Measurement over 1 Year of Neutralizing Antibodies in Cattle Immunized with Trivalent Vaccines Recombinant Alpha, Beta and Epsilon of Clostridium perfringens

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Safe Vaccine Formulations for Use in Cattle
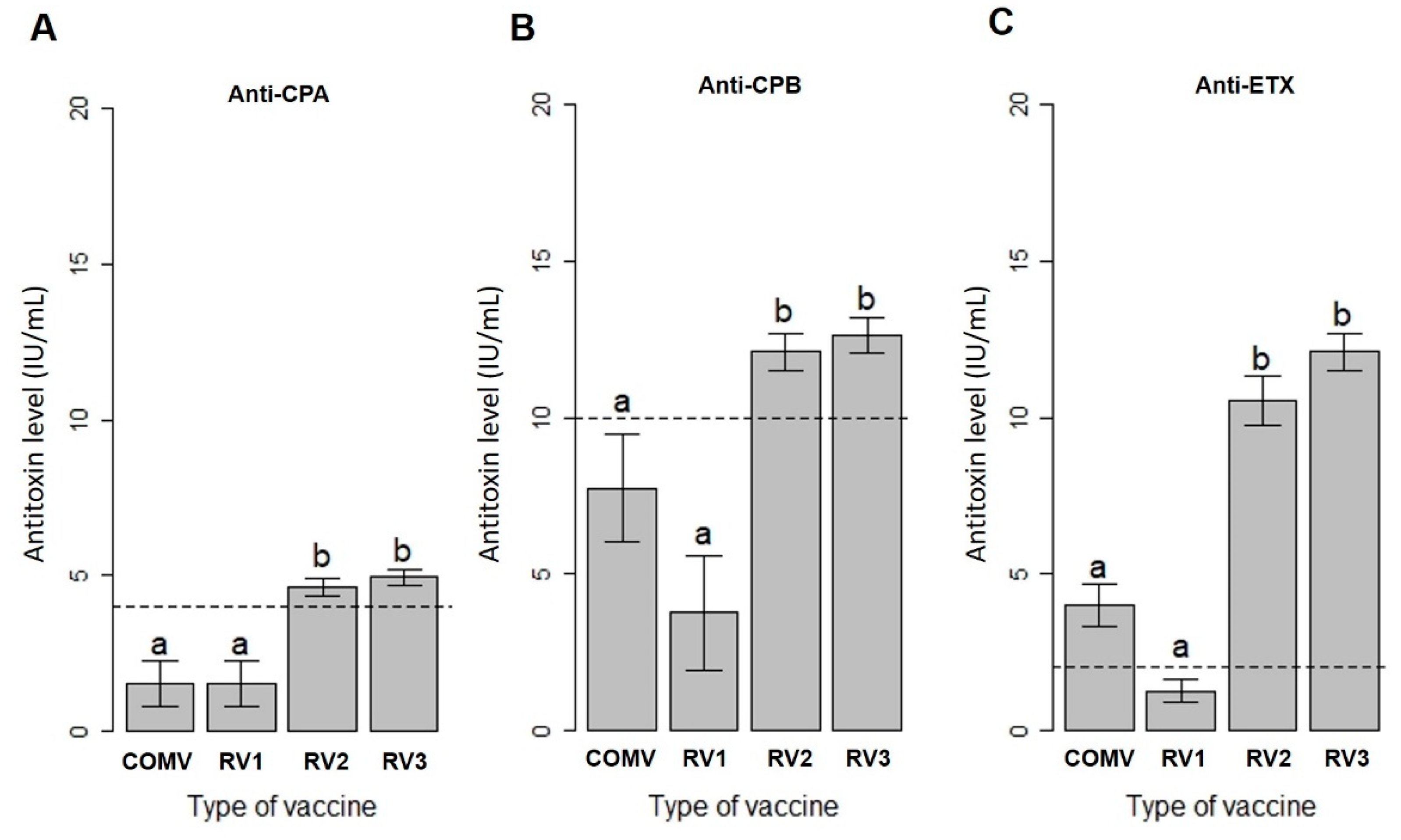
2.2. Antibody Vary According to the Vaccine Formulation
2.3. The Amount of Recombinant Protein Influences the Longevity of the Humoral Immune Response
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Ethics Declaration
5.2. Recombinant Vaccines
5.3. Sterility and Safety Test
5.4. Vaccination of Animals
5.5. Evaluation of the Humoral Immune Response
5.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- ABIEC—Associação Brasileira Das Indústrias Exportadoras de Carnes. Available online: http://abiec.com.br/ (accessed on 15 April 2020).
- Ferreira, M.; Moreira, G.; Cunha, C.; Mendonça, M.; Salvarani, F.; Moreira, Â.; Conceição, F. Recombinant Alpha, Beta, and Epsilon Toxins of Clostridium perfringens: Production Strategies and Applications as Veterinary Vaccines. Toxins 2016, 8, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, G.M.S.G.; Salvarani, F.M.; da Cunha, C.E.P.; Mendonça, M.; Moreira, Â.N.; Gonçalves, L.A.; Pires, P.S.; Lobato, F.C.F.; Conceição, F.R. Immunogenicity of a Trivalent Recombinant Vaccine Against Clostridium perfringens Alpha, Beta, and Epsilon Toxins in Farm Ruminants. Sci. Rep. 2016, 6, 22816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, R.O.S.; Duarte, M.C.; Oliveira Junior, C.A.; de Assis, R.A.; Lana, A.M.Q.; Lobato, F.C.F. Comparison of Humoral Neutralizing Antibody Response in Rabbits, Guinea Pigs, and Cattle Vaccinated with Epsilon and Beta Toxoids from Clostridium perfringens and C. Botulinum Types C and D Toxoids. Anaerobe 2018, 54, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Vidal, J.E.; McClane, B.A.; Gurjar, A.A. Clostridium perfringens Toxins Involved in Mammalian Veterinary Diseases. Open Toxinol. J. 2010, 3, 24–42. [Google Scholar] [CrossRef]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the Clostridium perfringens Toxin-Based Typing Scheme. Anaerobe 2018, 53, 5–10. [Google Scholar] [CrossRef]
- Abdel-Glil, M.Y.; Thomas, P.; Linde, J.; Busch, A.; Wieler, L.H.; Neubauer, H.; Seyboldt, C. Comparative in Silico Genome Analysis of Clostridium perfringens Unravels Stable Phylogroups with Different Genome Characteristics and Pathogenic Potential. Sci. Rep. 2021, 11, 6756. [Google Scholar] [CrossRef]
- Mehdizadeh Gohari, I.; Navarro, M.A.; Li, J.; Shrestha, A.; Uzal, F.; McClane, B.A. Pathogenicity and Virulence of Clostridium perfringens. Virulence 2021, 12, 723–753. [Google Scholar] [CrossRef]
- Geier, R.R.; Rehberger, T.G.; Smith, A.H. Comparative Genomics of Clostridium perfringens Reveals Patterns of Host-Associated Phylogenetic Clades and Virulence Factors. Front. Microbiol. 2021, 12, 649953. [Google Scholar] [CrossRef]
- Lobato, F.C.F.; Lima, C.G.R.D.; Assis, R.A.; Pires, P.S.; Silva, R.O.S.; Salvarani, F.M.; Carmo, A.O.; Contigli, C.; Kalapothakis, E. Potency against Enterotoxemia of a Recombinant Clostridium perfringens Type D Epsilon Toxoid in Ruminants. Vaccine 2010, 28, 6125–6127. [Google Scholar] [CrossRef]
- Ferreira, M.R.A.; dos Santos, F.D.; da Cunha, C.E.P.; Moreira, C.; Donassolo, R.A.; Magalhães, C.G.; Belo Reis, A.S.; Oliveira, C.M.C.; Barbosa, J.D.; Leite, F.P.L.; et al. Immunogenicity of Clostridium perfringens Epsilon Toxin Recombinant Bacterin in Rabbit and Ruminants. Vaccine 2018, 36, 7589–7592. [Google Scholar] [CrossRef]
- Moreira, C.; Ferreira, M.; da Cunha, C.; Donassolo, R.; Finger, P.; Moreira, G.; Otaka, D.; de Sousa, L.; Barbosa, J.; Moreira, Â.; et al. Immunogenicity of a Bivalent Non-Purified Recombinant Vaccine against Botulism in Cattle. Toxins 2018, 10, 381. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, L.A.; Lobato, Z.I.P.; Silva, R.O.S.; Salvarani, F.M.; Pires, P.S.; Assis, R.A.; Lobato, F.C.F. Selection of a Clostridium perfringens Type D Epsilon Toxin Producer via Dot-Blot Test. Arch. Microbiol. 2009, 191, 847–851. [Google Scholar] [CrossRef]
- Titball, R.W. Clostridium perfringens Vaccines. Vaccine 2009, 27, D44–D47. [Google Scholar] [CrossRef]
- Salvarani, F.M.; Conceição, F.R.; Cunha, C.E.P.; Moreira, G.M.S.G.; Pires, P.S.; Silva, R.O.S.; Alves, G.G.; Lobato, F.C.F. Vaccination with Recombinant Clostridium perfringens Toxoids α and β Promotes Elevated Antepartum and Passive Humoral Immunity in Swine. Vaccine 2013, 31, 4152–4155. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.R.A.; Motta, J.F.; Azevedo, M.L.; dos Santos, L.M.; Júnior, C.M.; Rodrigues, R.R.; Donassolo, R.A.; dos Reis, A.S.B.; Barbosa, J.D.; Salvarani, F.M.; et al. Inactivated Recombinant Escherichia coli as a Candidate Vaccine against Clostridium perfringens Alpha Toxin in Sheep. Anaerobe 2019, 59, 163–166. [Google Scholar] [CrossRef]
- Otaka, D.; Barbosa, J.; Moreira, C.; Ferreira, M.; Cunha, C.; Brito, A.; Donassolo, R.; Moreira, Â.; Conceição, F.; Salvarani, F. Humoral Response of Buffaloes to a Recombinant Vaccine against Botulism Serotypes C and D. Toxins 2017, 9, 297. [Google Scholar] [CrossRef] [Green Version]
- Otaka, D.Y.; Barbosa, J.D.; de Souza, L.A.; Moreira, C.; Ferreira, M.R.A.; Donassolo, R.A.; Conceição, F.R.; Salvarani, F.M. Recombinant Vaccine against Botulism in Buffaloes: Evaluation of the Humoral Immune Response over 12 Months. Anaerobe 2020, 63, 102201. [Google Scholar] [CrossRef]
- Freitas, N.F.Q.R.; Barbosa, J.D.; Otaka, D.Y.; Ferreira, M.R.A.; Rodrigues, R.R.; Moreira, C., Jr.; Conceição, F.R.; Salvarani, F.M. Clostridium perfringens α and β Recombinant Toxoids in Equine Immunization. Pesq. Vet. Bras. 2020, 40, 776–780. [Google Scholar] [CrossRef]
- Moreira, C.; Ferreira, M.R.A.; Finger, P.F.; Magalhães, C.G.; Cunha, C.E.P.; Rodrigues, R.R.; Otaka, D.Y.; Galvão, C.C.; Salvarani, F.M.; Moreira, Â.N.; et al. Protective Efficacy of Recombinant Bacterin Vaccine against Botulism in Cattle. Vaccine 2020, 38, 2519–2526. [Google Scholar] [CrossRef]
- Jiang, Z.; De, Y.; Chang, J.; Wang, F.; Yu, L. Induction of Potential Protective Immunity against Enterotoxemia in Calves by Single or Multiple Recombinant Clostridium perfringens Toxoids: Immunogenicity of Recombinant Toxoids. Microbiol. Immunol. 2014, 58, 621–627. [Google Scholar] [CrossRef]
- Augusto de Oliveira, C.; Duarte, M.C.; Antunes de Assis, R.; Alves, G.G.; Silva, R.O.S.; Faria Lobato, F.C. Humoral Responses in Cattle to Commercial Vaccines Containing Clostridium perfringens Epsilon Toxoid and C. botulinum Types C and D Toxoids Last Less than a-Year. Anaerobe 2019, 59, 72–75. [Google Scholar] [CrossRef]
- Ministério Da Agricultura Pecuária e Abastecimento Do Brasil (MAPA). Portaria n 49;1997. Brasília: Diário Oficial Da União. Available online: http://sistemasweb.agricultura.gov.br/sislegis/action/detalhaAto.do?method=visualizarAtoPortalMapa&chave=1924117692 (accessed on 25 May 2020).
- Rossi, A.; Mónaco, A.; Guarnaschelli, J.; Silveira, F.; Iriarte, A.; Benecke, A.G.; Chabalgoity, J.A. Temporal Evolution of Anti- Clostridium Antibody Responses in Sheep after Vaccination with Polyvalent Clostridial Vaccines. Vet. Immunol. Immunopathol. 2018, 202, 46–51. [Google Scholar] [CrossRef]
- Schwartzkoff, C.; Egerton, J.; Stewart, D.; Lehrbach, P.; Elleman, T.; Hoyne, P. The Effects of Antigenic Competition on the Efficacy of Multivalent Footrot Vaccines. Aust. Veter. J. 1993, 70, 123–126. [Google Scholar] [CrossRef]
- Hunt, J.D.; Jackson, D.C.; Brown, L.E.; Wood, P.R.; Stewart, D.J. Antigenic Competition in a Multivalent Foot Rot Vaccine. Vaccine 1994, 12, 457–464. [Google Scholar] [CrossRef]
- Simpson, K.M.; Callan, R.J.; Van Metre, D.C. Clostridial Abomasitis and Enteritis in Ruminants. Vet. Clin. N. Am. Food Anim. Pract. 2018, 34, 155–184. [Google Scholar] [CrossRef]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.-H.; Lee, K.W.; Lillehoj, E.P.; Hong, Y.H.; An, D.-J.; Jeong, W.; Chun, J.-E.; Bertrand, F.; et al. Vaccination with Clostridium perfringens Recombinant Proteins in Combination with MontanideTM ISA 71 VG Adjuvant Increases Protection against Experimental Necrotic Enteritis in Commercial Broiler Chickens. Vaccine 2012, 30, 5401–5406. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Alves Ferreira, M.R.; Donassolo, R.A.; Ferreira Alves, M.L.; Motta, J.F.; Junior, C.M.; Salvarani, F.M.; Moreira, A.N.; Conceicao, F.R. Evaluation of the Expression and Immunogenicity of Four Versions of Recombinant Clostridium perfringens Beta Toxin Designed by Bioinformatics Tools. Anaerobe 2021, 69, 102326. [Google Scholar] [CrossRef]
- Roos, T.B.; de Moraes, C.M.; Sturbelle, R.T.; Dummer, L.A.; Fischer, G.; Leite, F.P.L. Probiotics Bacillus Toyonensis and Saccharomyces Boulardii Improve the Vaccine Immune Response to Bovine Herpesvirus Type 5 in Sheep. Res. Vet. Sci. 2018, 117, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Roos, T.B.; Tabeleão, V.C.; Dümmer, L.A.; Schwegler, E.; Goulart, M.A.; Moura, S.V.; Corrêa, M.N.; Leite, F.P.L.; Gil-Turnes, C. Effect of Bacillus cereus Var. Toyoi and Saccharomyces boulardii on the Immune Response of Sheep to Vaccines. Food Agric. Immunol. 2010, 21, 113–118. [Google Scholar] [CrossRef]
- Santos, F.D.S.; Mazzoli, A.; Maia, A.R.; Saggese, A.; Isticato, R.; Leite, F.; Iossa, S.; Ricca, E.; Baccigalupi, L. A Probiotic Treatment Increases the Immune Response Induced by the Nasal Delivery of Spore-Adsorbed TTFC. Microb. Cell Fact. 2020, 19, 42. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.D.S.; Ferreira, M.R.A.; Maubrigades, L.R.; Gonçalves, V.S.; Lara, A.P.S.; Moreira, C.; Salvarani, F.M.; Conceição, F.R.; Leivas Leite, F.P. Bacillus toyonensis BCT-7112 T Transient Supplementation Improves Vaccine Efficacy in Ewes Vaccinated against Clostridium perfringens Epsilon Toxin. J. Appl. Microbiol. 2021, 130, 699–706. [Google Scholar] [CrossRef]
- Souza, A.M.; Reis, J.K.P.; Assis, R.A.; Horta, C.C.; Siqueira, F.F.; Facchin, S.; Alvarenga, E.R.; Castro, C.S.; Salvarani, F.M.; Silva, R.O.S.; et al. Molecular Cloning and Expression of Epsilon Toxin from Clostridium perfringens Type D and Tests of Animal Immunization. Genet. Mol. Res. 2010, 9, 266–276. [Google Scholar] [CrossRef]
- United States Department of Agriculture (USDA). Conditional Licenses for Products Containing Clostridium Perfringens Type A; Center for Veterinary Biologics Notice No. 02-25; USDA: Ames, IA, USA, 2002. [Google Scholar]
- COE (Council of Europe). European Pharmacopoeia, 3rd ed.; Maisonneuve S.A.: Sainte-Ruffine, France, 1998; p. 561. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints l’2. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Toxinotype | Toxins | Diseases (Affected Animals) |
|---|---|---|
| A | CPA | Gas gangrene (all ruminants) and enterotoxemia (ovine) |
| CPA, CPE | Enteritis (caprine) | |
| CPA, CPB2 | Abomasitis (calves) | |
| B | CPA, CPB, ETX | Necrotic enteritis and hemorrhagic enterotoxemia (bovine and ovine) |
| C | CPA, CPB | Necrotic enteritis and enterotoxemia (bovine, ovine and caprine) |
| D | CPA, ETX | Enterotoxemia (ovine, bovine, and caprine) |
| E | CPA, ITX | Hemorrhagic enteritis (lambs, and calves) |
| a Seroconversion Rate | Anti-CPA (56 Days) | Anti-CPB (56 Days) | Anti-ETX (56 Days) | |
|---|---|---|---|---|
| Vaccines | ||||
| COMV | 37.5% | 75% | 100% | |
| RV1 | 37.5% | 37.5% | 62.5% | |
| RV2 | 100% | 100% | 100% | |
| RV3 | 100% | 100% | 100% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galvão, C.C.; Barbosa, J.D.; Oliveira, C.M.C.; Otaka, D.Y.; Silva, P.R.O.; Ferreira, M.R.A.; Moreira Júnior, C.; Conceição, F.R.; Salvarani, F.M. Measurement over 1 Year of Neutralizing Antibodies in Cattle Immunized with Trivalent Vaccines Recombinant Alpha, Beta and Epsilon of Clostridium perfringens. Toxins 2021, 13, 594. https://doi.org/10.3390/toxins13090594
Galvão CC, Barbosa JD, Oliveira CMC, Otaka DY, Silva PRO, Ferreira MRA, Moreira Júnior C, Conceição FR, Salvarani FM. Measurement over 1 Year of Neutralizing Antibodies in Cattle Immunized with Trivalent Vaccines Recombinant Alpha, Beta and Epsilon of Clostridium perfringens. Toxins. 2021; 13(9):594. https://doi.org/10.3390/toxins13090594
Chicago/Turabian StyleGalvão, Cleideanny C., José D. Barbosa, Carlos M. C. Oliveira, Denis Y. Otaka, Paulo R. O. Silva, Marcos R. A. Ferreira, Clóvis Moreira Júnior, Fabricio R. Conceição, and Felipe M. Salvarani. 2021. "Measurement over 1 Year of Neutralizing Antibodies in Cattle Immunized with Trivalent Vaccines Recombinant Alpha, Beta and Epsilon of Clostridium perfringens" Toxins 13, no. 9: 594. https://doi.org/10.3390/toxins13090594
APA StyleGalvão, C. C., Barbosa, J. D., Oliveira, C. M. C., Otaka, D. Y., Silva, P. R. O., Ferreira, M. R. A., Moreira Júnior, C., Conceição, F. R., & Salvarani, F. M. (2021). Measurement over 1 Year of Neutralizing Antibodies in Cattle Immunized with Trivalent Vaccines Recombinant Alpha, Beta and Epsilon of Clostridium perfringens. Toxins, 13(9), 594. https://doi.org/10.3390/toxins13090594