Botulinum Neurotoxin Type A Directly Affects Sebocytes and Modulates Oleic Acid-Induced Lipogenesis

Abstract
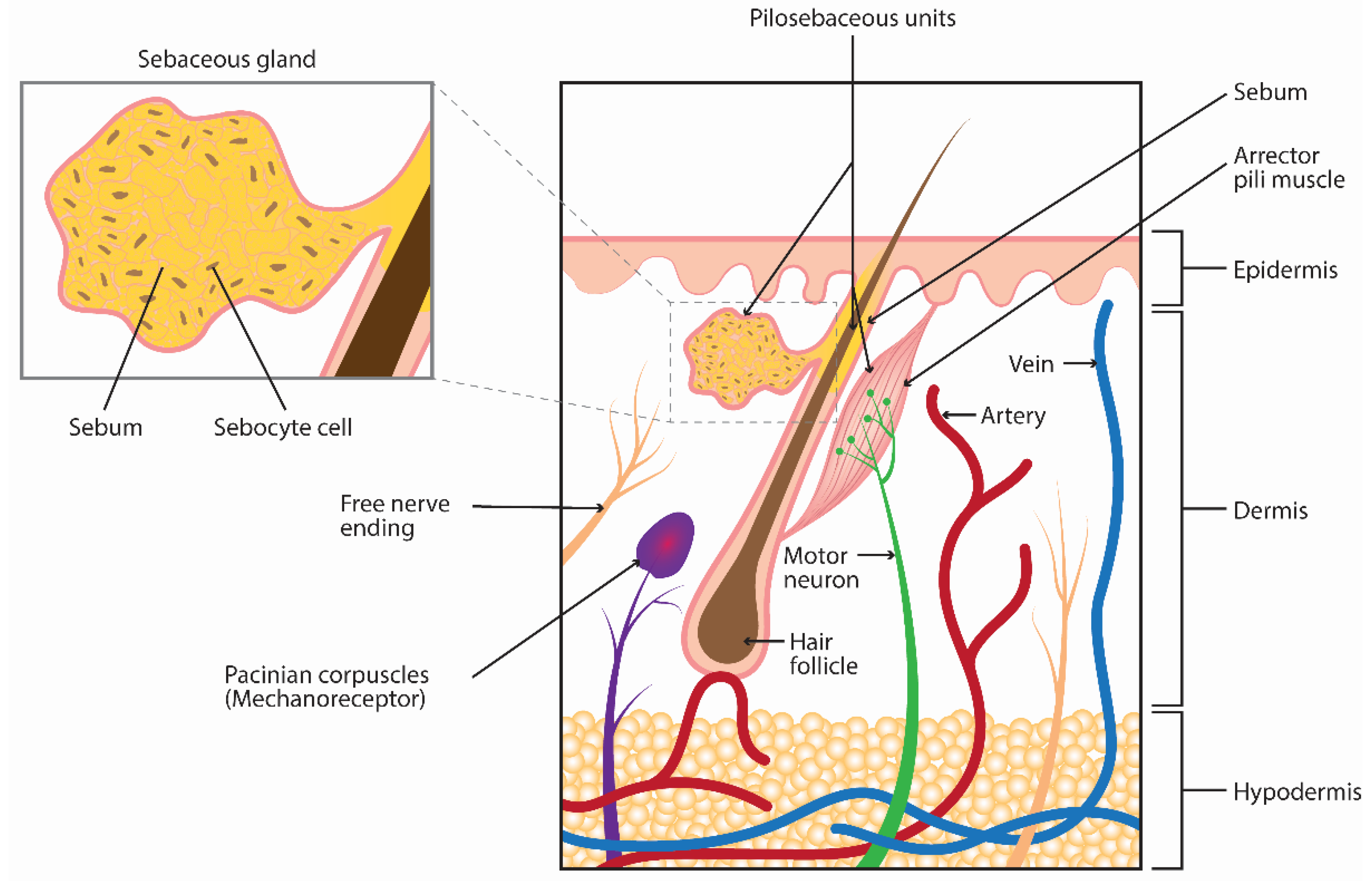
:1. Introduction
2. Results
2.1. Oleic Acid-Induced Lipogenesis in Cultured SEB-1 Cells as an In Vitro Model of Oily Skin
2.2. Effect of onabotA and rHC/A on OA-Induced Lipogenesis in SEB-1 Cells
2.3. Dose-Dependent Effects of BoNT/A on Lipogenesis in SEB-1 Cells
2.4. FGFRs Are Involved in BoNT/A-Mediated Inhibition of OA-Induced Lipogenesis
2.5. Effect of SV2 and Ganglioside Mutants of rHC/A on SEB-1 Lipogenesis
2.6. Effect of BoNT/A on OA-Treated Human Primary Sebocytes
3. Discussion
4. Materials and Methods
4.1. Primary Cells and Cell Lines
4.2. Drugs and Treatments
4.3. Oil Red O Staining and Bright Field Imaging
4.4. Lipid Droplets Fluorescence Assay
4.5. RNA Sequencing
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BoNT/A | botulinum neurotoxin type A |
| OnabotA | onabotulinumtoxinA |
| α-MSH | alpha melanocyte-stimulating hormone |
| EGF | epidermal growth factor |
| FGF | fibroblast growth factor |
| FGFR | fibroblast growth factor receptors |
| IGF-1 | insulin-like growth factor 1 |
| IGF1R | insulin-like growth factor 1 receptor |
| ACh | acetylcholine |
| LC | light chain of BoNT/A |
| HC | heavy chain of BoNT/A |
| HC | receptor binding domain |
| rHC/A | recombinant receptor binding domain of BoNT/A |
| HN | translocation domain |
| NTNH | nontoxic nonhemagglutinin protein |
| rNTNH-HC/A | recombinant binding domain homolog of nontoxic nonhemagglutinin protein |
| HAs | hemagglutinins |
| SNAP-25 | synaptosomal-associated protein-25 kDa |
| DHT | dihydrotestosterone |
| CaCl2 | calcium chloride |
| PPAR | peroxisome proliferator-activated receptor |
| TGF-α | transforming growth factor alpha |
| OA | oleic acid |
| EC50 | half maximal effective concentration |
| SV2 | synaptic vesicle protein 2 |
| AR | androgen receptor |
References
- Sakuma, T.H.; Maibach, H.I. Oily skin: An overview. Skin Pharmacol. Physiol. 2012, 25, 227–235. [Google Scholar] [CrossRef]
- Endly, D.C.; Miller, R.A. Oily Skin: A review of Treatment Options. J. Clin. Aesthet. Dermatol. 2017, 10, 49–55. [Google Scholar] [PubMed]
- Arbuckle, R.; Atkinson, M.J.; Clark, M.; Abetz, L.; Lohs, J.; Kuhagen, I.; Harness, J.; Draelos, Z.; Thiboutot, D.; Blume-Peytavi, U.; et al. Patient experiences with oily skin: The qualitative development of content for two new patient reported outcome questionnaires. Health Qual. Life Outcomes 2008, 6, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, M.E.; Downing, D.T. Chemistry and function of mammalian sebaceous lipids. Adv. Lipid Res. 1991, 24, 263–301. [Google Scholar] [CrossRef] [PubMed]
- Thiboutot, D. Regulation of human sebaceous glands. J. Investig. Dermatol. 2004, 123, 1–12. [Google Scholar] [CrossRef]
- Picardo, M.; Ottaviani, M.; Camera, E.; Mastrofrancesco, A. Sebaceous gland lipids. Dermato-Endocrinology 2009, 1, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.R.; Thiboutot, D.M. Thematic review series: Skin lipids. Sebaceous gland lipids: Friend or foe? J. Lipid Res. 2008, 49, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Zouboulis, C.C.; Baron, J.M.; Böhm, M.; Kippenberger, S.; Kurzen, H.; Reichrath, J.; Thielitz, A. Frontiers in sebaceous gland biology and pathology. Exp. Dermatol. 2008, 17, 542–551. [Google Scholar] [CrossRef]
- Fischer, H.; Fumicz, J.; Rossiter, H.; Napirei, M.; Buchberger, M.; Tschachler, E.; Eckhart, L. Holocrine Secretion of Sebum Is a Unique DNase2-Dependent Mode of Programmed Cell Death. J. Investig. Dermatol. 2017, 137, 587–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemann, C.; Horsley, V. Development and homeostasis of the sebaceous gland. Semin. Cell Dev. Biol. 2012, 23, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C. Sebaceous gland receptors. Dermato-Endocrinology 2009, 1, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Zouboulis, C.C. The human skin as a hormone target and an endocrine gland. Hormones (Athens) 2004, 3, 9–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whang, S.W.; Lee, S.E.; Kim, J.M.; Kim, H.J.; Jeong, S.K.; Zouboulis, C.C.; Seo, J.T.; Lee, S.H. Effects of α-melanocyte-stimulating hormone on calcium concentration in SZ95 sebocytes. Exp. Dermatol. 2011, 20, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Barrault, C.; Garnier, J.; Pedretti, N.; Cordier-Dirikoc, S.; Ratineau, E.; Deguercy, A.; Bernard, F.-X. Androgens induce sebaceous differentiation in sebocyte cells expressing a stable functional androgen receptor. J. Steroid Biochem. Mol. Biol. 2015, 152, 34–44. [Google Scholar] [CrossRef]
- Makrantonaki, E.; Vogel, K.; Fimmel, S.; Oeff, M.; Seltmann, H.; Zouboulis, C.C. Interplay of IGF-I and 17beta-estradiol at age-specific levels in human sebocytes and fibroblasts in vitro. Exp. Gerontol. 2008, 43, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Makrantonaki, E.; Adjaye, J.; Herwig, R.; Brink, T.C.; Groth, D.; Hultschig, C.; Lehrach, H.; Zouboulis, C.C. Age-specific hormonal decline is accompanied by transcriptional changes in human sebocytes in vitro. Aging Cell 2006, 5, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Dahlhoff, M.; Camera, E.; Picardo, M.; Zouboulis, C.C.; Chan, L.; Chang, B.H.-J.; Schneider, M.R. PLIN2, the major perilipin regulated during sebocyte differentiation, controls sebaceous lipid accumulation in vitro and sebaceous gland size in vivo. Biochim. Biophys. Acta 2013, 1830, 4642–4649. [Google Scholar] [CrossRef] [Green Version]
- Zouboulis, C.C. Epidermal growth factor receptor and the sebaceous gland. Exp. Dermatol. 2013, 22, 695–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akimoto, N.; Sato, T.; Sakiguchi, T.; Kitamura, K.; Kohno, Y.; Ito, A. Cell proliferation and lipid formation in hamster sebaceous gland cells. Dermatology 2002, 204, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.J.; Park, S.B.; Sohn, K.C.; Lee, Y.; Seo, Y.J.; Kim, C.D.; Kim, Y.S.; Lee, J.H.; Im, M. Regulation of lipid production by acetylcholine signalling in human sebaceous glands. J. Dermatol. Sci. 2013, 72, 116–122. [Google Scholar] [CrossRef]
- Kurzen, H.; Wessler, I.; Kirkpatrick, C.J.; Kawashima, K.; Grando, S.A. The non-neuronal cholinergic system of human skin. Horm. Metab. Res. 2007, 39, 125–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurzen, H.; Berger, H.; Jäger, C.; Hartschuh, W.; Näher, H.; Gratchev, A.; Goerdt, S.; Deichmann, M. Phenotypical and molecular profiling of the extraneuronal cholinergic system of the skin. J. Investig. Dermatol. 2004, 123, 937–949. [Google Scholar] [CrossRef] [PubMed]
- Grando, S.A. Basic and clinical aspects of non-neuronal acetylcholine: Biological and clinical significance of non-canonical ligands of epithelial nicotinic acetylcholine receptors. J. Pharmacol. Sci. 2008, 106, 174–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grando, S.A.; Kawashima, K.; Kirkpatrick, C.J.; Kummer, W.; Wessler, I. Recent progress in revealing the biological and medical significance of the non-neuronal cholinergic system. Int. Immunopharmacol. 2015, 29, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Roh, M.; Han, M.; Kim, D.; Chung, K. Sebum output as a factor contributing to the size of facial pores. Br. J. Dermatol. 2006, 155, 890–894. [Google Scholar] [CrossRef]
- Sugiyama-Nakagiri, Y.; Naoe, A.; Ohuchi, A.; Kitahara, T. Serum levels of IGF-1 are related to human skin characteristics including the conspicuousness of facial pores. Int. J. Cosmet. Sci. 2011, 33, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Seok, J.; Jeong, S.Y.; Park, K.Y.; Li, K.; Seo, S.J. Facial pores: Definition, causes, and treatment options. Dermatol. Surg. 2016, 42, 277–285. [Google Scholar] [CrossRef]
- Briganti, S.; Flori, E.; Mastrofrancesco, A.; Ottaviani, M. Acne as an altered dermato-endocrine response problem. Exp. Dermatol. 2020, 29, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.M.; Gilliland, K.L.; Cong, Z.; Thiboutot, D.M. 13-cis Retinoic acid induces apoptosis and cell cycle arrest in human SEB-1 sebocytes. J. Investig. Dermatol. 2006, 126, 2178–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirazzini, M.; Montecucco, C.; Rossetto, O. Toxicology and pharmacology of botulinum and tetanus neurotoxins: An update. Arch. Toxicol. 2022, 96, 1521–1539. [Google Scholar] [CrossRef]
- Dong, M.; Masuyer, G.; Stenmark, P. Botulinum and tetanus neurotoxins. Annu. Rev. Biochem. 2019, 88, 811–837. [Google Scholar] [CrossRef] [PubMed]
- Lacy, D.B.; Tepp, W.; Cohen, A.C.; DasGupta, B.R.; Stevens, R.C. Crystal structure of botulinum neurotoxin type A and implications for toxicity. Nat. Struct. Biol. 1998, 5, 898–902. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Stenmark, P. The structure and classification of botulinum toxins. In Botulinum Neurotoxin Therapy. Handbook Of Experimental Pharmacology; Whitcup, S.M., Hallett, M., Eds.; Springer Nature: Berlin, Germany, 2019; Volume 263, pp. 11–33. [Google Scholar] [CrossRef]
- Steward, L.; Brin, M.F.; Brideau-Andersen, A. Novel native and engineered botulinum neurotoxins. Handb. Exp. Pharmacol. 2021, 263, 63–89. [Google Scholar] [CrossRef] [PubMed]
- Turton, K.; Chaddock, J.A.; Acharya, K.R. Botulinum and tetanus neurotoxins: Structure, function and therapeutic utility. Trends Biochem. Sci. 2002, 27, 552–558. [Google Scholar] [CrossRef]
- Brin, M.F. Basic and clinical aspects of BOTOX. Toxicon 2009, 54, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Chen, S. Clinical uses of botulinum neurotoxins: Current indications, limitations and future developments. Toxins 2012, 4, 913–939. [Google Scholar] [CrossRef] [Green Version]
- Aoki, K.R.; Francis, J. Updates on the antinociceptive mechanism hypothesis of botulinum toxin A. Parkinsonism Relat. Disord. 2011, 17 (Suppl. 1), S28–S33. [Google Scholar] [CrossRef]
- Shah, A.R. Use of intradermal botulinum toxin to reduce sebum production and facial pore size. J. Drugs Dermatol. 2008, 7, 847–850. [Google Scholar]
- Rose, A.E.; Goldberg, D.J. Safety and efficacy of intradermal injection of botulinum toxin for the treatment of oily skin. Dermatol. Surg. 2013, 39, 443–448. [Google Scholar] [CrossRef]
- Min, P.; Xi, W.; Grassetti, L.; Trisliana Perdanasari, A.; Torresetti, M.; Feng, S.; Su, W.; Pu, Z.; Zhang, Y.; Han, S.; et al. Sebum Production Alteration after Botulinum Toxin Type A Injections for the Treatment of Forehead Rhytides: A Prospective Randomized Double-Blind Dose-Comparative Clinical Investigation. Aesthet. Surg. J. 2015, 35, 600–610. [Google Scholar] [CrossRef]
- Kesty, K.; Goldberg, D.J. A Randomized, Double-Blinded Study Evaluating the Safety and Efficacy of AbobotulinumtoxinA Injections for Oily Skin of the Forehead: A Dose-Response Analysis. Dermatol. Surg. 2021, 47, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Rho, N.-K.; Gil, Y.-C. Botulinum neurotoxin type A in the treatment of facial seborrhea and acne: Evidence and a proposed mechanism. Toxins 2021, 13, 817. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, S.; Jacky, B.; Gallagher, C.J. Preventive, cumulative effects of botulinum toxin type A in facial aesthetics. Dermatol. Surg. 2017, 43, S244–S251. [Google Scholar] [CrossRef] [PubMed]
- Grando, S.A.; Zachary, C.B. The non-neuronal and nonmuscular effects of botulinum toxin: An opportunity for a deadly molecule to treat disease in the skin and beyond. Br. J. Dermatol. 2018, 178, 1011–1019. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Seltmann, H.; Neitzel, H.; Orfanos, C.E. Establishment and characterization of an immortalized human sebaceous gland cell line (SZ95). J. Investig. Dermatol. 1999, 113, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Thiboutot, D.; Jabara, S.; McAllister, J.M.; Sivarajah, A.; Gilliland, K.; Cong, Z.; Clawson, G. Human skin is a steroidogenic tissue: Steroidogenic enzymes and cofactors are expressed in epidermis, normal sebocytes, and an immortalized sebocyte cell line (SEB-1). J. Investig. Dermatol. 2003, 120, 905–914. [Google Scholar] [CrossRef] [Green Version]
- Hong, I.; Lee, M.-H.; Na, T.-Y.; Zouboulis, C.C.; Lee, M.-O. LXRalpha enhances lipid synthesis in SZ95 sebocytes. J. Investig. Dermatol. 2008, 128, 1266–1272. [Google Scholar] [CrossRef] [Green Version]
- Zouboulis, C.C.; Seltmann, H.; Abdel-Naser, M.B.; Hossini, A.M.; Menon, G.K.; Kubba, R. Effects of Extracellular Calcium and 1,25 dihydroxyvitamin D3 on Sebaceous Gland Cells In vitro and In vivo. Acta Derm. Venereol. 2017, 97, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, N.R.; Cong, Z.; Nelson, A.M.; Albert, A.J.; Rosamilia, L.L.; Sivarajah, S.; Gilliland, K.L.; Liu, W.; Mauger, D.T.; Gabbay, R.A.; et al. Peroxisome proliferator-activated receptors increase human sebum production. J. Investig. Dermatol. 2006, 126, 2002–2009. [Google Scholar] [CrossRef] [Green Version]
- Melnik, B.C.; Schmitz, G.; Zouboulis, C.C. Anti-acne agents attenuate FGFR2 signal transduction in acne. J. Investig. Dermatol. 2009, 129, 1868–1877. [Google Scholar] [CrossRef]
- Melnik, B.C. Role of FGFR2-signaling in the pathogenesis of acne. Dermato-Endocrinology 2009, 1, 141–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnik, B. Milk consumption: Aggravating factor of acne and promoter of chronic diseases of Western societies. J. Dtsch. Dermatol. Ges. 2009, 7, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Guagnano, V.; Furet, P.; Spanka, C.; Bordas, V.; Le Douget, M.; Stamm, C.; Brueggen, J.; Jensen, M.R.; Schnell, C.; Schmid, H.; et al. Discovery of 3-(2,6-dichloro-3,5-dimethoxy-phenyl)-1-{6-[4-(4-ethyl-piperazin-1-yl)-phenylamino]-pyrimidin-4-yl}-1-methyl-urea (NVP-BGJ398), a potent and selective inhibitor of the fibroblast growth factor receptor family of receptor tyrosine kinase. J. Med. Chem. 2011, 54, 7066–7083. [Google Scholar] [CrossRef] [PubMed]
- Ciruelas, K.; Marcotulli, D.; Bajjalieh, S.M. Synaptic vesicle protein 2: A multi-faceted regulator of secretion. Semin. Cell Dev. Biol. 2019, 95, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Jacky, B.P.S.; Garay, P.E.; Dupuy, J.; Nelson, J.B.; Cai, B.; Molina, Y.; Wang, J.; Steward, L.E.; Broide, R.S.; Francis, J.; et al. Identification of fibroblast growth factor receptor 3 (FGFR3) as a protein receptor for botulinum neurotoxin serotype A (BoNT/A). PLoS Pathog. 2013, 9, e1003369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elias, M.; Al-Saleem, F.; Ancharski, D.M.; Singh, A.; Nasser, Z.; Olson, R.M.; Simpson, L.L. Evidence that botulinum toxin receptors on epithelial cells and neuronal cells are not identical: Implications for development of a non-neurotropic vaccine. J. Pharmacol. Exp. Ther. 2011, 336, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Benoit, R.M.; Frey, D.; Hilbert, M.; Kevenaar, J.T.; Wieser, M.M.; Stirnimann, C.U.; McMillan, D.; Ceska, T.; Lebon, F.; Jaussi, R.; et al. Structural basis for recognition of synaptic vesicle protein 2C by botulinum neurotoxin A. Nature 2014, 505, 108–111. [Google Scholar] [CrossRef]
- James, N.G.; Malik, S.; Sanstrum, B.J.; Rhéaume, C.; Broide, R.S.; Jameson, D.M.; Brideau-Andersen, A.; Jacky, B.S. Characterization of clostridium botulinum neurotoxin serotype A (BoNT/A) and fibroblast growth factor receptor interactions using novel receptor dimerization assay. Sci. Rep. 2021, 11, 7832. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Description | Source |
|---|---|---|
| Human primary sebocyte donor #1 | Sebocytes were isolated from the facial skin of a Caucasian 66-year-old female donor under PREMARIN® (conjugated estrogen tablets, USP) treatment for menopause | ZenBio® (Durham, NC, USA) Cat# SEBM011514B |
| Human primary sebocyte donor #2 | Sebocytes were isolated from the facial skin of a Caucasian 59-year-old female overweight (body mass index: 27.6 kg/m2) donor | ZenBio® (Durham, NC, USA) Cat# SEBM022614B |
| SEB-1 immortalized sebocyte cell line | Sebocytes were isolated from the facial skin of a 55-year-old male donor and SV40 immortalized [47] | Dr. Diane Thiboutot |
| Name | Description | Reference |
|---|---|---|
| onabotA (BOTOX®, Allergan Aesthetics, an AbbVie company, Irvine, CA, USA) | Commercially available preparation of BoNT/A | |
| BoNT/A complex (Metabiologics, Madison, WI, USA) | Research grade botulinum neurotoxin type A, native complex; purified from Clostridia | |
| BoNT/A 150 kDa (Metabiologics, Madison, WI, USA) | Research grade botulinum neurotoxin type A, holotoxin, 150 kDa | |
| rHC/A | Recombinant binding domain of BoNT/A | [56] |
| rHC/A-WY | Recombinant binding domain of BoNT/A with mutations W1266L and Y1267S that prevent binding to gangliosides (sugar receptors for BoNT/A) | [56,57] |
| rHC/A-TT | Recombinant binding domain of BoNT/A with mutations T1145A and T1146A that reduces binding to synaptic vesicle protein 2 (SV2), a protein receptor for BoNT/A | [56,58] |
| rNTNH-HC/A | Recombinant binding domain homolog of nontoxic nonhemagglutinin (NTNH) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brami-Cherrier, K.; Chernavsky, A.; You, H.; Grando, S.A.; Brideau-Andersen, A.; Sondergaard, B. Botulinum Neurotoxin Type A Directly Affects Sebocytes and Modulates Oleic Acid-Induced Lipogenesis. Toxins 2022, 14, 708. https://doi.org/10.3390/toxins14100708
Brami-Cherrier K, Chernavsky A, You H, Grando SA, Brideau-Andersen A, Sondergaard B. Botulinum Neurotoxin Type A Directly Affects Sebocytes and Modulates Oleic Acid-Induced Lipogenesis. Toxins. 2022; 14(10):708. https://doi.org/10.3390/toxins14100708
Chicago/Turabian StyleBrami-Cherrier, Karen, Alex Chernavsky, Hui You, Sergei A. Grando, Amy Brideau-Andersen, and Birgitte Sondergaard. 2022. "Botulinum Neurotoxin Type A Directly Affects Sebocytes and Modulates Oleic Acid-Induced Lipogenesis" Toxins 14, no. 10: 708. https://doi.org/10.3390/toxins14100708