 2017, 9, 240. [Google Scholar] [CrossRef] [Green Version]
- Busman, M.; Poling, S.M.; Maragos, C.M. Observation of T-2 toxin and HT-2 toxin glucosides from Fusarium sporotrichioides by liquid chromatography coupled to tandem mass spectrometry (LC-MS/MS). Toxins 2011, 3, 1554–1568. [Google Scholar] [CrossRef] [Green Version]
- McCormick, S.P.; Price, N.P.J.; Kurtzman, C.P. Glucosylation and other biotransformations of T-2 toxin by yeasts of the trichomonascus clade. Appl. Environ. Microbiol. 2012, 78, 8694–8702. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Nie, D.; Fan, K.; Yang, J.; Guo, W.; Meng, J.; Zhao, Z.; Han, Z. A systematic review of plant-conjugated masked mycotoxins: Occurrence, toxicology, and metabolism. Crit. Rev. Food Sci. Nutr. 2020, 60, 1523–1537. [Google Scholar] [CrossRef] [PubMed]
- Kuca, K.; Dohnal, V.; Jezkova, A.; Jun, D. Metabolic pathways of T-2 toxin. Curr. Drug Metabol. 2008, 9, 77–82. [Google Scholar] [CrossRef]
- Righetti, L.; Körber, T.; Rolli, E.; Galaverna, G.; Suman, M.; Bruni, R.; Dall’Asta, C. Plant biotransformation of T2 and HT2 toxin in cultured organs of Triticum durum Desf. Sci. Rep. 2019, 9, 14320. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Beier, R.C.; Shen, J.; Smet, D.D.; De Saeger, S.; Zhang, S. T-2 toxin, a trichothecene mycotoxin: Review of toxicity, metabolism, and analytical methods. J. Agric. Food Chem. 2011, 59, 3441–3453. [Google Scholar] [CrossRef]
- Wu, Q.; Dohnal, V.; Huang, L.; Kuča, K.; Yuan, Z. Metabolic pathways of trichothecenes. Drug Metab. Rev. 2010, 42, 250–267. [Google Scholar] [CrossRef]
- Wu, Q.; Engemann, A.; Cramer, B.; Welsch, T.; Yuan, Z.; Humpf, H.U. Intestinal metabolism of T-2 toxin in the pig cecum model. Mycotoxin Res. 2012, 28, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Qin, Z.; Kuca, K.; You, L.; Zhao, Y.; Liu, A.; Musilek, K.; Chrienova, Z.; Nepovimova, E.; Oleksak, P.; et al. An update on T-2 toxin and its modified forms: Metabolism, immunotoxicity mechanism, and human exposure assessment. Arch. Toxicol. 2020, 94, 3645–3669. [Google Scholar] [CrossRef]
- Borzekowski, A.; Drewitz, T.; Keller, J.; Pfeifer, D.; Kunte, H.-J.; Koch, M.; Rohn, S.; Maul, R. Biosynthesis and characterization of zearalenone-14-sulfate, zearalenone-14-glucoside and zearalenone-16-glucoside using common fungal strains. Toxins 2018, 10, 104. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Tan, Y.; Yan, Z.; Liao, Y.; Chen, J.; De Boevre, M.; De Saeger, S.; Wu, A. Antagonistic and detoxification potentials of trichoderma isolates for control of zearalenone (ZEN) producing Fusarium graminearum. Front. Microbiol. 2018, 8, 2710. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Molto, J.C.; Manes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Bryła, M.; Waśkiewicz, A.; Ksieniewicz-Woźniak, E.; Szymczyk, K.; Jędrzejczak, R. Modified Fusarium mycotoxins in cereals and their products—Metabolism, occurrence, and toxicity: An updated review. Molecules 2018, 23, 963. [Google Scholar] [CrossRef] [Green Version]
- Warth, B.; Sulyok, M.; Berthiller, F.; Schuhmacher, R.; Krska, R. New insights into the human metabolism of the Fusarium mycotoxins deoxynivalenol and zearalenone. Toxicol. Lett. 2013, 220, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Belhassen, H.; Jiménez-Díaz, I.; Ghali, R.; Ghorbel, H.; Molina-Molina, J.M.; Olea, N.; Hedili, A. Validation of a UHPLC–MS/MS method for quantification of zearalenone, α-zearalenol, β-zearalenol, α-zearalanol, β-zearalanol and zearalanone in human urine. J. Chromatogr. B 2014, 962, 68–74. [Google Scholar] [CrossRef]
- Richardson, K.E.; Hagler, W.M.; Mirocha, C.J. Production of zearalenone. alpha.-and. beta.-zearalenol, and. alpha.-and. beta.-zearalanol by Fusarium spp. in rice culture. J. Agric. Food Chem. 1985, 33, 862–866. [Google Scholar] [CrossRef]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef]
- Berthiller, F.; Lemmens, M.; Werner, U.; Krska, R.; Hauser, M.T.; Adam, G.; Schuhmacher, R. Short review: Metabolism of the Fusarium mycotoxins deoxynivalenol and zearalenone in plants. Mycotox Res. 2007, 23, 68–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohn, B.N.; Bunzel, M. Zearalenone-malonyl-glucosides as phase II metabolites in plant cell suspension cultures. Cereal Chem. 2021, 98, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, A.; Braber, S.; Akbari, P.; Kraneveld, A.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol and its modified forms: Are there major differences? Toxins 2016, 8, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, A.E.; McCormick, S.P.; Appell, M. Structure−activity relationships of trichothecene toxins in an Arabidopsis thaliana leaf assay. J. Agric. Food Chem. 2007, 55, 6487–6492. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.L.; Wannemacher, R.W., Jr. Structure-function relationships of 12,13-epoxytrichothecene mycotoxins in cell culture: Comparison to whole animal lethality. Toxicon 1986, 24, 985–994. [Google Scholar] [CrossRef]
- Pinton, P.; Tsybulskyy, D.; Lucioli, J.; Laffitte, J.; Callu, P.; Lyazhri, F.; Grosjean, F.; Bracarense, A.P.F.L.; Kolf-Clauw, M.; Oswald, I.P. Toxicity of deoxynivalenol and its acetylated derivatives on the intestine: Differential effects on morphology, barrier function, tight junction proteins, and mitogen-activated protein kinases. Toxicol. Sci. 2012, 130, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, N.; Devreese, M.; van Bergen, T.; Schauvliege, S.; De Boevre, M.; De Saeger, S.; Vanhaecke, L.; Berthiller, F.; Michlmayr, H.; Malachova, A. Deoxynivalenol-3-β-D-glucoside: In vitro cytotoxicity and in vivo oral bioavailability and hydrolysis in broiler chicken and pig. In Proceedings of the 38th Mycotoxin Workshop, Berlin, Germany, 2–4 May 2016. [Google Scholar]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.P.F.L.; Liaubet, L.; Schatzmayr, G.; Berthiller, F.; Moll, W.M.; et al. Intestinal toxicity of the masked mycotoxin deoxynivalenol-3-β-D-glucoside. Arch. Toxicol. 2016, 90, 2037–2046. [Google Scholar] [CrossRef]
- Springler, A.; Hessenberger, S.; Reisinger, N.; Kern, C.; Nagl, V.; Schatzmayr, G.; Mayer, E. Deoxynivalenol and its metabolite deepoxy-deoxynivalenol: Multi-parameter analysis for the evaluation of cytotoxicity and cellular effects. Mycotoxin Res. 2017, 33, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catteuw, A.; Devreese, M.; De Baere, S.; Antonissen, G.; Ivanova, L.; Uhlig, S.; Martens, A.; De Saeger, S.; De Boevre, M.; Croubels, S. Investigation of age-related differences in toxicokinetic processes of deoxynivalenol and deoxynivalenol-3-glucoside in weaned piglets. Arch. Toxicol. 2020, 94, 417–425. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; van Bergen, T.; Schauvliege, S.; De Boevre, M.; De Saeger, S.; Vanhaecke, L.; Berthiller, F.; Michlmayr, H.; Malachová, A.; et al. In vivo contribution of deoxynivalenol-3-β-d-glucoside to deoxynivalenol exposure in broiler chickens and pigs: Oral bioavailability, hydrolysis and toxicokinetics. Arch. Toxicol. 2017, 91, 699–712. [Google Scholar] [CrossRef] [Green Version]
- Gerez, J.R.; Desto, S.S.; Bracarense, A.P.F.R.L. Deoxynivalenol induces toxic effects in the ovaries of pigs: An ex vivo approach. Theriogenology 2017, 90, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Peng, Z.; Chen, L.; Nüssler, A.K.; Liu, L.; Yang, W. Deoxynivalenol, gut microbiota and immunotoxicity: A potential approach? Food Chem. Toxicol. 2018, 112, 342–354. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, R.; Zhu, M.X.; Zhang, N.Y.; Liu, X.L.; Wang, Y.W.; Qin, T.; Zheng, L.Y.; Liu, Q.; Zhang, W.P.; et al. Effect of deoxynivalenol on the porcine acquired immune response and potential remediation by a novel modified HSCAS adsorbent. Food Chem. Toxicol. 2020, 138, 111187. [Google Scholar] [CrossRef]
- Estienne, A.; Chorfi, Y.; Guerrero-Netro, H.M.; Price, C. The mycotoxin metabolite deepoxy-deoxynivalenol increases apoptosis and decreases steroidogenesis in bovine ovarian theca cells. Biol. Reprod. 2017, 97, 746–757. [Google Scholar]
- Alassane-Kpembi, I.; Kolf-Clauw, M.; Gauthier, T.; Abrami, R.; Abiola, F.A.; Oswald, I.P.; Puel, O. New insights into mycotoxin mixtures: The toxicity of low doses of Type B trichothecenes on intestinal epithelial cells is synergistic. Tetrahedron Lett. 2013, 272, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Juan-García, A.; Juan, C.; Manyes, L.; Ruiz, M.J. Binary and tertiary combination of alternariol, 3-acetyl-deoxynivalenol and 15-acetyl-deoxynivalenol on HepG2 cells: Toxic effects and evaluation of degradation products. Toxicol. In Vitro 2016, 34, 264–273. [Google Scholar] [CrossRef]
- Novak, B.; Vatzia, E.; Springler, A.; Pierron, A.; Gerner, W.; Reisinger, N.; Hessenberger, S.; Schatzmayr, G.; Mayer, E. Bovine peripheral blood mononuclear cells are more sensitive to deoxynivalenol than those derived from poultry and swine. Toxins 2018, 10, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taroncher, M.; Rodríguez-Carrasco, Y.; Ruiz, M.-J. T-2 toxin and its metabolites: Characterization, cytotoxic mechanisms and adaptive cellular response in human hepatocarcinoma (HepG2) cells. Food Chem. Toxicol. 2020, 145, 111654. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, N.; Devreese, M.; De Boevre, M.; De Saeger, S.; Croubels, S. T-2 toxin-3α-glucoside in broiler chickens: Toxicokinetics, absolute oral bioavailability, and in vivo hydrolysis. J. Agric. Food Chem. 2017, 65, 4797–4803. [Google Scholar] [CrossRef] [Green Version]
- Zhanrui, H.; Yaling, W.; Xiaobo, W.; Bing, M.; Mei, Q.; Lijun, S.; Kangjian, C.; Jian, L. Effect of masked T-2 toxin residues in Litopenaeus vannamei on routine blood and serum biochemical indexes of mice. Acta Agric. Zhejiangensis 2017, 29, 1110–1118. [Google Scholar]
- Zhang, J.; Liu, S.; Zhang, H.; Li, Y.; Wu, W.; Zhang, H. Gut satiety hormones cholecystokinin and glucagon-like Peptide-1(7-36) amide mediate anorexia induction by trichothecenes T-2 toxin, HT-2 toxin, diacetoxyscirpenol and neosolaniol. Toxicol. Appl. Pharmacol. 2017, 335, 49–55. [Google Scholar] [CrossRef]
- Zhang, J.; Sheng, K.; Wu, W.; Zhang, H. Anorectic responses to T-2 toxin, HT-2 toxin, diacetoxyscirpenol and neosolaniol correspond to plasma elevations of neurotransmitters 5-hydroxytryptamine and substance P. Ecotoxicol. Environ. Saf. 2018, 161, 451–458. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Liu, S.; Wu, W.; Zhang, H. Comparison of anorectic potencies of type A trichothecenes T-2 toxin, HT-2 toxin, diacetoxyscirpenol, and neosolaniol. Toxins 2018, 10, 179. [Google Scholar] [CrossRef] [Green Version]
- Grundy, D. 5-HT system in the gut: Roles in the regulation of visceral sensitivity and motor functions. Eur. Rev. Med. Pharmacol. Sci. 2008, 12, 63–67. [Google Scholar]
- Jean, A.; Conductier, G.; Manrique, C.; Bouras, C.; Berta, P.; Hen, R.; Charnay, Y.; Bockaert, J.; Compan, V. Anorexia induced by activation of serotonin 5-ht4 receptors is mediated by increases in cart in the nucleus accumbens. Proc. Natl. Acad. Sci. USA 2007, 104, 16335–16340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisch, P.; Bilkei-Gorzó, A.; Rácz, I.; Zimmer, A. Modulation of the CRH system by substance P/NKA in an animal model of depression. Behav. Brain Res. 2010, 213, 103–108. [Google Scholar] [CrossRef]
- Wu, Q.; Kuca, K.; Nepovimova, E.; Wu, W. Type A trichothecene diacetoxyscirpenol-induced emesis corresponds to secretion of peptide YY and serotonin in mink. Toxins 2020, 12, 419. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, X.; Wan, D.; Li, J.; Yuan, Z.H. Crosstalk of JNK1-STAT3 is critical for RAW264.7 cell survival. Cell. Signal. 2014, 26, 2951–2960. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Qiu, M.; Sun, L.; Wang, X.; Li, C.; Xu, D.; Gooneratne, R. Cytotoxicity of T-2 and modified T-2 toxins: Induction of JAK/STAT pathway in RAW264.7 cells by hepatopancreas and muscle extracts of shrimp fed with T-2 toxin. Toxicol Res. 2017, 6, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Myatt, G.J.; Ahlberg, E.; Akahori, Y.; Allen, D.; Amberg, A.; Anger, L.T.; Aptula, A.; Auerbach, S.; Beilke, L.; Bellion, P.; et al. In silico toxicology protocols. Regul. Toxicol. Pharmacol. 2018, 96, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Lou, C.; Sun, L.; Li, J.; Cai, Y.; Wang, Z.; Li, W.; Liu, G.; Tang, Y. admetSAR 2.0: Web-service for prediction and optimization of chemical ADMET properties. Bioinformatics 2019, 35, 1067–1069. [Google Scholar] [CrossRef] [PubMed]
- Dellafiora, L.; Galaverna, G.; Dall’Asta, C. In silico analysis sheds light on the structural basis underlying the ribotoxicity of trichothecenes—A tool for supporting the hazard identification process. Toxicol. Lett. 2017, 270, 80–87. [Google Scholar] [CrossRef]
- Gratz, S.W.; Dinesh, R.; Yoshinari, T.; Holtrop, G.; Richardson, A.J.; Duncan, G.; MacDonald, S.; Lloyd, A.; Tarbin, J. Masked trichothecene and zearalenone mycotoxins withstand digestion and absorption in the upper gi tract but are efficiently hydrolyzed by human gut microbiota in vitro. Mol. Nutr. Food Res. 2016, 61, 1600680. [Google Scholar] [CrossRef]
- McCormick, S.P.; Kato, T.; Maragos, C.; Busman, M.; Lattanzio, V.M.T.; Galaverna, G.; Dall’Asta, C.; Crich, D.; Price, N.P.J.; Kurtzman, C.P. Anomericity of T-2 Toxin-glucoside: Masked mycotoxin in cereal crops. J. Agric. Food Chem. 2015, 63, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Daud, N.; Currie, V.; Duncan, G.; Busman, M.; Gratz, S.W. Intestinal hydrolysis and microbial biotransformation of diacetoxyscirpenol-α-glucoside, HT-2-β-glucoside and N-(1-deoxy-d-fructos-1-yl) fumonisin B1 by human gut microbiota in vitro. Int. J. Food Sci. Nutr. 2019, 71, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Kasimir, M.; Behrens, M.; Schulz, M.; Kuchenbuch, H.; Focke, C.; Humpf, H.-U. Intestinal metabolism of α- and β-glucosylated modified mycotoxins T-2 and HT-2 toxin in the pig cecum model. J. Agric. Food Chem. 2020, 68, 5455–5461. [Google Scholar] [CrossRef]
- Sang, Y.; Li, W.; Zhang, G. The protective effect of resveratrol against cytotoxicity induced by mycotoxin, zearalenone. Food Funct. 2016, 7, 3703–3715. [Google Scholar] [CrossRef] [PubMed]
- Tatay, E.; Espín, S.; García-Fernández, A.J.; Ruiz, M.-J. Oxidative damage and disturbance of antioxidant capacity by zearalenone and its metabolites in human cells. Toxicol. In Vitro 2017, 45, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.-L.; Sun, X.-F.; Feng, Y.-Z.; Li, B.; Li, Y.-P.; Yang, F.; Nyachoti, C.M.; Shen, W.; Sun, S.-D.; Li, L. Zearalenone exposure impairs ovarian primordial follicle formation via down-regulation of Lhx8 expression in vitro. Toxicol. Appl. Pharmacol. 2017, 317, 33–40. [Google Scholar] [CrossRef]
- Salah-Abbès, J.B.; Belgacem, H.; Ezzdini, K.; Abdel-Wahhab, M.A.; Abbès, S. Zearalenone nephrotoxicity: DNA fragmentation, apoptotic gene expression and oxidative stress protected by Lactobacillus plantarum MON03. Toxicon 2020, 175, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Abid-Essefi, S.; Bouaziz, C.; El Golli-Bennour, E.; Ouanes, Z.; Bacha, H. Comparative study of toxic effects of zearalenone and its two major metabolites α-zearalenol and β-zearalenol on cultured human Caco-2 cells. J. Biochem. Mol. Toxicol. 2009, 23, 233–243. [Google Scholar] [CrossRef]
- Lu, J.; Yu, J.Y.; Lim, S.S.; Son, Y.O.; Kim, D.H.; Lee, S.A.; Shi, X.L.; Lee, J.C. Cellular mechanisms of the cytotoxic effects of the zearalenone metabolites alpha-zearalenol and beta-zearalenol on RAW264.7 macrophages. Toxicol. In Vitro 2013, 27, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Tatay, E.; Meca, G.; Font, G.; Ruiz, M.J. Cytotoxic and interactive effects of zearalenone, alpha-zearalenol and beta-zearalenol and formation of metabolites in HepG2 cells. Rev. Toxicol. 2014, 31, 187–195. [Google Scholar]
- Ennamany, R.; Marzetto, S.; Saboureau, D.; Creppy, E. Lipid peroxidation induced by Boletus satanas: Implication in m5dC variation in Vero cells related to inhibition of cell growth. Cell Biol. Toxicol. 1995, 11, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Vallyathan, V.; Castranova, V.; Shi, X. Cell apoptosis induced by carcinogenic metals. Mol. Cell Biochem. 2001, 222, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Chu, S.H.; Jia, L.S.; Rose, P.; Sabapathy, K.; Schantz, J.T.; Cheung, N.S.; Spencer, J.P.E.; Armstrong, J.S. The pro-inflammatory oxidant hypochlorous acid induces Bax-dependent mitochondrial permeabilisation and cell death through AIF-/EndoG-dependent pathways. Cell. Signal. 2007, 19, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Bouaziz, C.; Sharaf el dein, O.; El Golli, E.; Abid-Essefi, S.; Brenner, C.; Lemaire, C.; Bacha, H. Different apoptotic pathways induced by zearalenone, T-2 toxin and ochratoxin A in human hepatoma cells. Toxicology 2008, 254, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Ben Salem, I.; Prola, A.; Boussabbeh, M.; Guilbert, A.; Bacha, H.; Lemaire, C.; Abid-Essefi, S. Activation of ER stress and apoptosis by α- and β-zearalenol in HCT116 cells, protective role of Quercetin. Neurotoxicology 2016, 53, 334–342. [Google Scholar] [CrossRef]
- Dellafiora, L.; Perotti, A.; Galaverna, G.; Buschini, A.; Dall’Asta, C. On the masked mycotoxin zearalenone-14-glucoside. Does the mask truly hide? Toxicon 2016, 111, 139–142. [Google Scholar] [CrossRef]
- Cirlini, M.; Barilli, A.; Galaverna, G.; Michlmayr, H.; Adam, G.; Berthiller, F.; Dall’Asta, C. Study on the uptake and deglycosylation of the masked forms of zearalenone in human intestinal Caco-2 cells. Food Chem. Toxicol. 2016, 98, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Catteuw, A.; Broekaert, N.; De Baere, S.; Lauwers, M.; Gasthuys, E.; Huybrechts, B.; Callebaut, A.; Ivanova, L.; Uhlig, S.; De Boevre, M.; et al. Insights into in vivo absolute oral bioavailability, biotransformation, and toxicokinetics of zearalenone, α-zearalenol, β-zearalenol, zearalenone-14-glucoside, and zearalenone-14-sulfate in pigs. J. Agric. Food Chem. 2019, 67, 3448–3458. [Google Scholar] [CrossRef]
- Yang, S.; Li, Y.; De Boevre, M.; De Saeger, S.; Zhou, J.; Li, Y.; Zhang, H.; Sun, F. Toxicokinetics of α-zearalenol and its masked form in rats and the comparative biotransformation in liver microsomes from different livestock and humans. J. Hazard. Mater. 2020, 393, 121403. [Google Scholar] [CrossRef]
- Catteuw, A.; Devreese, M.; De Baere, S.; Antonissen, G.; Huybrechts, B.; Ivanova, L.; Uhlig, S.; Martens, A.; De Saeger, S.; De Boevre, M.; et al. Toxicokinetic studies in piglets reveal age-related differences in systemic exposure to zearalenone, zearalenone-14-glucoside, and zearalenone-14-sulfate. J. Agric. Food Chem. 2020, 68, 7757–7764. [Google Scholar] [CrossRef]
- Binder, S.B.; Schwartz-Zimmermann, H.E.; Varga, E.; Bichl, G.; Michlmayr, H.; Adam, G.; Berthiller, F. Metabolism of zearalenone and its major modified forms in pigs. Toxins 2017, 9, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, J.; Borzekowski, A.; Haase, H.; Menzel, R.; Rueß, L.; Koch, M. Toxicity assay for citrinin, zearalenone and zearalenone-14-sulfate using the nematode Caenorhabditis elegans as model organism. Toxins 2018, 10, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luongo, D.; Severino, L.; Bergamo, P.; De Luna, R.; Lucisano, A.; Rossi, M. Interactive effects of fumonisin B1 and alpha-zearalenol on proliferation and cytokine expression in Jurkat T cells. Toxicol. In Vitro 2006, 20, 1403–1410. [Google Scholar] [CrossRef]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Tudor, D.S. Effects of zearalenone and its derivatives on the innate immune response of swine. Toxicon 2010, 56, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Manda, G.; Motiu, M.; Neagoe, I.; Dragomir, C.; Stancu, M.; Calin, L. Effects of zearalenone and its derivatives on porcine immune response. Toxicol. In Vitro 2011, 25, 1981–1988. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain. Scientific opinion on the appropriateness to set a group health-based guidance value for zearalenone and its modified forms. EFSA J. 2016, 14, 4425. [Google Scholar]
- Tatay, E.; Espín, S.; García-Fernández, A.-J.; Ruiz, M.-J. Estrogenic activity of zearalenone, α-zearalenol and β-zearalenol assessed using the E-screen assay in MCF-7 cells. Toxicol. Mech. Methods 2018, 28, 239–242. [Google Scholar] [CrossRef]
- Ben Salem, I.; Boussabbeh, M.; Da Silva, J.P.; Guilbert, A.; Bacha, H.; Abid-Essefi, S.; Lemaire, C. SIRT1 protects cardiac cells against apoptosis induced by zearalenone or its metabolites α- and β-zearalenol through an autophagy-dependent pathway. Toxicol. Appl. Pharmacol. 2017, 314, 82–90. [Google Scholar] [CrossRef]
- Steinkellner, H.; Binaglia, M.; Dall’Asta, C.; Gutleb, A.C.; Metzler, M.; Oswald, I.P.; Parent-Massin, D.; Alexander, J. Combined hazard assessment of mycotoxins and their modified forms applying relative potency factors: Zearalenone and t2/ht2 toxin. Food Chem. Toxicol. 2019, 131, 110599. [Google Scholar] [CrossRef]
- Bittner, G.D.; Denison, M.S.; Yang, C.Z.; Stoner, M.A.; He, G. Chemicals having estrogenic activity can be released from some bisphenol a-free, hard and clear, thermoplastic resins. Environ. Health 2014, 13, 103. [Google Scholar] [CrossRef] [Green Version]
- Eze, U.A.; Huntriss, J.; Routledge, M.N.; Gong, Y.Y.; Connolly, L. The effect of individual and mixtures of mycotoxins and persistent organochloride pesticides on oestrogen receptor transcriptional activation using in vitro reporter gene assays. Food Chem. Toxicol. 2019, 130, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Dellafiora, L.; Ruotolo, R.; Perotti, A.; Cirlini, M.; Galaverna, G.; Cozzini, P.; Buschini, A.; Dall’Asta, C. Molecular insights on xenoestrogenic potential of zearalenone-14-glucoside through a mixed in vitro/in silico approach. Food Chem. Toxicol. 2017, 108, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Poppenberger, B.; Berthiller, F.; Bachmann, H.; Lucyshyn, D.; Peterbauer, C.; Mitterbauer, R.; Schuhmacher, R.; Krska, R.; Glössl, J.; Adam, G. Poppenberger i wsp. Heterologous expression of Arabidopsis UDP-glucosyltransferases in Saccharomyces cerevisiae for production of zearalenone-4-O-glucoside. Appl. Env. Microbiol. 2006, 72, 4404–4410. [Google Scholar] [CrossRef] [Green Version]
- Mallebrera, B.; Juan-Garcia, A.; Font, G.; Ruiz, M.J. Mechanisms of beauvericin toxicity and antioxidant cellular defense. Toxicol. Lett. 2016, 246, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Tatay, E.; Font, G.; Ruiz, M.J. Cytotoxic effects of zearalenone and its metabolites and antioxidant cell defense in CHO-K1 cells. Food Chem. Toxicol. 2016, 96, 43–49. [Google Scholar] [CrossRef]
- Foryst-Ludwig, A.; Clemenz, M.; Hohmann, S.; Hartge, M.; Sprang, C.; Frost, N.; Krikov, M.; Bhanot, S.; Barros, R.; Morani, A.; et al. Metabolic actions of estrogen receptor beta (ERβ) are mediated by a negative cross-talk with PPARγ. PLoS Genet. 2008, 4, e1000108. [Google Scholar] [CrossRef]
- Karaman, E.F.; Ozden, S. Alterations in global DNA methylation and metabolism-related genes caused by zearalenone in MCF7 and MCF10F cells. Mycotoxin Res. 2019, 35, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.N.; Khan, A.U. Role of histone acetylation in cell physiology and diseases: An update. Clin. Chim. Acta 2010, 411, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Ma, C.; Chen, L.; Luo, D.; Chen, R.; Liang, F. Mechanistic insights into the interaction between transcription factors and epigenetic modifications and the contribution to the development of obesity. Front. Endocrinol. 2018, 9, 370. [Google Scholar] [CrossRef] [Green Version]
- Tailleux, A.; Wouters, K.; Staels, B. Roles of PPARs in NAFLD: Potential therapeutic targets. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2012, 1821, 809–818. [Google Scholar] [CrossRef] [PubMed]
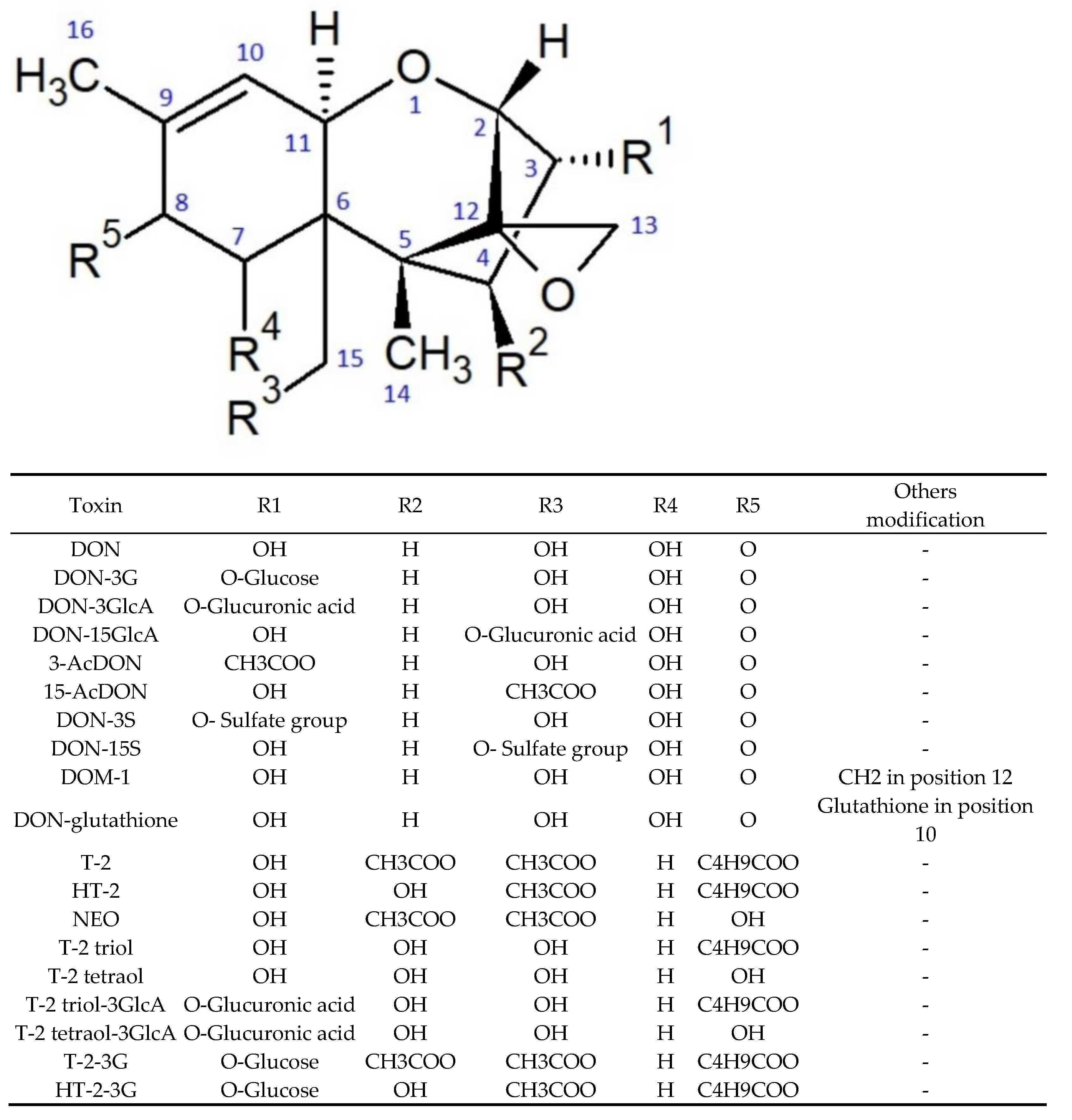

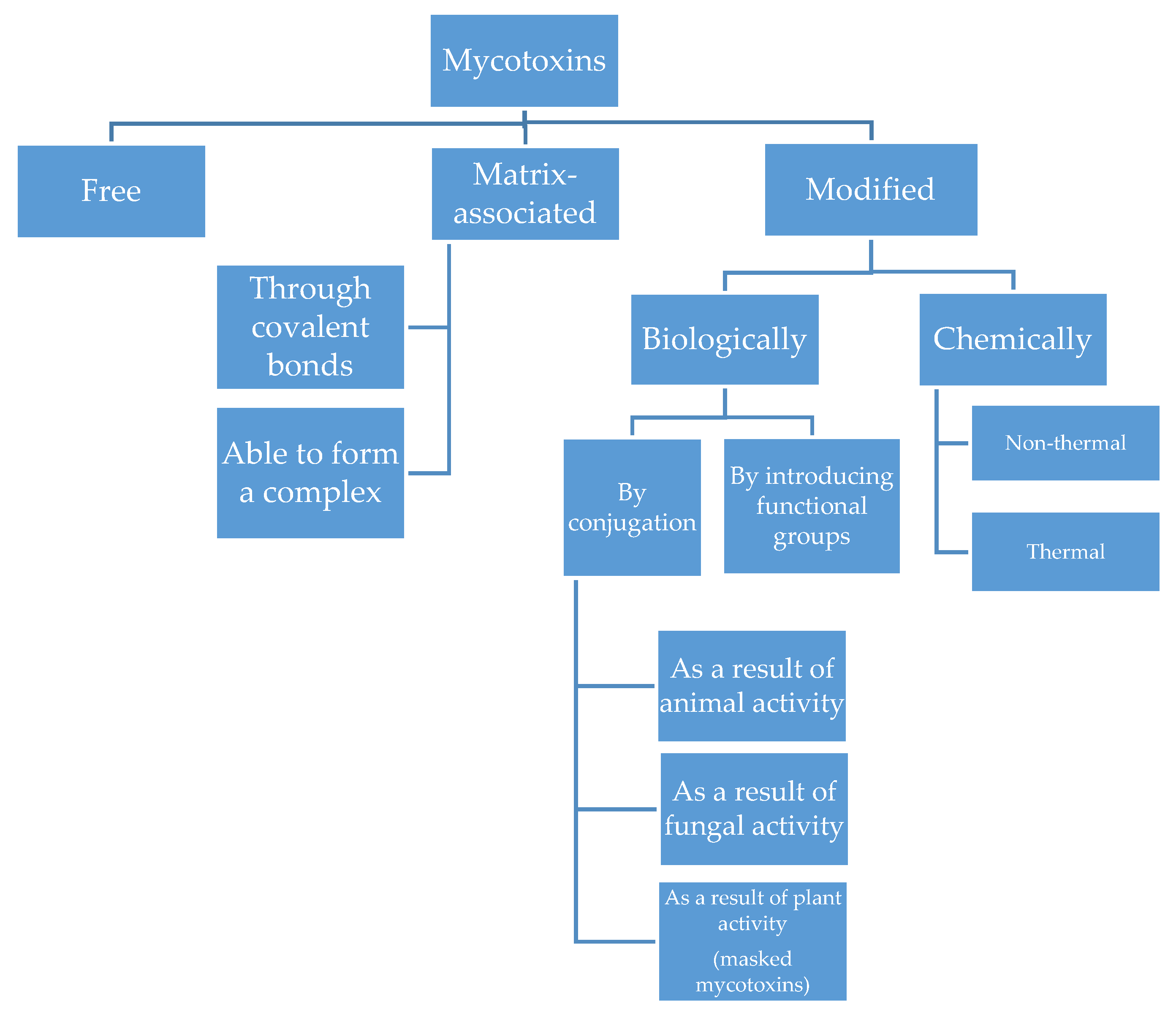

{kind=link}
{kind=link}
{kind=link}
| Unprocessed Cereals | Maximum Levels (μg/kg) | Source |
|---|---|---|
| DON | ||
| Cereals other than durum wheat, oat, maize | 1250 | [7,8] |
| Oat, maize, durum wheat | 1750 | |
| ZEN | ||
| Cereals other than maize | 100 | [7,8] |
| Maize | 350 | |
| Sum of T-2 and HT-2 | ||
| Oat | 1000 | [9] |
| Barley, maize | 200 | |
| Wheat, rye and other cereals | 100 | |
| Tested Animals | Tested Toxins | Exposure Type | Dose | Main Observations | Source |
|---|---|---|---|---|---|
| Pigs | DON DOM-1 | Orally | 0.5 nmol/kg BW for 21 days 1 mol/kg BW for 14 days | DOM-1 does not cause vomiting, body weight changes, or pathological changes in intestines and liver | [37] |
| DON DOM-1 3-epi-DON | Orally | Unrestricted access to feed with toxin content of 3 mg/kg | DON and DOM-1, 3-epi-DON do not cause vomiting, body weight changes, or pathological changes in intestines and liver | [38] | |
| DON DON-3G | Orally and intravenously | 55.7 µg/kg BW for DON-3G 36 µg/kg BW for DON | DON-3G does not undergo hydrolysis within the circulatory system and is not absorbed when administered orally | [89,90] | |
| Chickens Broilers | DON DON-3G | Orally and intravenously | 500 µg/kg BW for DON 774 µg/kg BW for DON-3G | DON-3G does not undergo hydrolysis within the circulatory system and gastrointestinal tract | [90] |
| Cell Line | Tested Toxins | Concentration Range and Exposure Time | Methodology | Main Conclusions | Source |
|---|---|---|---|---|---|
| HEPG2 | DON 3-AcDON 15-AcDON | 0–12.5 (µM) for 24 h | MTT | IC50 values (µM) were 4.3, 6.2, and 8.1 for DON, 3-AcDON, and 15-AcDON, respectively | [30] |
| H2-DCFDA applied and fluorescence measured, TBARS applied and absorbance measured | Increase in ROS level was observed upon exposure to 15-AcDON. Lipid peroxidation was observed upon exposure to DON, 3-AcDON, and 15-AcDON | ||||
| 0–7.5 (µM) 3-AcDON, 15-AcDON 0–15 (µM) DON for 24 h | Neutral red assay | IC50 values (µM) were 3.90, 6.00, and 10.15 for 3-AcDON, 15-AcDON, and DON, respectively | [39] | ||
| 0–4.8 (µM) DON 0–3 (µM) 3-AcDON, 15-AcDON for 48 h | Flow cytometry | Toxins have the ability to disturb the cell cycle and induce micronucleus formation | |||
| GES-1 | DON 3-AcDON 15-AcDON DON-3G | 0–3 (ppm) DON, 15-AcDON 0–12 (ppm) 3-AcDON, DON-3G for 24 h | Cell counting kit-8 (DOJINDO, Kumamoto, Japan) | The following toxicity ranking was proposed: DON >15-AcDON >>3-AcDON >DON-3G | [31] |
| DON 15-AcDON | 0–5 (μM) for 8 h | High-performance liquid chromatography-tandem high-resolution mass | Proving metabolic balance disturbances upon exposure to DON, 15-AcDON | [41] | |
| Western blot | Induction of apoptosis by DON and 15-AcDON was observed as a result of the activation of mitogene-activated kinases (MAPK) p38 and JNK and ERK1/2 kinases inhibition | ||||
| 5 (μM) for 30 min for the ROS assay 5 (μM) for 24 h for ATP and NAD+/NADH measurements | DCF-DA applied and fluorescence measured ATP Assay Kit NAD+/NADH Assay Kit | Increase in ROS level Decrease in ATP level and NAD+/NADH balance disturbances upon exposure to DON and 15-AcDON. | |||
| IPEC-J2 | DON 3-AcDON 15-AcDON DON-3G | 0–20 µg/ml for 72 h | Flow cytometry | Following toxicity ranking: 15-AcDON ≈DON >3-AcDON >>DON-3G was drawn | [25] |
| DON DOM-1 | 0–100 (µM) for 24, 48, and 72 h | NR, SRB, LDH, WST-1, MTT, CTG | DOM-1 showed no cytotoxicity | [88] | |
| CACO-2 | DON DON-3G | 0–10 (µM) for 48 h | CellTiter-Glo Luminescent Cell Viability Assay (Promega, Madison, USA) | DON-3G showed no cytotoxicity | [87] |
| DON 3-AcDON 15-AcDON DON-3G DOM-1 | 0–8.4 (µM) for 24 h for LDH assay for 4, 8, 12, and 24 h for TEER assays | LDH, TEER assay | Lower 3-AcDON cytotoxicity compared with that of 15-AcDON and DON DON-3G and DOM-1 showed no cytotoxicity | [82] | |
| DON 3-AcDON 15-AcDON | 0–0.5 (µM) for 24 h | RT-PCR | Inhibition of cell cycle by DON, 3-AcDON, and 15-AcDON as a result of ATM kinase activation | [43] | |
| 0–10 (µM) for 24, 48, and 72 h | Cell-counting kit-8 (Shanghai, China) | Toxicity ranking below reported: 15-AcDON ≈DON >3-AcDON | |||
| PBMC from bovine, pig and chicken sources | DON DOM-1 | 0–3.37 (µM) for 28 h DON 0–357 (µM) for 72 h DOM-1 | Bromodeoxyuridine assay (BrdU) | DOM-1 at a concentration of 357 (µM) inhibits proliferation of PBMC obtained from bovine, pig and chicken sources | [97] |
| Tested Animals | Tested Toxins | Exposure Type | DOSE | Main Observations | Source |
|---|---|---|---|---|---|
| Pigs | ZEN-14G ZEN-14S | Intravenously/orally | 500 μg/kg BW for ZEN-14G 415 μg/kg BW for ZEN-14S | ZEN-14G may hydrolyse in the circulatory system. ZEN-14G and ZEN-14S are fully hydrolysed within the GI tract | [132] |
| ZEN-14G ZEN-14S | Orally | 15.1 µg/kg BW for ZEN-14G 12.5 µg/kg BW for ZEN-14S | No detectable quantities of tested toxins were found in urine or faeces | [135] | |
| Rats | α-ZOL α-ZOL-14G | Intravenously/orally | 0.5 mg/kg BW for α-ZOL 0.75 mg/kg BW for α-ZOL-14G | Efficient conversion of α-ZOL-14G into ZEN. Low bioavailability of oral α-ZOL and α-ZOL-14G after oral administration | [133] |
| Caenorhabditis elegans (nematodes) | ZEN ZEN-14S | Medium culture containing mycotoxins | ZEN: 24; 228 (µM) ZEN-14S: 19; 95 (µM) | Comparable reduction in the offspring number of nematodes by ZEN and ZEN-14S | [136] |
| Cell Line | Tested Toxins | Applied Concentrations | Methodology | Main Conclusions | Source |
|---|---|---|---|---|---|
| HEPG2 | ZEN α-ZOL β-ZOL | 0–100 (µM) for 72 h | Neutral red assay | Established IC50 values(μM): 13.1 for β-ZOL 39.7 forZEN 119 for α-ZOL | [35] |
| Qiagen RNeasy midi kit (QIAGEN GmbH, Germany) | IL-1β, IL-8, and TNF-α expression were inhibited by ZEN, α-ZOL, and β-ZOL | ||||
| ZEN α-ZOL | 0–250 (μM) for 24 h | MTT | Established IC50 values (µM): 131.40 for α-ZOL 143.35 forZEN | [36] | |
| ZEN α-ZOL β-ZOL | 0–100 (μM) for: 24, 48, and 72 h | MTT | Toxicity ranking: α-ZOL >β-ZOL >ZEN | [124] | |
| ZEN α-ZOL β-ZOL | 0–25 (μM) for 2 h | Fluorescence measured using dichlorofluorescein | Induction of ROS formation by ZEN, α-ZOL, and β-ZOL at all concentrations used | [119] | |
| Comet assay | Dose-dependent induction of DNA damage by ZEN, α-ZOL, and β-ZOL | ||||
| Spectrophotometry, Ransod (Randox Laboratories, UK) | Increase in the activities of SOD and GPx, decrease in CAT activity upon exposure to ZEN, α-ZOL, and β-ZOL | ||||
| ZEN α-ZOL | 0–50 (μM) for 24 h | Western blot | Increased activities of methyltransferase and acetyltransferase. Increased expression of genes coding components of metabolic pathways and nuclear receptors. | [36] | |
| CACO-2 | ZEN α-ZOL β-ZOL | 0–100 (μM) for 48 h | MTT | Established IC50 values (µM): 20 for ZEN 60 forβ-ZOL 80 forα-ZOL | [122] |
| ZEN-14G | 0–40 (μM) for 6 h | Resazurin dyeing | No ZEN-14G cytotoxicity found | [131] | |
| SH-SY5Y | α-ZOL β-ZOL | 0–100 (μM) for 72 h | MTT | Established IC50 values (μM): 7.5 for β-ZOL 14 for α-ZOL | [48] |
| 0–12.5 (μM) for: 24, 48, and 72 h | Synergy in the induction of toxic effect found for the mixture of α-ZOL and β-ZOL | ||||
| Neutrophils isolated from porcine peripheral blood | ZEN α-ZOL β-ZOL ZAN | 0–50 (μM) for 1 h | MTT | Established IC50 values (μM): 53.1 for ZAN 56.8 for β-ZOL 59.0 for α-ZOL 73.4 for ZEN | [138] |
| 0–10 (μM) for 3 h | ELISA | IL8 expression in neutrophils reduction caused by ZEN, α-ZOL, β-ZOL, and ZAN | |||
| PBMC isolated from porcine peripheral blood | ZEN α-ZOL β-ZOL ZAN | 0–100 (μM) for 48 h | MTT | Established IC50 values (μM): 17.3 for β-ZOL 22.7 for ZEN 26.3 for ZAN 29.1 for α-ZOL | [139] |
| 0–10 (μM) for 7 days | ELISA | ZEN, α-ZOL, β-ZOL, and ZAN show the ability to decrease production of antibodies in classes: IgG, IgA, and IgM | |||
| RAW264.7 | α-ZOL β-ZOL | 0–50 (μM) for 24 h | WST-8 | Higher cytotoxicity of β-ZOL, compared with that of α-ZOL. | [123] |
| Flow cytometry | α-ZOL and β-ZOL induce cell death to a greater extent by apoptosis than by necrosis | ||||
| Flow cytometry Western blot | α-ZOL and β-ZOL induce apoptosis independently of caspases through mitochondrial stress | ||||
| MCF-7 | ZEN-14G | 0–1 (μM) for 6 h | MTS | ZEN-14G shows no cytotoxicity | [130] |
| ZEN α-ZOL β-ZOL | 0–25 (µM) for 6 days | E-Screen | Oestrogen activity displayed by ZEN, α-ZOL, and β-ZOL | [141] | |
| CHO-K1 | ZEN α-ZOL β-ZOL | 0–25 (μM) for 2 h | Fluorescence measured using dichlorofluorescin | Induction of ROS formation by ZEN, α-ZOL, and β-ZOL at all concentrations tested | [149] |
| Comet assay | Dose-dependent induction of DNA damage by ZEN, α-ZOL, and β-ZOL | ||||
| Spectrophotometry, Ransod (Randox Laboratories, UK) | SOD and GPx activity increase, CAT activity decreases upon exposure to ZEN, α-ZOL, and β-ZOL | ||||
| Jurkat T Cells | α-ZOL | 0–80 (μM) for 24 h | RT–PCR | α-ZOL inhibits expression of IL-2 and IFNγ in a T cell culture | [137] |
| 3H-thymidine incorporation measurement | T cell proliferation inhibited by α-ZOL | ||||
| HCT116 | α-ZOL β-ZOL | 180 (mM) α-ZOL 300 (mM) β-ZOL for 24 h | qRT-PCR | Presence of endoplasmic reticulum stress markers identified upon cell exposure to α-ZOL or β-ZOL | [129] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pierzgalski, A.; Bryła, M.; Kanabus, J.; Modrzewska, M.; Podolska, G. Updated Review of the Toxicity of Selected Fusarium Toxins and Their Modified Forms. Toxins 2021, 13, 768. https://doi.org/10.3390/toxins13110768
Pierzgalski A, Bryła M, Kanabus J, Modrzewska M, Podolska G. Updated Review of the Toxicity of Selected Fusarium Toxins and Their Modified Forms. Toxins. 2021; 13(11):768. https://doi.org/10.3390/toxins13110768
Chicago/Turabian StylePierzgalski, Adam, Marcin Bryła, Joanna Kanabus, Marta Modrzewska, and Grażyna Podolska. 2021. "Updated Review of the Toxicity of Selected Fusarium Toxins and Their Modified Forms" Toxins 13, no. 11: 768. https://doi.org/10.3390/toxins13110768
APA StylePierzgalski, A., Bryła, M., Kanabus, J., Modrzewska, M., & Podolska, G. (2021). Updated Review of the Toxicity of Selected Fusarium Toxins and Their Modified Forms. Toxins, 13(11), 768. https://doi.org/10.3390/toxins13110768