Cyanotoxins and Food Contamination in Developing Countries: Review of Their Types, Toxicity, Analysis, Occurrence and Mitigation Strategies

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Common Cyanotoxins: Classification, Accumulation, and Toxicity
2.1. Toxic Cyclic Peptides
2.1.1. Microcystins
2.1.2. Nodularins
2.2. Toxic Alkaloids
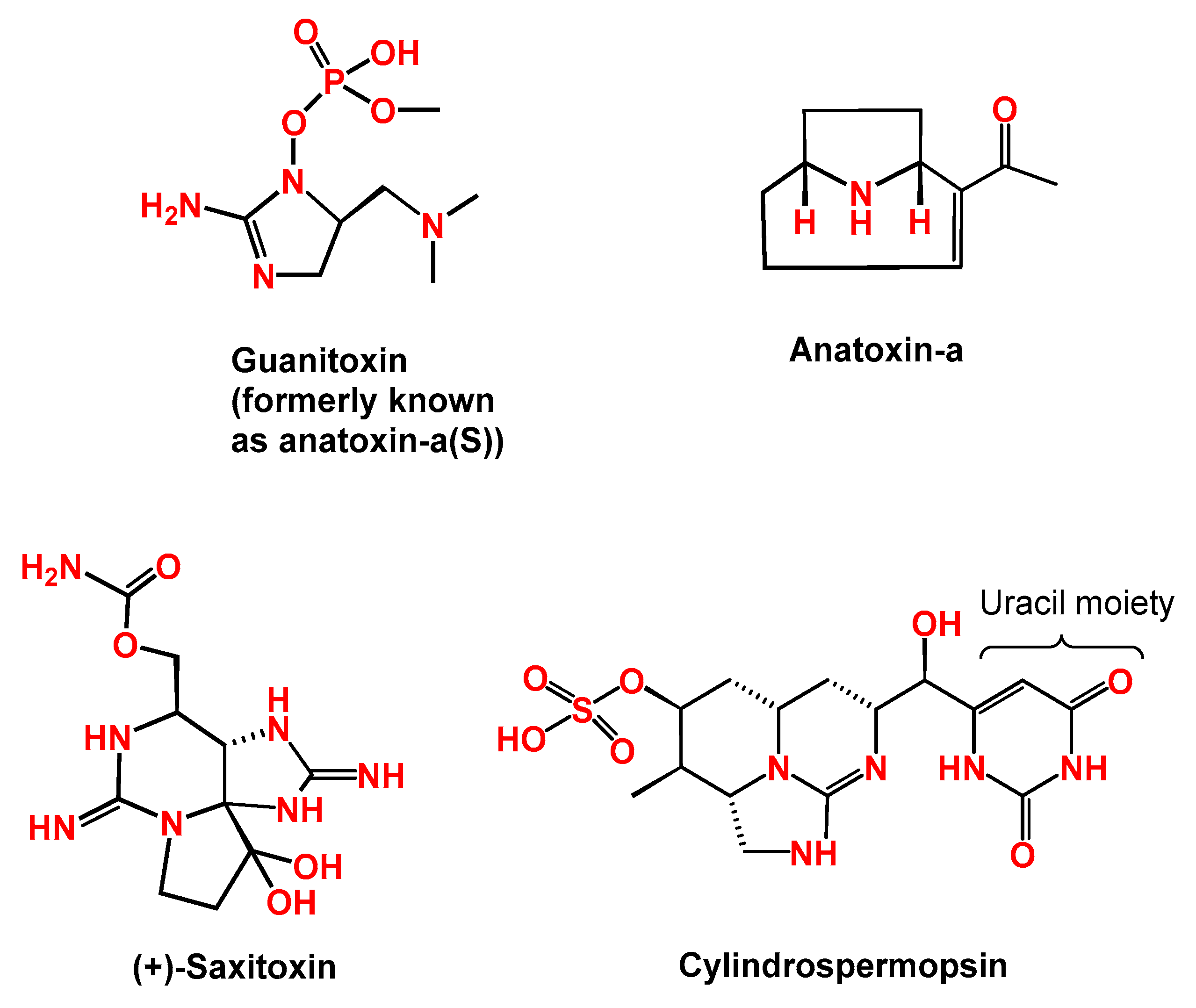
2.2.1. Anatoxin-a and Anatoxin-a(s)
2.2.2. Cylindrospermopsin
2.2.3. Saxitoxins (STXs)
2.3. Lipopolysaccharides
3. Analysis of Cyanotoxins in Food
4. Occurrence of Cyanotoxins in Food from Developing Countries
4.1. Occurrence of Cyanotoxins in Seafood from the Developing Countries of Africa

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Location/Year | Detected Toxins and Concentration Range (μg/kg, DW or WW) | Matrix | N | P | Method of Detection | References |
|---|---|---|---|---|---|---|---|
| Algeria | Lake Oubeir/2010–2011 | MCs (329–680, DW) | Cyprinus carpio (muscles) | 36 | 36 | PP2A inhibition assay | [123] |
| Egypt | Sohag city/2000 | MCs (45.7–102, WW) | Oreochromis niloticus (muscles) | NM | NM | ELISA | [52] |
| Sohag city/2012–2013 | CYN (19–280, WW) | Oreochromis niloticus (muscles) | 99 | 63 | ELISA | [79] | |
| Sohag city/2012–2013 | Free MCs (0.02–0.38, WW); Bound MCs (15,000–19,000, WW) | Oreochromis niloticus (muscles) | 198 | NM | ELISA & Lemieux oxidation reactions and LC-PDA | [50] | |
| Ethiopia | Lake Hora-Arsedi/2015 | MCs <LOD | Oreochromis niloticus; Tilapia zillii (muscles) | 8 | Not detected | LC-HRMS | [129] |
| Addis Ababa/2015–2016 | MCs <LOD | Oreochromis niloticus; Cyprinus Carpio; Clarias gariepinus (muscles) | 36 | Not detected | LC-HRMS | [124] | |
| Kenya | Nyanza Gulf, Lake Victoria/2011-2012 | MC-YR (8–20, DW) | Rastrineobola argentea (whole small fish) | 16 | 4 | LC-MS/MS | [125] |
| South Africa | Mpumalanga province/2012 | MC-LR (0.17–0.19, WW) | Labeo rosae; Oreochromis mossambicus (muscles) | 26 | 21 | ELISA | [126] |
| Uganda | Lake Mburo and Murchison Bay, Lake Victoria/2004–2005 | MCs (5–121, WW) | Oreochromis niloticus (muscles) | 72 | 51 | LC-MS/MS | [122] |
| Lakes (Victoria, Albert, Edward, George, Mburo, Nkuruba, Saka)./2007–2009 | MCs (0.5–1917,WW) | 19 species of fish (muscles) | 399 | NM | ELISA | [53] | |
| Lake Mburo & Murchison Bay, Lake Victoria/2004–2005 | MCs (9.6–208.6, WW) | Oreochromis niloticus (muscles) | 24 | 17 | LC-MS/MS | [121] |
4.2. Occurrence of Cyanotoxins in Seafood from the Developing Countries in Asia
| Country | Location/ Year | Detected Toxins and Concentration Ranges (μg/kg, DW or WW) | Matrix | N | P | Method of Detection | References |
|---|---|---|---|---|---|---|---|
| China | Tianjin/2017 | MC-RR (up to 290, DW); MC-LR (up to120, DW) | Carassius auratus (muscles) | 21 | NM | LC-PDA | [153] |
| Lake Taihu/2005 | MC-RR (up to 2, DW) | Hypophthalmichthys molitrix (muscles) | 9 | NM | LC-MS/MS | [6] | |
| Lake Taihu/2004–2005 | MCs (up to 887, DW) | Aristichthys nobilis (muscles) | 48 | NM | LC-MS/MS | [135] | |
| Lake Taihu/2003–2004 | MCs (70.6–584, DW) | Anodonta woodiana; Hyriopsis cumingii; Cristaria plicata; Lamprotula leai (muscles) | 28 | 15 | LC-UV | [134] | |
| Lake Chaohu, Anhui Province/2003 | MC-LR and -RR (up to 530, DW) | Palae-monetes; Macrobrachium nipponensis (shrimps) | 11 | 5 | LC-UV | [51] | |
| Lake Taihu/2004–2005 | MCs (up to 1244, DW) | Hypophthalmichthys molitrix (muscles) | 12 | 11 | LC-MS/MS | [133] | |
| Lake Chaohu, Anhui Province/2003 | MC-LR and -RR (up to 180, DW) | Corbicula fluminea (muscles) | 7 | 4 | LC-UV | [132] | |
| China’s Tiesha River, Hangzhou/NM | MCs (up to 2860, WW) | Hypophthalmichthys molitrix (muscles) | 3 | 3 | LC-MS/MS | [154] | |
| Lake Taihu/2011 | MCs (25.5–31.7, DW) | Hypophthalmichthys molitrix; Aristichthys nobilis; Carassius carassius; Cyprinus carpio (muscles) | 46 | NM | LC-MS/MS | [138] | |
| Lake Chaohu, Anhui Province/2012 | MCs (4.3–27.9, DW) | Hypophthalmichthys molitrix; Carassius auratus (muscles) | NM | NM | ELISA | [139] | |
| Lake Chaohu, Anhui Province/2014 | MCs (1.3–350, DW) | Hypophthalmichthys molitrix; Carassius auratus (muscles) | 60 | NM | ELISA | [140] | |
| WJD Reservoir, Guizhou/2016–2017 | MC-LR (1.0–450,WW) and MC-RR (1.7–678, WW) | C. idellus; P. pekinensis; H. molitrix; A. nobilis; C. carpio; C. auratus; T. tinca; M. piceus; L. japonicas; P. asotus (muscles) | 205 | 106 (MC-LR), 110 (MCs-RR) | LC-PDA | [141] | |
| Fish pond, Hangzhou/2009 | MC-LR (240, DW) | Aristichthys nobilis (muscles) | 1 | 1 | LC-MS/MS | [155] | |
| Lake Taihu and Lake Chaohu/2009–2010 | MCs (18.6–48.2, DW) | Nine fish species (muscles) | NM | NM | ELISA | [136] | |
| Eight different lakes in four provinces (Hubei, Hunan, Jiangsu and Anhui)/2008 | MCs (14–36, DW) | Hypophthalmichthys molitrix (muscles) | 24 | 24 | LC-MS/MS | [143] | |
| Lake Dianchi/2008 | MCs (up to 1360, DW) | Margarya melanioides (whole snail) | 6 | 6 | ELISA | [156] | |
| Lake Chaohu, Anhui Prov-ince/2003 | MC-LR (up to 2290, DW) | Hypophthalmichthys molitrix; Carassius auratus; Cyprinus carpio; Coilia ectenes (muscles) | 8 | 4 | LC-UV | [157] | |
| Lake Taihu/2005 | MCs (13–170, DW) | Hypophthalmichthys molitrix; Cyprinus carpio; Carassius auratus (muscles) | NM | NM | LC-MS/MS | [137] | |
| India | Lakshmikund pond, Varanasi/2011-2012 | MC-LR (1.44–3.01, DW); MC-RR (5.94–65.03, DW); MC-YR (1.39–1.71, DW) | Cyprinus carpio; Clarias batrachus (muscles) | 3 | 3 | LC-MS/MS | [147] |
| Iran | Anzali wetland, Guilan Province/2014 | MC-LR (10.1–40.9, WW) | Hypophthalmichthys molitrix; Esox lucius (muscles) | 18 | NM | LC-UV | [148] |
| Thailand | Fish ponds, Bang Bo District, Samut Prakan Province/NM | MC-LR < LOD | Clarias macrocephalus (muscles) | 720 | - | LC-MS/MS | [158] |
| Phayao Lake, Phayao Province/2011 | MC-LR (up to 0.26, DW) | Oreochromis niloticus (muscles) | 24 | 7 | LC-UV | [159] | |
| Experimental fish pound, Chiang Mai/NM | MC-LR (8.3–14.4, DW) | Oreochromis niloticus (muscles) and Macrobrachium Rosenbergii (whole prawns) | 120 | NM | ELISA | [149] | |
| Experimental fish pound, Chiang Mai/2006-2007 | MC-LR (0.84 to 3.20, DW) | Tilapia nilotica (muscles) and Macrobrachium Rosenbergii (whole prawns) | 30 | NM | ELISA | [150] | |
| Turkey | Lake Eğirdir, Turkey/2013 | MCs (0.36–17.54, DW) | Cyprinus carpio; Cyprinus gibelio; Atherina boyeri (muscles) | 110 | 51 | ELISA | [144] |
| Vietnam | Mekong basin, South Vietnam/ 2015–2016 | MC-dmLR (150–300, DW) | Pomacea canaliculate (muscles) | 68 | 3 | LC-MS/MS | [152] |
| Phuoc Ninh/2011 | MCs (1150–2370, DW) | Corbicula sp.; Ensidens sp. (whole bivalves) | 200 | 200 | LC-PDA | [151] |
4.3. Occurrence of Cyanotoxins in Food from the Developing Countries in Latin American
| Country | Location/ Year | Detected Toxins and Concentration Ranges (μg/kg, DW or WW) | Matrix | N | P | Method of Detection | References |
|---|---|---|---|---|---|---|---|
| Argentina | Los Padres Lake/2007 | MCs (1.0–8.0, WW) | Odontesthes bonariensis (muscles) | 20 | NM | LC-MS/MS | [169] |
| San Roque reservoir/2004 | MC-RR (4.8–340, WW) | Odontesthes bonariensis (muscles) | 30 | NM | LC-UV | [168] | |
| San Roque Reservoir/2011 | NODs (80–90, WW) | Palaemonetes argentines (whole shrimp) | 80 | NM | LC-MS/MS | [66] | |
| Brazil | Rio de Janeiro/2003 | MCs (0.9–12, WW) | Oreochromis niloticus; Tilapia rendalli (muscles) | 27 | 27 | ELISA | [164] |
| Jacarepaguá lagoon/2012 | MCs (up to 4, WW) | Oreochromis niloticus (muscles) | 17 | 10 | ELISA | [171] | |
| Alagados Reservoir/2007–2008 | GTXs (22.1–30.6, WW) | Geophagus brasiliensis (muscles) | 64 | 64 | LC-FLD | [95] | |
| Paranoa Lake/2006–2007 | MC-LR (up to 3380, WW) | Hypophthalmichthys molitrix (muscles) | 78 | NM | LC-PDA | [163] | |
| Sepetiba Bay, Rio de Janeiro/1999 | MCs (up to 103.3, WW) | Fish muscles; shrimp; crab | 33 | 16 | ELISA | [166] | |
| Rio de Janeiro/1996–1999 | MCs (1.6–337.3, WW) | Tilapia rendalli (muscles) | 144 | 108 | ELISA | [54] | |
| Garça city/2005 | STXs (up to 22, WW) | Oreochromis niloticus (muscles) | 24 | NM | LC-FLD | [93] | |
| Iraí Reservoir, Paraná State/2015 | MCs (up to 7, WW) | Geophagus brasiliensis (muscles) | 50 | NM | LC-MS/MS | [162] | |
| Guatemala | Lake Amatitlán cost/NM | MC-RR (up to 1.03 and 1.16, DW) | Solanum lycopersicum (Tomato); Capsicum annuum (peppers) | 12 | 12 | LC/MS/MS | [24] |
| Mexico | Lago Catemaco, Veracruz/NM | CYN (up to 3.35, WW); STXs (up to 1.04, WW) | Pomacea patula catemacensi (snails) | NM | NM | ELISA | [94] |
| Lake Catemaco, Veracruz/2009 | CYN (0.09–1.26, WW); STXs (0.03–0.71, WW) | Rhamidia sp.; Oreochromis aureus; Vieja sp.; V. fenestrate; Heterandria Jonesii; Bramocharax cabelleroi; Cichlasoma urophtalmus; C. helleri; Dorosoma mexicana (Fish tissue) | 14 | 14 | ELISA | [81] | |
| Michoacan/2008–2009 | MCs (4.99–157, WW) | Cyprinus carpio sp.; Goodea sp. (Fish tissue) | NM | NM | ELISA | [167] | |
| Lake Zumpango, Mexico City/2016–2017 | MCs (5–24, DW) | Chirostoma jordan (muscles) | 30 | 30 | ELISA | [55] |
5. Regulation and Mitigation of Cyanotoxins
5.1. Management of Nutrients
5.2. Chemical and Physical Approaches
5.3. Biological Approach
5.4. Public Awareness
6. Conclusions and Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dittmann, E.; Wiegand, C. Cyanobacterial toxins—Occurrence, biosynthesis and impact on human affairs. Mol. Nutr. Food Res. 2006, 50, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, D.; Xie, P.; Wang, Q.; Ma, Z. Simultaneous determination of microcystin contaminations in various vertebrates (fish, turtle, duck and water bird) from a large eutrophic Chinese lake, Lake Taihu, with toxic Microcystis blooms. Sci. Total Environ. 2009, 407, 3317–3322. [Google Scholar] [CrossRef] [PubMed]
- Žegura, B.; Štraser, A.; Filipič, M. Genotoxicity and potential carcinogenicity of cyanobacterial toxins—A review. Mutat. Res. Rev. Mutat. Res. 2011, 727, 16–41. [Google Scholar] [CrossRef]
- El-Shehawy, R.; Gorokhova, E.; Fernández-Piñas, F.; del Campo, F.F. Global warming and hepatotoxin production by cyanobacteria: What can we learn from experiments? Water Res. 2012, 46, 1420–1429. [Google Scholar] [CrossRef]
- Paerl, H.W.; Gardner, W.S.; Havens, K.E.; Joyner, A.R.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Testai, E.; Scardala, S.; Vichi, S.; Buratti, F.M.; Funari, E. Risk to human health associated with the environmental occurrence of cyanobacterial neurotoxic alkaloids anatoxins and saxitoxins. Crit. Rev. Toxicol. 2016, 46, 385–419. [Google Scholar] [CrossRef]
- Reichwaldt, E.S.; Ghadouani, A. Effects of rainfall patterns on toxic cyanobacterial blooms in a changing climate: Between simplistic scenarios and complex dynamics. Water Res. 2012, 46, 1372–1393. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, C.; Liu, Y.; Jeppesen, E.; Svenning, J.C.; Wu, J.; Zhang, W.; Zhou, T.; Wang, P.; Nangombe, S.; et al. From unusual suspect to serial killer: Cyanotoxins boosted by climate change may jeopardize megafauna. Innovation 2021, 2, 100092. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Bloom dynamics of cyanobacteria and their toxins: Environmental health impacts and mitigation strategies. Front. Microbiol. 2015, 6, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurawell, R.W.; Chen, H.; Burke, J.M.; Prepas, E.E. Hepatotoxic cyanobacteria: A review of the biological importance of microcystins in freshwater environments. J. Toxicol. Environ. Health Part B Crit. Rev. 2005, 8, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2013, 15, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Praena, D.; Jos, Á.; Pichardo, S.; Moreno, I.M.; Cameán, A.M. Presence and bioaccumulation of microcystins and cylindrospermopsin in food and the effectiveness of some cooking techniques at decreasing their concentrations: A review. Food Chem. Toxicol. 2013, 53, 139–152. [Google Scholar] [CrossRef]
- Massey, I.Y.; Yang, F.; Ding, Z.; Yang, S.; Guo, J.; Tezi, C.; Al-Osman, M.; Kamegni, R.B.; Zeng, W. Exposure routes and health effects of microcystins on animals and humans: A mini-review. Toxicon 2018, 151, 156–162. [Google Scholar] [CrossRef]
- Malbrouck, C.; Kestemont, P. Effects of microcystins on fish. Environ. Toxicol. Chem. 2006, 25, 72–86. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.D.S.; Kozlowsky-Suzuki, B. Cyanotoxins: Bioaccumulation and effects on aquatic animals. Mar. Drugs 2011, 9, 2729–2772. [Google Scholar] [CrossRef]
- Nielsen, M.C.; Jiang, S.C. Can cyanotoxins penetrate human skin during water recreation to cause negative health effects? Harmful Algae 2020, 98, 101872. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Al Shehri, A.M. Microcystins in groundwater wells and their accumulation in vegetable plants irrigated with contaminated waters in Saudi Arabia. J. Hazard. Mater. 2009, 172, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.; Campos, A.; Vasconcelos, V.; Freitas, M. Effects of microcystin-LR and cylindrospermopsin on plant-soil systems: A review of their relevance for agricultural plant quality and public health. Environ. Res. 2017, 153, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbel, S.; Mougin, C.; Bouaïcha, N. Cyanobacterial toxins: Modes of actions, fate in aquatic and soil ecosystems, phytotoxicity and bioaccumulation in agricultural crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Romero-Oliva, C.S.; Contardo-Jara, V.; Block, T.; Pflugmacher, S. Accumulation of microcystin congeners in different aquatic plants and crops-A case study from lake Amatitlán, Guatemala. Ecotoxicol. Environ. Saf. 2014, 102, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt-Oliveira, M.d.C.; Cordeiro-Araújo, M.K.; Chia, M.A.; Arruda-Neto, J.D.d.T.; Oliveira, Ê.T.d.; dos Santos, F. Lettuce irrigated with contaminated water: Photosynthetic effects, antioxidative response and bioaccumulation of microcystin congeners. Ecotoxicol. Environ. Saf. 2016, 128, 83–90. [Google Scholar] [CrossRef]
- Zhang, Y.; Whalen, J.K.; Sauvé, S. Phytotoxicity and bioconcentration of microcystins in agricultural plants: Meta-analysis and risk assessment. Environ. Pollut. 2021, 272, 115966. [Google Scholar] [CrossRef]
- Testai, E.; Buratti, F.M.; Funari, E.; Manganelli, M.; Vichi, S.; Arnich, N.; Biré, R.; Fessard, V.; Sialehaamoa, A. Review and analysis of occurrence, exposure and toxicity of cyanobacteria toxins in food. EFSA Supporting Publ. 2017, 13, 1–309. [Google Scholar] [CrossRef]
- Codd, G.A.; Metcalf, J.S.; Beattie, K.A. Retention of Microcystis aeruginosa and microcystin by salad lettuce (Lactuca sativa) after spray irrigation with water containing cyanobacteria. Toxicon 1999, 37, 1181–1185. [Google Scholar] [CrossRef]
- Crush, J.R.; Briggs, L.R.; Sprosen, J.M.; Nichols, S.N. Effect of irrigation with lake water containing microcystins on microcystin content and growth of ryegrass, clover, rape, and lettuce. Environ. Toxicol. 2008, 23, 246–252. [Google Scholar] [CrossRef]
- Chen, J.; Han, F.X.; Wang, F.; Zhang, H.; Shi, Z. Accumulation and phytotoxicity of microcystin-LR in rice (Oryza sativa). Ecotoxicol. Environ. Saf. 2012, 76, 193–199. [Google Scholar] [CrossRef]
- Giannuzzi, L.; Sedan, D.; Echenique, R.; Andrinolo, D. An acute case of intoxication with cyanobacteria and cyanotoxins in recreational water in Salto Grande Dam, Argentina. Mar. Drugs 2011, 9, 2164–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, F.; Sedan, D.; D’Agostino, D.; Cavalieri, M.L.; Mullen, E.; Parot Varela, M.M.; Flores, C.; Caixach, J.; Andrinolo, D. Recreational exposure during algal bloom in carrasco beach, uruguay: A liver failure case report. Toxins 2017, 9, 267. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, J.S.; Richer, R.; Cox, P.A.; Codd, G.A. Cyanotoxins in desert environments may present a risk to human health. Sci. Total Environ. 2012, 421–422, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, S.M.F.O.; Carmichael, W.W.; Jochimsen, E.M.; Rinehart, K.L.; Lau, S.; Shaw, G.R.; Eaglesham, G.K. Human intoxication by microcystins during renal dialysis treatment in Caruaru-Brazil. Toxicology 2002, 181–182, 441–446. [Google Scholar] [CrossRef]
- Van Apeldoorn, M.E.; Van Egmond, H.P.; Speijers, G.J.A.; Bakker, G.J.I. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef] [PubMed]
- Wood, R. Acute animal and human poisonings from cyanotoxin exposure—A review of the literature. Environ. Int. 2016, 91, 276–282. [Google Scholar] [CrossRef]
- Kaebernick, M.; Neilan, B.A. Ecological and molecular investigations of cyanotoxin production. FEMS Microbiol. Ecol. 2006, 35, 1–9. [Google Scholar] [CrossRef]
- De Figueiredo, D.R.; Azeiteiro, U.M.; Esteves, S.M.; Gonçalves, F.J.M.; Pereira, M.J. Microcystin-producing blooms–A serious global public health issue. Ecotoxicol. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Aráoz, R.; Molgó, J.; Tandeau de Marsac, N. Neurotoxic cyanobacterial toxins. Toxicon 2010, 56, 813–828. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.P. Cyanobacterial Toxins as Allelochemicals with Potential Applications as Algaecides, Herbicides and Insecticides. Mar. Drugs 2008, 6, 117–146. [Google Scholar] [CrossRef]
- Demuez, M.; González-Fernández, C.; Ballesteros, M. Algicidal microorganisms and secreted algicides: New tools to induce microalgal cell disruption. Biotechnol. Adv. 2015, 33, 1615–1625. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sinha, R.P.; Incharoensakdi, A. The cyanotoxin-microcystins: Current overview. Rev. Environ. Sci. Biotechnol. 2014, 13, 215–249. [Google Scholar] [CrossRef]
- Díez-Quijada, L.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Occurrence and toxicity of microcystin congeners other than MC-LR and MC-RR: A review. Food Chem. Toxicol. 2019, 125, 106–132. [Google Scholar] [CrossRef] [PubMed]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural diversity, characterization and toxicology of microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef] [Green Version]
- Massey, I.Y.; Yang, F. A mini review on microcystins and bacterial degradation. Toxins 2020, 12, 268. [Google Scholar] [CrossRef] [Green Version]
- Díez-Quijada, L.; Puerto, M.; Gutiérrez-Praena, D.; Llana-Ruiz-Cabello, M.; Jos, A.; Cameán, A.M. Microcystin-RR: Occurrence, content in water and food and toxicological studies. A review. Environ. Res. 2019, 168, 467–489. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Wu, Q.; Liu, C.; Shen, J.Z.; Yan, W. Microcystin-LR exposure induced nephrotoxicity by triggering apoptosis in female zebrafish. Chemosphere 2019, 214, 598–605. [Google Scholar] [CrossRef]
- Hinojosa, M.G.; Gutiérrez-Praena, D.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Neurotoxicity induced by microcystins and cylindrospermopsin: A review. Sci. Total Environ. 2019, 668, 547–565. [Google Scholar] [CrossRef] [PubMed]
- Nasri, H.; El Herry, S.; Bouaïcha, N. First reported case of turtle deaths during a toxic Microcystis spp. bloom in Lake Oubeira, Algeria. Ecotoxicol. Environ. Saf. 2008, 71, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Z.; Ahmed, Z.; Bakr, A.; Hashem, M.; Alamri, S. Detection of free and bound microcystins in tilapia fish from Egyptian fishpond farms and its related public health risk assessment. J. Verbrauch. Lebensm. 2020, 15, 37–47. [Google Scholar] [CrossRef]
- Chen, J.; Xie, P. Tissue distributions and seasonal dynamics of the hepatotoxic microcystins-LR and -RR in two freshwater shrimps, Palaemon modestus and Macrobrachium nipponensis, from a large shallow, eutrophic lake of the subtropical China. Toxicon 2005, 45, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Z.A.; Carmichael, W.W.; Hussein, A.A. Estimation of microcystins in the freshwater fish Oreochromis niloticus in an Egyptian fish farm containing a Microcystis bloom. Environ. Toxicol. 2003, 18, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Poste, A.E.; Hecky, R.E.; Guildford, S.J. Evaluating microcystin exposure risk through fish consumption. Environ. Sci. Technol. 2011, 45, 5806–5811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas de Magalhães, V.; Moraes Soares, R.; Azevedo, S.M.F.O. Microcystin contamination in fish from the Jacarepaguá Lagoon (Rio de Janeiro, Brazil): Ecological implication and human health risk. Toxicon 2001, 39, 1077–1085. [Google Scholar] [CrossRef]
- Zamora-Barrios, C.A.; Nandini, S.; Sarma, S.S.S. Bioaccumulation of microcystins in seston, zooplankton and fish: A case study in Lake Zumpango, Mexico. Environ. Pollut. 2019, 249, 267–276. [Google Scholar] [CrossRef]
- Llana-Ruiz-Cabello, M.; Jos, A.; Cameán, A.; Oliveira, F.; Barreiro, A.; MacHado, J.; Azevedo, J.; Pinto, E.; Almeida, A.; Campos, A.; et al. Analysis of the use of cylindrospermopsin and/or microcystin-contaminatedwater in the growth, mineral content, and contamination of spinacia oleracea and lactuca sativa. Toxins 2019, 11, 624. [Google Scholar] [CrossRef] [Green Version]
- McElhiney, J.; Lawton, L.A.; Leifert, C. Investigations into the inhibitory effects of microcystins on plant growth, and the toxicity of plant tissues following exposure. Toxicon 2001, 39, 1411–1420. [Google Scholar] [CrossRef]
- Cordeiro-Araújo, M.K.; Chia, M.A.; Arruda-Neto, J.D.d.T.; Tornisielo, V.L.; Vilca, F.Z.; Bittencourt-Oliveira, M.d.C. Microcystin-LR bioaccumulation and depuration kinetics in lettuce and arugula: Human health risk assessment. Sci. Total Environ. 2016, 566–567, 1379–1386. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. IARC monographs on the evaluation of carcinogenic risks to humans. Ingested nitrate and nitrite, and cyanobacterial peptide toxins. Int. Agency Res. Cancer 2010, 94, 1–412. [Google Scholar]
- Zheng, C.; Zeng, H.; Lin, H.; Wang, J.; Feng, X.; Qiu, Z.; Chen, J.A.; Luo, J.; Luo, Y.; Huang, Y.; et al. Serum microcystin levels positively linked with risk of hepatocellular carcinoma: A case-control study in southwest China. Hepatology 2017, 66, 1519–1528. [Google Scholar] [CrossRef]
- Liu, W.; Wang, L.; Zheng, C.; Liu, L.; Wang, J.; Li, D.; Tan, Y.; Zhao, X.; He, L.; Shu, W. Microcystin-LR increases genotoxicity induced by aflatoxin B1 through oxidative stress and DNA base excision repair genes in human hepatic cell lines. Environ. Pollut. 2018, 233, 455–463. [Google Scholar] [CrossRef]
- Zhou, L.; Yu, H.; Chen, K. Relationship between microcystin in drinking water and colorectal cancer. Biomed. Environ. Sci. 2002, 15, 166–171. [Google Scholar] [PubMed]
- Pan, C.; Zhang, L.; Meng, X.; Qin, H.; Xiang, Z.; Gong, W.; Luo, W.; Li, D.; Han, X. Chronic exposure to microcystin-LR increases the risk of prostate cancer and induces malignant transformation of human prostate epithelial cells. Chemosphere 2021, 263, 128295. [Google Scholar] [CrossRef]
- Gehringer, M.M.; Adler, L.; Roberts, A.A.; Moffitt, M.C.; Mihali, T.K.; Mills, T.J.T.; Fieker, C.; Neilan, B.A. Nodularin, a cyanobacterial toxin, is synthesized in planta by symbiotic Nostoc sp. ISME J. 2012, 6, 1834–1847. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Shen, D.; Fang, D. Nodularins in poisoning. Clin. Chim. Acta 2013, 425, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Galanti, L.N.; Amé, M.V.; Wunderlin, D.A. Accumulation and detoxification dynamic of cyanotoxins in the freshwater shrimp Palaemonetes argentinus. Harmful Algae 2013, 27, 88–97. [Google Scholar] [CrossRef]
- Stewart, I.; Seawright, A.A.; Shaw, G.R. Cyanobacterial poisoning in livestock, wild mammals and birds—An overview. Adv. Exp. Med. 2008, 619, 613–637. [Google Scholar] [CrossRef]
- Krienitz, L.; Ballot, A.; Kotut, K.; Wiegand, C.; Pütz, S.; Metcalf, J.S.; Codd, G.A.; Stephan, P. Contribution of hot spring cyanobacteria to the mysterious deaths of Lesser Flamingos at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 2003, 43, 141–148. [Google Scholar] [CrossRef]
- Becker, V.; Ihara, P.; Yunes, J.S.; Huszar, V.L.M. Occurrence of anatoxin-a(s) during a bloom of Anabaena crassa in a water-supply reservoir in southern Brazil. J. Appl. Phycol. 2010, 22, 235–241. [Google Scholar] [CrossRef]
- Cook, W.O.; Beasley, V.R.; Dahlem, A.M.; Dellinger, J.A.; Harlin, K.S.; Carmichael, W.W. Comparison of effects of anatoxin-a(s) and paraoxon, physostigmine and pyridostigmine on mouse brain cholinesterase activity. Toxicon 1988, 26, 750–753. [Google Scholar] [CrossRef]
- Fiore, M.F.; de Lima, S.T.; Carmichael, W.W.; McKinnie, S.M.K.; Chekan, J.R.; Moore, B.S. Guanitoxin, re-naming a cyanobacterial organophosphate toxin. Harmful Algae 2020, 92, 101737. [Google Scholar] [CrossRef]
- Ohtani, I.; Moore, R.E.; Runnegar, M.T.C. Cylindrospermopsin: A Potent Hepatotoxin from the Blue-Green Alga Cylindrospermopsis raciborskii. J. Am. Chem. Soc. 1992, 114, 7941–7942. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B. In search of environmental role of cylindrospermopsin: A review on global distribution and ecology of its producers. Water Res. 2014, 66, 320–337. [Google Scholar] [CrossRef]
- Antunes, J.T.; Leão, P.N.; Vasconcelos, V.M. Cylindrospermopsis raciborskii: Review of the distribution, phylogeography, and ecophysiology of a global invasive species. Front. Microbiol. 2015, 6, 473. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yu, G.; Chen, Y.; Jia, N.; Li, R. Four decades of progress in cylindrospermopsin research: The ins and outs of a potent cyanotoxin. J. Hazard. Mater. 2021, 406, 124653. [Google Scholar] [CrossRef]
- Bouvy, M.; Falcão, D.; Marinho, M.; Pagano, M.; Moura, A. Occurrence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian tropical reservoirs during the 1998 drought. Aquat. Microb. Ecol. 2000, 23, 13–27. [Google Scholar] [CrossRef]
- Chellappa, N.T.; Mederios Costa, M.A. Dominant and co-existing species of Cyanobacteria from a Eutrophicated reservoir of Rio Grande do Norte State, Brazil. Acta Oecol. 2003, 24, S3–S10. [Google Scholar] [CrossRef]
- Briand, J.F.; Leboulanger, C.; Humbert, J.F.; Bernard, C.; Dufour, P. Cylindrospermopsis raciborskii (Cyanobacteria) invasion at mid-latitudes: Selection, wide physiological tolerance, or global warming? J. Phycol. 2004, 40, 231–238. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Bakr, A. Concentrations of cylindrospermopsin toxin in water and tilapia fish of tropical fishponds in Egypt, and assessing their potential risk to human health. Environ. Sci. Pollut. Res. 2018, 25, 36287–36297. [Google Scholar] [CrossRef] [PubMed]
- Costa, I.A.S.; Azevedo, S.M.F.O.; Senna, P.A.C.; Bernardo, R.R.; Costa, S.M.; Chellappa, N.T. Occurrence of toxin-producing cyanobacteria blooms in a Brazilian semiarid reservoir. Braz. J. Biol. 2006, 66, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Berry, J.P.; Jaja-Chimedza, A.; Dávalos-Lind, L.; Lind, O. Apparent bioaccumulation of cylindrospermopsin and paralytic shellfish toxins by finfish in lake catemaco (Veracruz, Mexico). Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2012, 29, 314–321. [Google Scholar] [CrossRef]
- De La Cruz, A.A.; Hiskia, A.; Kaloudis, T.; Chernoff, N.; Hill, D.; Antoniou, M.G.; He, X.; Loftin, K.; O’Shea, K.; Zhao, C.; et al. A review on cylindrospermopsin: The global occurrence, detection, toxicity and degradation of a potent cyanotoxin. Environ. Sci. Process. Impacts 2013, 15, 1979–2003. [Google Scholar] [CrossRef]
- Bittencourt-Oliveira, M.d.C.; Piccin-Santos, V.; Kujbida, P.; Moura, A.d.N. Cylindrospermopsin in Water Supply Reservoirs in Brazil Determined by Immunochemical and Molecular Methods. J. Water Resour. Prot. 2011, 3, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro-Araújo, M.K.; Chia, M.A.; Bittencourt-Oliveira, M.d.C. Potential human health risk assessment of cylindrospermopsin accumulation and depuration in lettuce and arugula. Harmful Algae 2017, 68, 217–223. [Google Scholar] [CrossRef]
- Hercog, K.; Štern, A.; Maisanaba, S.; Filipič, M.; Žegura, B. Plastics in cyanobacterial blooms—Genotoxic effects of binary mixtures of cylindrospermopsin and bisphenols in HepG2 cells. Toxins 2020, 12, 219. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, S.; Jiang, X. Cyanobacterial Toxins in Freshwater and Food: Important Sources of Exposure to Humans. Annu. Rev. Food Sci. Technol. 2017, 8, 281–304. [Google Scholar] [CrossRef] [PubMed]
- Pichardo, S.; Cameán, A.M.; Jos, A. In vitro toxicological assessment of cylindrospermopsin: A review. Toxins 2017, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, D.J.; Saker, M.L. The Palm Island mystery disease 20 years on: A review of research on the cyanotoxin cylindrospermopsin. Environ. Toxicol. 2003, 18, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusick, K.D.; Sayler, G.S. An overview on the marine neurotoxin, saxitoxin: Genetics, moleculartargets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [Green Version]
- Thottumkara, A.P.; Parsons, W.H.; Du Bois, J. Saxitoxin. Angew. Chem. Int. Ed. 2014, 53, 5760–5784. [Google Scholar] [CrossRef] [PubMed]
- Etheridge, S.M. Paralytic shellfish poisoning: Seafood safety and human health perspectives. Toxicon 2010, 56, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvão, J.A.; Oetterer, M.; Bittencourt-Oliveira, M.d.C.; Gouvêa-Barros, S.; Hiller, S.; Erler, K.; Luckas, B.; Pinto, E.; Kujbida, P. Saxitoxins accumulation by freshwater tilapia (Oreochromis niloticus) for human consumption. Toxicon 2009, 54, 891–894. [Google Scholar] [CrossRef]
- Berry, J.P.; Lind, O. First evidence of “paralytic shellfish toxins” and cylindrospermopsin in a Mexican freshwater system, Lago Catemaco, and apparent bioaccumulation of the toxins in “tegogolo” snails (Pomacea patula catemacensis). Toxicon 2010, 55, 930–938. [Google Scholar] [CrossRef]
- Clemente, Z.; Busato, R.H.; Oliveira Ribeiro, C.A.; Cestari, M.M.; Ramsdorf, W.A.; Magalhães, V.F.; Wosiack, A.C.; Silva de Assis, H.C. Analyses of paralytic shellfish toxins and biomarkers in a southern Brazilian reservoir. Toxicon 2010, 55, 396–406. [Google Scholar] [CrossRef]
- Durai, P.; Batool, M.; Choi, S. Structure and effects of cyanobacterial lipopolysaccharides. Mar. Drugs 2015, 13, 4217–4230. [Google Scholar] [CrossRef]
- Bláhová, L.; Adamovský, O.; Kubala, L.; Švihálková Šindlerová, L.; Zounková, R.; Bláha, L. The isolation and characterization of lipopolysaccharides from Microcystis aeruginosa, a prominent toxic water bloom forming cyanobacteria. Toxicon 2013, 76, 187–196. [Google Scholar] [CrossRef]
- Stewart, I.; Schluter, P.J.; Shaw, G.R. Cyanobacterial lipopolysaccharides and human health—A review. Environ. Health: Glob. Access Sci. Source 2006, 5, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.D.; Waack, J.; Lewis, A.; Edwards, C.; Lawton, L. Development and single-laboratory validation of a UHPLC-MS/MS method for quantitation of microcystins and nodularin in natural water, cyanobacteria, shellfish and algal supplement tablet powders. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1074–1075, 111–123. [Google Scholar] [CrossRef]
- Roy-Lachapelle, A.; Vo Duy, S.; Munoz, G.; Dinh, Q.T.; Bahl, E.; Simon, D.F.; Sauvé, S. Analysis of multiclass cyanotoxins (microcystins, anabaenopeptins, cylindrospermopsin and anatoxins) in lake waters using on-line SPE liquid chromatography high-resolution Orbitrap mass spectrometry. Anal. Methods 2019, 11, 5289–5300. [Google Scholar] [CrossRef] [Green Version]
- Tran, N.H.; Li, Y.; Reinhard, M.; Goh, K.C.; Sukarji, N.H.B.; You, L.; He, Y.; Gin, K.Y.H. Quantification of cylindrospermopsin, anatoxin-a and homoanatoxin-a in cyanobacterial bloom freshwater using direct injection/SPE coupled with UPLC-MS/MS. Sci. Total. Environ. 2020, 731, 139014. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Rautela, A.; Kesari, V.; Szlag, D.; Westrick, J.; Kumar, S. Recent developments in the methods of quantitative analysis of microcystins. J. Biochem. Mol. Toxicol. 2020, 34, e22582. [Google Scholar] [CrossRef]
- Xu, X.; Zhu, B.; Liu, Z.; Wang, F.; Liang, J. Simultaneous determination of eight microcystins in fish by PRiME pass-through cleanup and online solid phase extraction coupled to ultra high performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1125, 121709. [Google Scholar] [CrossRef]
- Manubolu, M.; Lee, J.; Riedl, K.M.; Kua, Z.X.; Collart, L.P.; Ludsin, S.A. Optimization of extraction methods for quantification of microcystin-LR and microcystin-RR in fish, vegetable, and soil matrices using UPLC–MS/MS. Harmful Algae 2018, 76, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.W.; Zhan, X.J.; Xiang, L.; Deng, Z.S.; Huang, B.H.; Wen, H.F.; Sun, T.F.; Cai, Q.Y.; Li, H.; Mo, C.H. Analysis of trace microcystins in vegetables using solid-phase extraction followed by high performance liquid chromatography triple-quadrupole mass spectrometry. J. Agric. Food Chem. 2014, 62, 11831–11839. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.Y.; Li, Z.G.; Ma, J.; Gong, T.-t.; Xian, Q.M. Analysis of trace microcystins in vegetables using matrix solid-phase dispersion followed by high performance liquid chromatography triple-quadrupole mass spectrometry detection. Talanta 2017, 173, 101–106. [Google Scholar] [CrossRef]
- Massey, I.Y.; Wu, P.; Wei, J.; Luo, J.; Ding, P.; Wei, H.; Yang, F. A mini-review on detection methods of microcystins. Toxins 2020, 12, 641. [Google Scholar] [CrossRef]
- Geis-Asteggiante, L.; Lehotay, S.J.; Fortis, L.L.; Paoli, G.; Wijey, C.; Heinzen, H. Development and validation of a rapid method for microcystins in fish and comparing LC-MS/MS results with ELISA. Anal. Bioanal. Chem. 2011, 401, 2617–2630. [Google Scholar] [CrossRef]
- Parker, C.H.; Stutts, W.L.; Degrasse, S.L. Development and Validation of a Liquid Chromatography-Tandem Mass Spectrometry Method for the Quantitation of Microcystins in Blue-Green Algal Dietary Supplements. J. Agric. Food Chem. 2015, 63, 10303–10312. [Google Scholar] [CrossRef]
- Díez-Quijada, L.; Guzmán-Guillén, R.; Prieto Ortega, A.I.; Llana-Ruíz-Cabello, M.; Campos, A.; Vasconcelos, V.; Jos, Á.; Cameán, A.M. New method for simultaneous determination of microcystins and cylindrospermopsin in vegetable matrices by SPE-UPLC-MS/MS. Toxins 2018, 10, 406. [Google Scholar] [CrossRef] [Green Version]
- Díez-Quijada Jiménez, L.; Guzmán-Guillén, R.; Cătunescu, G.M.; Campos, A.; Vasconcelos, V.; Jos, Á.; Cameán, A.M. A new method for the simultaneous determination of cyanotoxins (Microcystins and Cylindrospermopsin) in mussels using SPE-UPLC-MS/MS. Environ. Res. 2020, 185, 109284. [Google Scholar] [CrossRef]
- Haddad, S.P.; Bobbitt, J.M.; Taylor, R.B.; Lovin, L.M.; Conkle, J.L.; Chambliss, C.K.; Brooks, B.W. Determination of microcystins, nodularin, anatoxin-a, cylindrospermopsin, and saxitoxin in water and fish tissue using isotope dilution liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2019, 1599, 66–74. [Google Scholar] [CrossRef]
- Greer, B.; Maul, R.; Campbell, K.; Elliott, C.T. Detection of freshwater cyanotoxins and measurement of masked microcystins in tilapia from Southeast Asian aquaculture farms. Anal. Bioanal. Chem. 2017, 409, 4057–4069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romera-García, E.; Helmus, R.; Ballesteros-Gómez, A.; Visser, P.M. Multi-class determination of intracellular and extracellular cyanotoxins in freshwater samples by ultra-high performance liquid chromatography coupled to high resolution mass spectrometry. Chemosphere 2021, 274, 129770. [Google Scholar] [CrossRef]
- Di Pofi, G.; Favero, G.; Nigro Di Gregorio, F.; Ferretti, E.; Viaggiu, E.; Lucentini, L. Multi-residue Ultra Performance Liquid Chromatography-High resolution mass spectrometric method for the analysis of 21 cyanotoxins in surface water for human consumption. Talanta 2020, 211, 120738. [Google Scholar] [CrossRef] [PubMed]
- Roy-Lachapelle, A.; Solliec, M.; Bouchard, M.F.; Sauvé, S. Detection of cyanotoxins in algae dietary supplements. Toxins 2017, 9, 76. [Google Scholar] [CrossRef] [Green Version]
- Bogialli, S.; Bortolini, C.; Di Gangi, I.M.; Di Gregorio, F.N.; Lucentini, L.; Favaro, G.; Pastore, P. Liquid chromatography-high resolution mass spectrometric methods for the surveillance monitoring of cyanotoxins in freshwaters. Talanta 2017, 170, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Song, D.; Fang, S.; Liu, Y.; Zhou, X.; Long, F.; Zhu, A. Development of novel portable and reusable fiber optical chemiluminescent biosensor and its application for sensitive detection of microcystin-LR. Biosens. Bioelectron. 2018, 121, 27–33. [Google Scholar] [CrossRef]
- Vogiazi, V.; De La Cruz, A.; Mishra, S.; Shanov, V.; Heineman, W.R.; Dionysiou, D.D. A Comprehensive Review: Development of Electrochemical Biosensors for Detection of Cyanotoxins in Freshwater. ACS Sens. 2019, 4, 1151–1173. [Google Scholar] [CrossRef] [PubMed]
- Modley, L.A.S.; Rampedi, I.T.; Avenant-Oldewage, A.; Mhuka, V.; Nindi, M.; Van Dyk, C. Microcystin concentrations and liver histopathology in Clarias gariepinus and Oreochromis mossambicus from three impacted rivers flowing into a hyper-eutrophic freshwater system: A pilot study. Environ. Toxicol. Pharmacol. 2019, 71, 103222. [Google Scholar] [CrossRef]
- Nyakairu, G.W.A.; Nagawa, C.B.; Mbabazi, J. Assessment of cyanobacteria toxins in freshwater fish: A case study of Murchison Bay (Lake Victoria) and Lake Mburo, Uganda. Toxicon 2010, 55, 939–946. [Google Scholar] [CrossRef]
- Semyalo, R.; Rohrlack, T.; Naggawa, C.; Nyakairu, G.W. Microcystin concentrations in Nile tilapia (Oreochromis niloticus) caught from Murchison Bay, Lake Victoria and Lake Mburo: Uganda. Hydrobiologia 2010, 638, 235–244. [Google Scholar] [CrossRef]
- Amrani, A.; Nasri, H.; Azzouz, A.; Kadi, Y.; Bouaïcha, N. Variation in cyanobacterial hepatotoxin (microcystin) content of water samples and two species of fishes collected from a shallow lake in Algeria. Arch. Environ. Contam. Toxicol. 2014, 66, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Zewde, T.W.; Johansen, J.A.; Kifle, D.; Demissie, T.B.; Hansen, J.H.; Tadesse, Z. Concentrations of microcystins in the muscle and liver tissues of fish species from Koka reservoir, Ethiopia: A potential threat to public health. Toxicon 2018, 153, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Simiyu, B.M.; Oduor, S.O.; Rohrlack, T.; Sitoki, L.; Kurmayer, R. Microcystin content in phytoplankton and in small fish from eutrophic nyanza gulf, Lake Victoria, Kenya. Toxins 2018, 10, 275. [Google Scholar] [CrossRef] [Green Version]
- Nchabeleng, T.; Cheng, P.; Oberholster, P.J.; Botha, A.M.; Smit, W.J.; Luus-Powell, W.J. Microcystin-LR equivalent concentrations in fish tissue during a postbloom Microcystis exposure in Loskop Dam, South Africa. Afr. J. Aquat. Sci. 2014, 39, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Humpage, A.R.; Falconer, I.R. Oral toxicity of the cyanobacterial toxin cylindrospermopsin in male Swiss albino mice: Determination of no observed adverse effect level for deriving a drinking water guideline value. Environ. Toxicol. 2003, 18, 94–103. [Google Scholar] [CrossRef]
- Kotut, K.; Ballot, A.; Krienitz, L. Toxic cyanobacteria and their toxins in standing waters of Kenya: Implications for water resource use. J. Water Health 2006, 4, 233–245. [Google Scholar] [CrossRef]
- Zewde, T.W.; Kifle, D.; Johansen, J.A.; Demissie, T.B.; Hansen, J.H.; Tadesse, Z. Cyanobacterial abundance and microcystins in water, seston and fish tissues in Lake Hora-Arsedi (Ethiopia). Afr. J. Aquat. Sci. 2020, 45, 475–485. [Google Scholar] [CrossRef]
- Mchau, G.J. Cyanotoxins Occurrence in Drinking Waters and Risk of Exposure to Human in Ukerewe District Mwanza, Tanzania. Ph.D. Dissertation, The Nelson Mandela African Institution of Science and Technology, Arusha, Tanzania, 2020. [Google Scholar]
- Mchau, G.J.; Makule, E.; Machunda, R.; Gong, Y.Y.; Kimanya, M. Harmful algal bloom and associated health risks among users of Lake Victoria freshwater: Ukerewe Island, Tanzania. J. Water Health 2019, 17, 826–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Xie, P. Accumulation of hepatotoxic microcystes in freshwater mussels, aquatic insect larvae and oligochaetes in a large, shallow eutrophic lake (Lake Chaohu) of subtropical China. Fresenius Environ. Bull. 2008, 17, 849–854. [Google Scholar]
- Chen, J.; Xie, P.; Zhang, D.; Ke, Z.; Yang, H. In situ studies on the bioaccumulation of microcystins in the phytoplanktivorous silver carp (Hypophthalmichthys molitrix) stocked in Lake Taihu with dense toxic Microcystis blooms. Aquaculture 2006, 261, 1026–1038. [Google Scholar] [CrossRef]
- Chen, J.; Xie, P. Seasonal dynamics of the hepatotoxic microcystins in various organs of four freshwater bivalves from the large eutrophic Lake Taihu of subtropical China and the risk to human consumption. Environ. Toxicol. 2005, 20, 572–584. [Google Scholar] [CrossRef]
- Chen, J.; Xie, P.; Zhang, D.; Lei, H. In situ studies on the distribution patterns and dynamics of microcystins in a biomanipulation fish–bighead carp (Aristichthys nobilis). Environ. Pollut. 2007, 147, 150–157. [Google Scholar] [CrossRef]
- Peng, L.; Tang, Q.; Gu, J.; Lei, L.; Chen, W.; Song, L. Seasonal variation of microcystins and their accumulation in fish in two large shallow lakes of China. Ecotoxicology 2020, 29, 790–800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Xie, P.; Liu, Y.; Qiu, T. Transfer, distribution and bioaccumulation of microcystins in the aquatic food web in Lake Taihu, China, with potential risks to human health. Sci. Total Environ. 2009, 407, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Luo, W.; Lu, Y.; Giesy, J.P. Bioaccumulation of microcystins (MCs) in four fish species from Lake Taihu, China: Assessment of risks to humans. Sci. Total Environ. 2014, 487, 224–232. [Google Scholar] [CrossRef]
- Jiang, Y.; Xie, P.; Nie, Y. Concentration and bioaccumulation of cyanobacterial bioactive and odorous metabolites occurred in a large, shallow chinese lake. Bull. Environ. Contam. Toxicol. 2014, 93, 643–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Yang, Y.; Wu, Y.; Tao, J.; Cheng, B. Microcystin Bioaccumulation in Freshwater Fish at Different Trophic Levels from the Eutrophic Lake Chaohu, China. Bull. Environ. Contam. Toxicol. 2017, 99, 69–74. [Google Scholar] [CrossRef]
- Jing, M.; Lin, D.; Lin, J.; Li, Q.; Yan, H.; Feng, X. Mercury, microcystins and Omega-3 polyunsaturated fatty acids in farmed fish in eutrophic reservoir: Risk and benefit assessment. Environ. Pollut. 2021, 270, 116047. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Liu, Y.; Chen, W.; Liu, L.; Kent, M.; Song, L. Health risks associated with consumption of microcystin-contaminated fish and shellfish in three Chinese lakes: Significance for freshwater aquacultures. Ecotoxicol. Environ. Saf. 2010, 73, 1804–1811. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Deng, X.; Xie, P.; Chen, J.; Guo, L. Risk assessment of microcystins in silver carp (Hypophthalmichthys molitrix) from eight eutrophic lakes in China. Food Chem. 2013, 140, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Gurbuz, F.; Uzunmehmetoğlu, O.Y.; Diler, Ö.; Metcalf, J.S.; Codd, G.A. Occurrence of microcystins in water, bloom, sediment and fish from a public water supply. Sci. Total Environ. 2016, 562, 860–868. [Google Scholar] [CrossRef]
- Akcaalan, R.; Köker, L.; Oğuz, A.; Spoof, L.; Meriluoto, J.; Albay, M. First report of cylindrospermopsin production by two cyanobacteria (Dolichospermum mendotae and Chrysosporum ovalisporum) in Lake Iznik, Turkey. Toxins 2014, 6, 3173–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koker, L.; Akcaalan, R.; Oguz, A.; Gaygusuz, O.; Kose, C.A.; Gucver, S.; Karaaslan, Y.; Albay, M.; Kinaci, C. Distribution of toxic cyanobacteria and cyanotoxins in Turkish waterbodies. J. Environ. Prot. Ecol. 2017, 18, 425–432. [Google Scholar]
- Singh, S.; Asthana, R.K. Assessment of microcystin concentration in carp and catfish: A case study from Lakshmikund Pond, Varanasi, India. Bull. Environ. Contam. Toxicol. 2014, 92, 687–692. [Google Scholar] [CrossRef]
- Rezaitabar, S.; Esmaili Sari, A.; Bahramifar, N.; Ramezanpour, Z. Transfer, tissue distribution and bioaccumulation of microcystin LR in the phytoplanktivorous and carnivorous fish in Anzali wetland, with potential health risks to humans. Sci. Total Environ. 2017, 575, 1130–1138. [Google Scholar] [CrossRef]
- Ruangrit, K.; Peerapornpisal, Y.; Pekkoh, J.; Whangchai, N. Microcystin Accumulation in Nile Tilapia, Oreochromis niloticus and Giant Freshwater Prawns, Macrobrachium rosenbergii in Green Water System Cultivation. Int. J. Geosci. 2013, 4, 60–63. [Google Scholar] [CrossRef] [Green Version]
- Ruangrit, K.; Whangchai, N.; Pekkoh, J.; Ruangyuttikarn, W.; Peerapornpisal, Y. First report on microcystins contamination in giant freshwater prawn (Macrobrachium rosenbergii) and Nile tilapia (Tilapia nilotica) cultured in earthen ponds. Int. J. Agric. Biol. 2011, 13, 1025–1028. [Google Scholar]
- Pham, T.L.; Shimizu, K.; Dao, T.S.; Motoo, U. First report on free and covalently bound microcystins in fish and bivalves from Vietnam: Assessment of risks to humans. Environ. Toxicol. Chem. 2017, 36, 2953–2957. [Google Scholar] [CrossRef] [PubMed]
- Trung, B.; Dao, T.S.; Faassen, E.; Lürling, M. Cyanobacterial blooms and microcystins in Southern Vietnam. Toxins 2018, 10, 471. [Google Scholar] [CrossRef] [Green Version]
- Bi, X.; Dai, W.; Wang, X.; Dong, S.; Zhang, S.; Zhang, D.; Wu, M. Microcystins distribution, bioaccumulation, and Microcystis genotype succession in a fish culture pond. Sci. Total Environ. 2019, 688, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.j.; Zhang, J.-y.; Hong, Y.; Chen, Y. xu Evaluation of organ distribution of microcystins in the freshwater phytoplanktivorous fish Hypophthalmichthys molitrix. J. Zhejiang Univ. Sci. B 2007, 8, 116–120. [Google Scholar] [CrossRef] [Green Version]
- Ni, W.; Zhang, J.; Luo, Y. Microcystin accumulation in bighead carp (Aristichthys nobilis) during a Microcystis-dominated bloom and risk assessment of the dietary intake in a fish pond in China. Environ. Sci. Pollut. Res. 2017, 24, 8894–8902. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Z.; Song, Z.; Xie, Z.; Li, L.; Song, L. Bioaccumulation of microcystins in two freshwater gastropods from a cyanobacteria-bloom plateau lake, Lake Dianchi. Environ. Pollut. 2012, 164, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Xie, P.; Guo, L.; Li, L.; Miyabara, Y.; Park, H.D. Organ distribution and bioaccumulation of microcystins in freshwater fish at different trophic levels from the eutrophic Lake Chaohu, China. Environ. Toxicol. 2005, 20, 293–300. [Google Scholar] [CrossRef]
- Ruangsomboon, S.; Yongmanitchai, W.; Taveekijakarn, P.; Ganmanee, M. Cyanobacterial composition and microcystin accumulation in catfish pond. Chiang Mai J. Sci. 2014, 41, 27–38. [Google Scholar]
- Whangchai, N.; Wanno, S.; Gutierrez, R.; Kannika, K.; Promna, R.; Iwami, N.; Itayama, T. Accumulation of microcystins in water and economic fish in Phayao Lake, and fish ponds along the Ing River tributary in Chiang Rai, Thailand. Agric. Sci. 2013, 4, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Liu, W.; Zeng, H.; Pu, C.; Zhang, R.; Qiu, Z.; Chen, J.A.; Wang, L.; Tan, Y.; Zheng, C.; et al. Determination of Environmental Exposure to Microcystin and Aflatoxin as a Risk for Renal Function Based on 5493 Rural People in Southwest China. Environ. Sci. Technol. 2016, 50, 5346–5356. [Google Scholar] [CrossRef]
- Greer, B.; Meneely, J.P.; Elliott, C.T. Uptake and accumulation of Microcystin-LR based on exposure through drinking water: An animal model assessing the human health risk. Sci. Rep. 2018, 8, 4913. [Google Scholar] [CrossRef] [Green Version]
- Calado, S.L.d.M.; Santos, G.S.; Leite, T.P.B.; Wojciechowski, J.; Nadaline, M.; Bozza, D.C.; de Magalhães, V.F.; Cestari, M.M.; Prodocimo, V.; Silva de Assis, H.C. Depuration time and sublethal effects of microcystins in a freshwater fish from water supply reservoir. Chemosphere 2018, 210, 805–815. [Google Scholar] [CrossRef]
- Oliveira, N.B.; Schwartz, C.A.; Bloch, C.; Paulino, L.; Pires, O.R. Bioacumulation of cyanotoxins in hypophthalmichthys molitrix (silver carp) in paranoá lake, Brasilia-DF, Brazil. Bull. Environ. Contam. Toxicol. 2013, 90, 308–313. [Google Scholar] [CrossRef]
- Deblois, C.P.; Aranda-Rodriguez, R.; Giani, A.; Bird, D.F. Microcystin accumulation in liver and muscle of tilapia in two large Brazilian hydroelectric reservoirs. Toxicon 2008, 51, 435–448. [Google Scholar] [CrossRef]
- Chellappa, N.T.; Chellappa, S.L.; Chellappa, S. Harmful Phytoplankton Blooms and Fish Mortality in a eutrophicated reservoir of Northeast Brazil. Braz. Arch. Biol. Technol. 2008, 51, 833–841. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, V.F.; Marinho, M.M.; Domingos, P.; Oliveira, A.C.; Costa, S.M.; Azevedo, L.O.; Azevedo, S.M.F.O. Microcystins (cyanobacteria hepatotoxins) bioaccumulation in fish and crustaceans from Sepetiba Bay (Brasil, RJ). Toxicon 2003, 42, 289–295. [Google Scholar] [CrossRef]
- Berry, J.P.; Lee, E.; Walton, K.; Wilson, A.E.; Bernal-Brooks, F. Bioaccumulation of microcystins by fish associated with a persistent cyanobacterial bloom in Lago de Patzcuaro (Michoacan, Mexico). Environ. Toxicol. Chem. 2011, 30, 1621–1628. [Google Scholar] [CrossRef]
- Cazenave, J.; Wunderlin, D.A.; Bistoni, M.D.L.Á.; Amé, M.V.; Krause, E.; Pflugmacher, S.; Wiegand, C. Uptake, tissue distribution and accumulation of microcystin-RR in Corydoras paleatus, Jenynsia multidentata and Odontesthes bonariensis: A field and laboratory study. Aquat. Toxicol. 2005, 75, 178–190. [Google Scholar] [CrossRef]
- Amé, M.V.; Galanti, L.N.; Menone, M.L.; Gerpe, M.S.; Moreno, V.J.; Wunderlin, D.A. Microcystin-LR, -RR, -YR and -LA in water samples and fishes from a shallow lake in Argentina. Harmful Algae 2010, 9, 66–73. [Google Scholar] [CrossRef]
- De Bock, M.F.S.; Moraes, G.S.d.O.; Almeida, R.G.d.S.; Vieira, K.D.d.S.; Santoro, K.R.; Bicudo, Á.J.d.A.; Molica, R.J.R. Exposure of Nile Tilapia (Oreochromis niloticus) Fingerlings to a Saxitoxin-Producing Strain of Raphidiopsis (Cylindrospermopsis) raciborskii (Cyanobacterium) Reduces Growth Performance and Increases Mortality Rate. Environ. Toxicol. Chem. 2020, 39, 1409–1420. [Google Scholar] [CrossRef]
- Hauser-Davis, R.A.; Lavradas, R.T.; Lavandier, R.C.; Rojas, E.G.A.; Guarino, A.W.S.; Ziolli, R.L. Accumulation and toxic effects of microcystin in tilapia (Oreochromis niloticus) from an eutrophic Brazilian lagoon. Ecotoxicol. Environ. Saf. 2015, 112, 132–136. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for Drinking-Water Quality, 3rd ed.; WHO: Geneva, Switzerland, 2008; Volume 1, ISBN 9789241547611. [Google Scholar]
- WHO. Cyanobacterial toxins: Microcystin-LR in Drinking-water. In WHO Guidelines for Drinking-Water Quality; WHO: Geneva, Switzerland, 2003; pp. 95–110. [Google Scholar]
- World Health Organization. Cyanobacterial Toxins: Microcystins; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- World Health Organization. Cyanobacterial Toxins: Cylindrospermopsins; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- World Health Organization. Cyanobacterial Toxins: Saxitoxins; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- World Health Organization. Cyanobacterial Toxins: Anatoxin-a and Analogues; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- The European Parliament and the Council of the European Union Directive (EU). 2020/2184, EU (revised) Drinking Water Directive. Annex 1. Part B. Off. J. Eur. Communities 2020, 2019, 35. [Google Scholar]
- Aguilera, A.; Haakonsson, S.; Martin, M.V.; Salerno, G.L.; Echenique, R.O. Bloom-forming cyanobacteria and cyanotoxins in Argentina: A growing health and environmental concern. Limnologica 2018, 69, 103–114. [Google Scholar] [CrossRef]
- Copetti, D.; Finsterle, K.; Marziali, L.; Stefani, F.; Tartari, G.; Douglas, G.; Reitzel, K.; Spears, B.M.; Winfield, I.J.; Crosa, G.; et al. Eutrophication management in surface waters using lanthanum modified bentonite: A review. Water Res. 2015, 97, 162–174. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Wei, G.; Tan, X.; Li, L.; Li, M. Species-dependent variation in sensitivity of Microcystis species to copper sulfate: Implication in algal toxicity of copper and controls of blooms. Sci. Rep. 2017, 7, 40393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamyadi, A.; Greenstein, K.E.; Glover, C.M.; Adams, C.; Rosenfeldt, E.; Wert, E.C. Impact of hydrogen peroxide and copper sulfate on the delayed release of microcystin. Water 2020, 12, 1105. [Google Scholar] [CrossRef]
- Li, X.; Chen, S.; Zeng, J.; Chabi, K.; Song, W.; Xian, X.; Yu, X. Impact of chlorination on cell inactivation, toxin release and degradation of cyanobacteria of development and maintenance stage. Chem. Eng. J. 2020, 397, 125378. [Google Scholar] [CrossRef]
- Matthijs, H.C.P.; Visser, P.M.; Reeze, B.; Meeuse, J.; Slot, P.C.; Wijn, G.; Talens, R.; Huisman, J. Selective suppression of harmful cyanobacteria in an entire lake with hydrogen peroxide. Water Res. 2012, 46, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Roegner, A.F.; Brena, B.; González-Sapienza, G.; Puschner, B. Microcystins in potable surface waters: Toxic effects and removal strategies. J. Appl. Toxicol. 2014, 34, 441–457. [Google Scholar] [CrossRef]
- Abbas, T.; Kajjumba, G.W.; Ejjada, M.; Masrura, S.U.; Marti, E.J.; Khan, E.; Jones-lepp, T.L. Recent advancements in the removal of cyanotoxins from water using conventional and modified adsorbents—A contemporary review. Water 2020, 12, 2756. [Google Scholar] [CrossRef]
- Drogui, P.; Daghrir, R.; Simard, M.C.; Sauvageau, C.; Blais, J.F. Removal of microcystin-LR from spiked water using either activated carbon or anthracite as filter material. Environ. Technol. 2012, 33, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Nasri, H.; Bouaïcha, N.; Harche, M.K. A new morphospecies of Microcystis sp. forming bloom in the cheffia dam (Algeria): Seasonal variation of microcystin concentrations in raw water and their removal in a full-scale treatment plant. Environ. Toxicol. 2007, 22, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Park, J.A.; Jung, S.M.; Choi, J.W.; Kim, J.H.; Hong, S.; Lee, S.H. Mesoporous carbon for efficient removal of microcystin-LR in drinking water sources, Nak-Dong River, South Korea: Application to a field-scale drinking water treatment plant. Chemosphere 2018, 193, 883–891. [Google Scholar] [CrossRef]
- Shi, H.; Ding, J.; Timmons, T.; Adams, C. PH effects on the adsorption of saxitoxin by powdered activated carbon. Harmful Algae 2012, 19, 61–67. [Google Scholar] [CrossRef]
- Bajracharya, A.; Liu, Y.L.; Lenhart, J.J. The influence of natural organic matter on the adsorption of microcystin-LR by powdered activated carbon. Environ. Sci. Water Res. Technol. 2019, 5, 256–267. [Google Scholar] [CrossRef]
- Lian, L.; Cao, X.; Wu, Y.; Sun, D.; Lou, D. A green synthesis of magnetic bentonite material and its application for removal of microcystin-LR in water. Appl. Surf. Sci. 2014, 289, 245–251. [Google Scholar] [CrossRef]
- Lee, J.; Walker, H.W. Adsorption of microcystin-Lr onto iron oxide nanoparticles. Colloids Surf. A Physicochem. Eng. Asp. 2011, 373, 94–100. [Google Scholar] [CrossRef]
- He, X.; de la Cruz, A.A.; Hiskia, A.; Kaloudis, T.; O’Shea, K.; Dionysiou, D.D. Destruction of microcystins (cyanotoxins) by UV-254 nm-based direct photolysis and advanced oxidation processes (AOPs): Influence of variable amino acids on the degradation kinetics and reaction mechanisms. Water Res. 2015, 74, 227–238. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Z.; Zhou, N.; Shen, J.; Ye, M. Degradation and detoxification of microcystin-LR in drinking water by sequential use of UV and ozone. J. Environ. Sci. 2010, 22, 1897–1902. [Google Scholar] [CrossRef]
- Chang, J.; Chen, Z.L.; Wang, Z.; Kang, J.; Chen, Q.; Yuan, L.; Shen, J.M. Oxidation of microcystin-LR in water by ozone combined with UV radiation: The removal and degradation pathway. Chem. Eng. J. 2015, 276, 97–105. [Google Scholar] [CrossRef]
- Afzal, A.; Oppenländer, T.; Bolton, J.R.; El-Din, M.G. Anatoxin-a degradation by Advanced Oxidation Processes: Vacuum-UV at 172 nm, photolysis using medium pressure UV and UV/H2O2. Water Res. 2010, 44, 278–286. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Pelaez, M.; Westrick, J.A.; O’Shea, K.E.; Hiskia, A.; Triantis, T.; Kaloudis, T.; Stefan, M.I.; de la Cruz, A.A.; Dionysiou, D.D. Efficient removal of microcystin-LR by UV-C/H2O2 in synthetic and natural water samples. Water Res. 2012, 46, 1501–1510. [Google Scholar] [CrossRef]
- Chintalapati, P.; Mohseni, M. Degradation of cyanotoxin microcystin-LR in synthetic and natural waters by chemical-free UV/VUV radiation. J. Hazard. Mater. 2020, 381, 120921. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Hu, X.; Du, Y.; Jin, Q.; Liu, Y.; Tang, C.; Hu, X.; Li, G.; Chen, Z.; Zhou, D.; et al. Light-driven breakdown of microcystin-LR in water: A critical review. Chem. Eng. J. 2021, 417, 129244. [Google Scholar] [CrossRef]
- He, X.; Liu, Y.L.; Conklin, A.; Westrick, J.; Weavers, L.K.; Dionysiou, D.D.; Lenhart, J.J.; Mouser, P.J.; Szlag, D.; Walker, H.W. Toxic cyanobacteria and drinking water: Impacts, detection, and treatment. Harmful Algae 2016, 54, 174–193. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, H.; Liu, A.; Cao, Z.; Hao, J.; Gong, R. Identification of a new microcystin-degrading bacterium isolated from Lake Chaohu, China. Bull. Environ. Contam. Toxicol. 2015, 94, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhang, F.; Liu, C.; Wang, M. Biodegradation of microcystins by Bacillus sp. Strain EMB. Energy Procedia 2012, 16, 2054–2059. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, R.; Li, J. Current research scenario for microcystins biodegradation–A review on fundamental knowledge, application prospects and challenges. Sci. Total Environ. 2017, 595, 615–632. [Google Scholar] [CrossRef]
- Guzmán-Guillén, R.; Prieto, A.I.; Moreno, I.; Soria, M.E.; Cameán, A.M. Effects of thermal treatments during cooking, microwave oven and boiling, on the unconjugated microcystin concentration in muscle of fish (Oreochromis niloticus). Food Chem. Toxicol. 2011, 49, 2060–2067. [Google Scholar] [CrossRef]
- Guzmán-Guillén, R.; Maisanaba, S.; Prieto Ortega, A.I.; Valderrama-Fernández, R.; Jos, Á.; Cameán, A.M. Changes on cylindrospermopsin concentration and characterization of decomposition products in fish muscle (Oreochromis niloticus) by boiling and steaming. Food Control 2017, 77, 210–220. [Google Scholar] [CrossRef]
- Prieto, A.I.; Guzmán-Guillén, R.; Valderrama-Fernández, R.; Jos, Á.; Cameán, A.M. Influence of cooking (Microwaving and broiling) on cylindrospermopsin concentration in muscle of nile tilapia (Oreochromis niloticus) and characterization of decomposition products. Toxins 2017, 9, 177. [Google Scholar] [CrossRef]
- Meneely, J.P.; Elliott, C.T. Microcystins: Measuring human exposure and the impact on human health. Biomarkers 2013, 18, 639–649. [Google Scholar] [CrossRef]
- Mchau, G.J.; Machunda, R.; Kimanya, M.; Makule, E.; Gong, Y.Y.; Mpolya, E.; Meneely, J.P.; Elliott, C.T.; Greer, B. First Report of the Co-occurrence of Cylindrospermopsin, Nodularin and Microcystins in the Freshwaters of Lake Victoria, Tanzania. Expo. Health 2021, 13, 185–194. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.-a.; Zhao, Q.; Pu, C.; Qiu, Z.; Zhang, R.; Shu, W. A cross-sectional investigation of chronic exposure to microcystin in relationship to childhood liver damage in the three gorges reservoir region, China. Environ. Health Perspect. 2011, 119, 1483–1488. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, J.S.; Codd, G.A. Co-occurrence of cyanobacteria and cyanotoxins with other environmental health hazards: Impacts and implications. Toxins 2020, 12, 629. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ji, J.; Wang, J.-S.; Sun, X. Co-contamination and interaction of fungal toxins and other environmental toxins. Trends Food Sci. Technol. 2020, 103, 162–178. [Google Scholar] [CrossRef]
- Meneely, J.P.; Hajšlová, J.; Krska, R.; Elliott, C.T. Assessing the combined toxicity of the natural toxins, aflatoxin B1, fumonisin B1 and microcystin-LR by high content analysis. Food Chem. Toxicol. 2018, 121, 527–540. [Google Scholar] [CrossRef] [PubMed]



| Evaluation Parameters | Biological Methods | Chemical Methods | |||||
|---|---|---|---|---|---|---|---|
| Mouse Bioassay | ELISA Assay | PPI Assay | LC-UV, LC-PDA, LC-FLD | LC-MS/MS | LC-HRMS | Sensors | |
| Sensitive | low | ✓ | ✓ | ✓ | ✓✓ | ✓✓✓ | ✓✓ |
| Expensive | cheap | ✓ | ✓ | ✓ | ✓✓ | ✓✓✓ | cheap |
| Distinguished variants | ✓ | ✓✓ | ✓✓✓ | ||||
| Multiple classes of toxins | ✓ | ✓✓ | ✓✓ | ✓ | |||
| Quantitation | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |
| Retrospective analysis | ✓ | ||||||
| In situ analysis | ✓ | ||||||
| Trendy | ✓ | ✓ | ✓ | ||||
| Other | Ethical approval | Matrix interference | Not for all cyanotoxins | Derivatization for LC-FLD analysis | Highly trained analysts | Highly trained analysts | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdallah, M.F.; Van Hassel, W.H.R.; Andjelkovic, M.; Wilmotte, A.; Rajkovic, A. Cyanotoxins and Food Contamination in Developing Countries: Review of Their Types, Toxicity, Analysis, Occurrence and Mitigation Strategies. Toxins 2021, 13, 786. https://doi.org/10.3390/toxins13110786
Abdallah MF, Van Hassel WHR, Andjelkovic M, Wilmotte A, Rajkovic A. Cyanotoxins and Food Contamination in Developing Countries: Review of Their Types, Toxicity, Analysis, Occurrence and Mitigation Strategies. Toxins. 2021; 13(11):786. https://doi.org/10.3390/toxins13110786
Chicago/Turabian StyleAbdallah, Mohamed F., Wannes H. R. Van Hassel, Mirjana Andjelkovic, Annick Wilmotte, and Andreja Rajkovic. 2021. "Cyanotoxins and Food Contamination in Developing Countries: Review of Their Types, Toxicity, Analysis, Occurrence and Mitigation Strategies" Toxins 13, no. 11: 786. https://doi.org/10.3390/toxins13110786