Bimodal Cell Size and Fusing Cells Observed in a Clonal Culture of the Ciguatoxin-Producing Benthic Dinoflagellate Gambierdiscus (WC1/1)


Abstract
1. Introduction
2. Results and Discussion
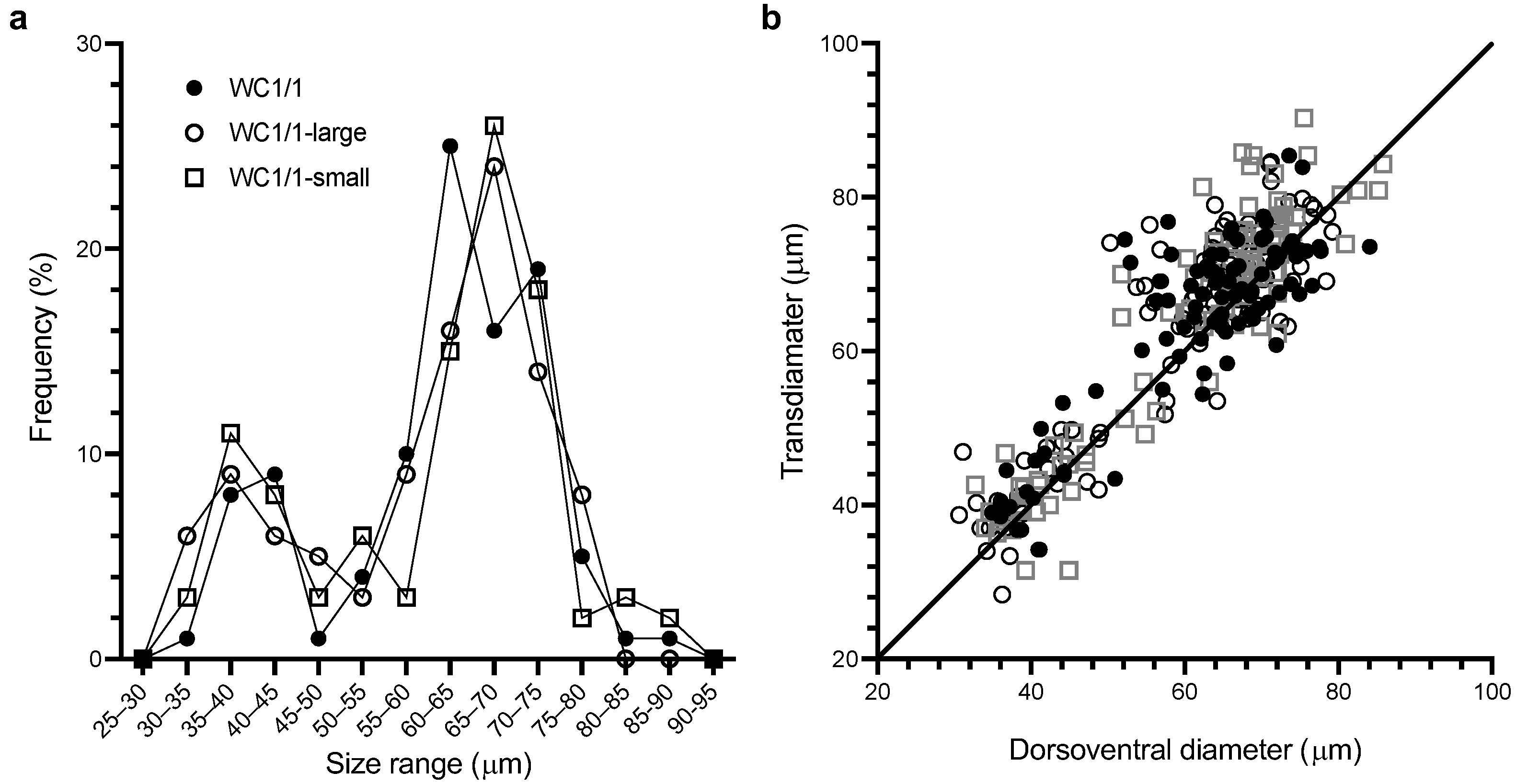
2.1. A bimodal Cell Size Produced by Gambierdiscus in Culture
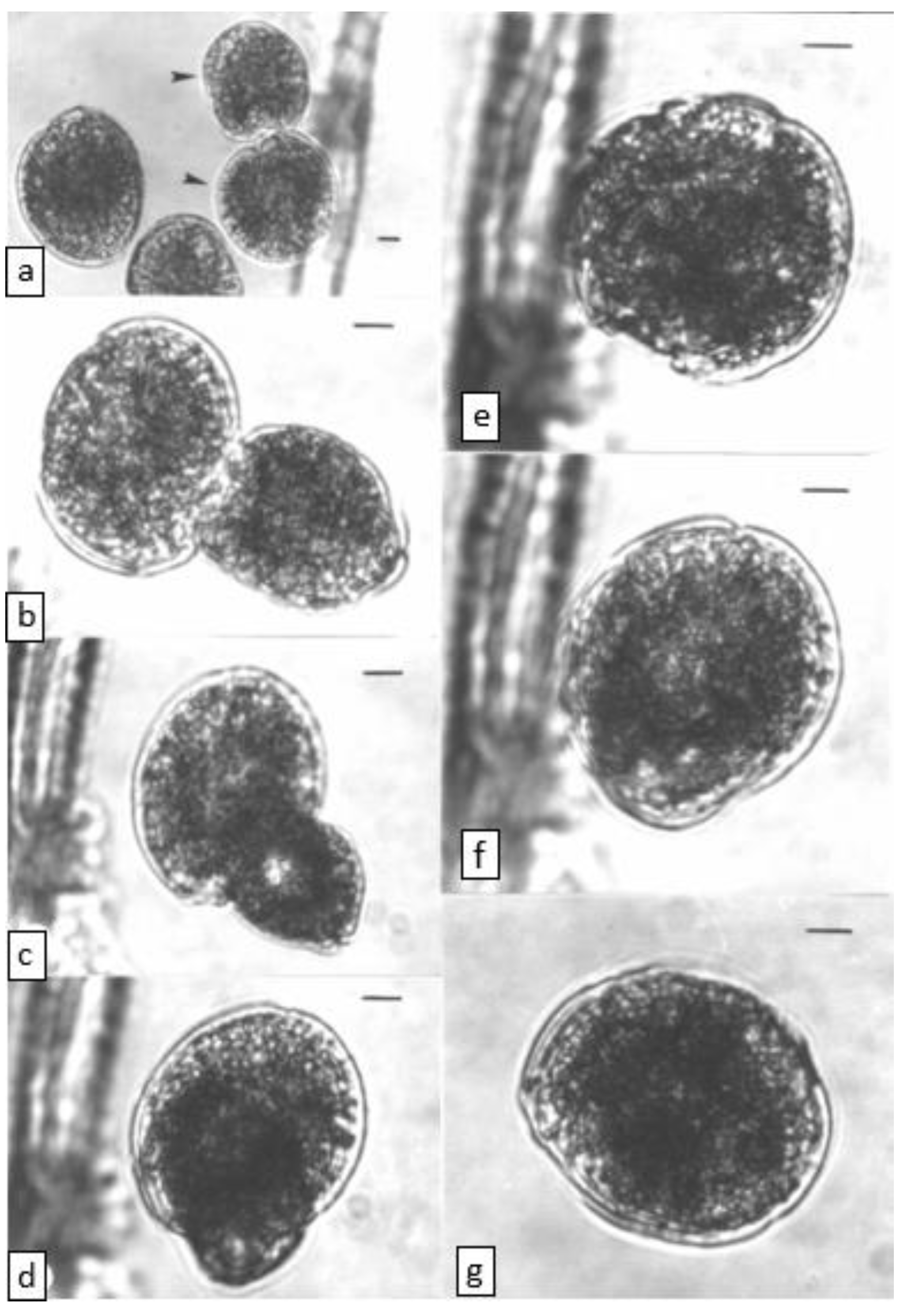
2.2. Cell Fusion (Light Microscope Observations)
2.3. Toxicity
3. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO; WHO. Report of the Expert Meeting on Ciguatera Poisoning: Rome, 19–23 November 2018; Food Safety and Quality No. 9; FAO: Rome, Italy, 2020. [CrossRef]
- Holmes, M.J.; Venables, B.; Lewis, R.J. Critical review and conceptual and quantitative models for the transfer and depuration of ciguatoxins in fishes. Toxins 2021, 13, 515. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.J.; Lewis, R.J. Origin of ciguateric fish: Quantitative modelling of the flow of ciguatoxin through a marine food chain. Toxins 2022, 14, 534. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Bagnis, R.; Vernoux, J.-P. Toxicity of the surgeonfishes –II Properties of the principal water-soluble toxin. Bull. Jpn. Soc. Sci. Fish. 1976, 42, 359–365. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J.; Gillespie, N.C. Toxicity of Australian and French Polynesian strains of Gambierdiscus toxicus (Dinophyceae) grown in culture: Characterization of a new type of maitotoxin. Toxicon 1990, 28, 1159–1172. [Google Scholar] [CrossRef]
- Ngai, H.; Torigoe, K.; Satake, M.; Murata, M.; Yasumoto, T.; Hirota, H. Gambieric acids: Unprecedented potent antifungal substances isolated from cultures of a marine dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1992, 114, 1102–1103. [Google Scholar] [CrossRef]
- Murata, M.; Naoki, H.; Iwashita, T.; Matsunaga, S.; Sasaki, M.; Yokoyama, A.; Yasumoto, T. Structure of maitotoxin. J. Am. Chem. Soc. 1993, 115, 2060–2062. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J. Purification and characterization of large and small maitotoxins from cultured Gambierdiscus toxicus. Nat. Toxins 1994, 2, 64–72. [Google Scholar] [CrossRef]
- Rodríguez, I.; Genta-Jouve, G.; Alfonso, C.; Calabro, K.; Alonso, E.; Sánchez, J.A.; Alfonso, A.; Oliver, T.P.; Botana, L.M. Gambierone, a ladder-shaped polyether from the dinoflagellate Gambierdiscus belizeanus. Org. Lett. 2015, 17, 2392–2395. [Google Scholar] [CrossRef]
- Pisapia, F.; Sibat, M.; Herrenknecht, C.; Lhaute, K.; Gaini, G.; Ferron, P.-J.; Fessard, V.; Fraga, S.; Nascimento, S.M.; Litaker, R.W.; et al. Maitotoxin-4, a novel MTX analog produced by Gambierdiscus excentricus. Mar. Drugs 2017, 15, 220. [Google Scholar] [CrossRef]
- Murray, J.S.; Selwood, A.I.; Harwood, D.T.; van Ginkel, R.; Puddick, J.; Rhodes, L.L.; Rise, F.; Wilkins, A.L. 44-Methylgambierone, a new gambierone analogue isolated from Gambierdiscus australes. Tetrahedron Lett. 2019, 60, 621–625. [Google Scholar] [CrossRef]
- Boente-Juncal, A.; Álvarez, M.; Antelo, Á.; Rodríguez, I.; Calabro, K.; Vale, C.; Thomas, O.P.; Botana, L.M. Structure elucidation and biological evaluation of maitotoxin-3, a homologue of gambierone, from Gambieridiscus belizeanus. Toxins 2019, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Estevez, P.; Castro, D.; Leão-Martins, J.M.; Sibat, M.; Tudó, A.; Dickey, R.; Diogene, J.; Hess, P.; Gago-Martinez, A. Toxicity screening of a Gambierdiscus australes strain from the Western Mediterranean Sea and identification of a novel maitotoxin analogue. Mar. Drugs 2021, 19, 460. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.S.; Finch, S.C.; Mudge, E.M.; Wilkins, A.L.; Puddick, J.; Harwood, D.T.; Rhodes, L.L.; van Ginkel, R.; Rise, F.; Prinsep, M.R. Structural characterization of maitotoxins produced by toxic Gambierdiscus species. Mar. Drugs 2022, 20, 453. [Google Scholar] [CrossRef]
- Adachi, R.; Fukuyo, Y. The thecal structure of a marine toxic dinoflagellate Gambierdiscus toxicus gen. et sp. nov. collected in a ciguatera endemic area. Bull. Jpn. Soc. Sci. Fish. 1979, 45, 67–71. [Google Scholar] [CrossRef]
- Bagnis, R.; Chanteau, S.; Chungue, E.; Hurtel, J.M.; Yasumoto, T.; Inoue, A. Origins of ciguatera fish poisoning: A new dinoflagellate, Gambierdiscus toxicus Adachi et Fukuyo, definitely involved as a causal agent. Toxicon 1980, 18, 198–208. [Google Scholar] [CrossRef]
- Durand, M.; Pusieux-Dao, S. Physiological and ultrastructural features of the toxic dinoflagellate Gambierdiscus toxicus in culture. In Toxic Dinoflagellates; Anderson, D.M., White, A.W., Baden, D.G., Eds.; Elsevier: Oxford, UK, 1985; pp. 61–68. [Google Scholar]
- Pfiester, L.A.; Anderson, D.A. Dinoflagellate reproduction. In The Biology of Dinoflagellates; Taylor, F.J.R., Ed.; Blackwell: Oxford, UK, 1987; pp. 611–648. [Google Scholar]
- Taylor, F.J.R. A description of the benthic dinoflagellate associated with maitotoxin and ciguatoxin, including observations on Hawaiian material. In Toxic Dinoflagellate Blooms; Taylor, D.L., Seliger, H.H., Eds.; Elsevier: North Holland, The Netherlands, 1979; pp. 71–76. [Google Scholar]
- Bravo, I.; Figueroa, R.I.; Fraga, S. Cellular and nuclear morphological variability within a single species of the toxigenic dinoflagellate genus Gambierdiscus: Relationship to life-cycle processes. Harmful Algae 2014, 40, 1–8. [Google Scholar] [CrossRef]
- Ramilo, I.; Figueroa, R.I.; Rayón-Viña, F.; Cuadrado, Á.; Bravo, I. Temperature-dependent growth and sexuality of the ciguatoxin producer dinoflagellate Gambierdiscus spp. in cultures established from the Canary Islands. Harmful Algae 2021, 110, 102130. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J.; Poli, M.A.; Gillespie, N.C. Strain dependent production of ciguatoxin precursors (gambiertoxins) by Gambierdiscus toxicus (Dinophyceae) in culture. Toxicon 1991, 29, 761–775. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J. Multiple gambiertoxins (ciguatoxin precursors) from an Australian strain of Gambierdiscus toxicus in culture. In Recent Advances in Toxinology Research; Gopalakrishnakone, P., Tan, C.K., Eds.; National University of Singapore: Singapore, 1992; Volume 2, pp. 520–529. [Google Scholar]
- Lewis, R.J.; Holmes, M.J.; Alewood, P.F.; Jones, A. Ionspray mass spectrometry of ciguatoxin-1, maitotoxin-2 and -3, and related marine polyether toxins. Nat. Toxins 1994, 2, 56–63. [Google Scholar] [CrossRef]
- GEOHAB. Global Ecology and Oceanography of Harmful Algal Blooms, GEOHAB Core Research Project: HABs in Benthic Systems; Berdalet, E., Tester, P., A. Zingone, A., Eds.; IOC of UNESCO and SCOR: Paris, France; Newark, NJ, USA, 2012; p. 64. [Google Scholar]
- Taylor, F.J.R. Dinoflagellate morphology. In The Biology of Dinoflagellates; Taylor, F.J.R., Ed.; Blackwell: Oxford, UK, 1987; pp. 24–91. [Google Scholar]
- Gaines, G.; Elbrächter, M. Heterotrophic nutrition. In The Biology of Dinoflagellates; Taylor, F.J.R., Ed.; Blackwell: Oxford, UK, 1987; pp. 224–268. [Google Scholar]
- Schnepf, E.; Deichgräber, G. “Myzocytosis”, a kind of endocytosis with implications to compartmentation in endosymbiosis. Naturwissenschaften 1984, 71, 218–219. [Google Scholar] [CrossRef]
- Yang, H.; Hu, Z.; Shang, L.; Deng, Y.; Tang, Y.Z. A strain of the toxic dinoflagellate Karlodinium veneficum isolated from the East China Sea is an omnivorous phagotroph. Harmful Algae 2020, 93, 101775. [Google Scholar] [CrossRef] [PubMed]
- Von Stosch, H.A. Observations on vegetative reproduction and sexual life cycles of two freshwater dinoflagellates, Gymnodinium pseudopalustre Schiller and Woloszynskia apiculata sp. nov. Br. Phycol. J. 1973, 8, 105–134. [Google Scholar] [CrossRef]
- Bhaud, Y.; Soyer-Gobillard, M.-O.; Salmon, J.M. Transmission of gametic nuclei through a fertilization tube during mating in a primitive dinoflagellate Prorocentrum micans Ehr. J. Cell Sci. 1988, 89, 197–206. [Google Scholar] [CrossRef]
- Faust, M.A. Observations on the morphology and sexual reproduction of Coolia monotis (Dinophyceae). J. Phycol. 1992, 28, 94–104. [Google Scholar]
- Jeong, H.J.; Latz, M.J. Growth and grazing rates of the heterotrophic dinoflagellates Protoperidinium spp. on red tide dinoflagellates. Mar. Ecol. Prog. Ser. 1994, 106, 173–185. [Google Scholar] [CrossRef]
- Faust, M.A. Mixotrophy in tropical benthic dinoflagellates. In Harmful Algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; Xunta de Galicia and International Oceanographic Commission of UNESCO: Santiago de Compostela, Spain, 1998; pp. 390–393. [Google Scholar]
- Price, D.C.; Farinholt, N.; Gates, C.; Schumaker, A.; Wagner, N.E.; Bienfang, P.; Bhattacharya, D. Analysis of Gambierdiscus transcriptome data supports ancient origins of mixotrophic pathways in dinoflagellates. Environ. Microbiol. 2016, 18, 4501–4510. [Google Scholar] [CrossRef]
- Holmes, M.J. Gambierdiscus yasumotoi sp. nov. (Dinophyceae), a toxic, benthic dinoflagellates from southeastern Asia. J. Phycol. 1998, 34, 661–668. [Google Scholar] [CrossRef]
- Chinain, M.; Faust, M.A.; Pauillac, S. Morphology and molecular analyses of three toxic species of Gambierdiscus (Dinophyceae): G. pacificus, sp. nov., G. australes, sp. nov., and G. polynesiensis, sp. nov. J. Phycol. 1999, 35, 1282–1296. [Google Scholar] [CrossRef]
- Macdonald, P.D.M.; Pitcher, T.J. Age-groups from size-frequency data: A versatile and efficient method of analyzing distribution mixtures. J. Fish Res. Bd. Can. 1979, 36, 987–1001. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Clonal Culture | Dorsoventral Diameter (µm) and Percentage of Cells in Each Mode in Brackets | Transdiameter (µm) and Percentage of Cells Consistent with Each Mode in Brackets | ||
|---|---|---|---|---|
| Small Mode | Large Mode | Small mode | Large mode | |
| WC1/1-large | 40 ± 2 (28%) | 66 ± 1 (72%) | 42 ± 1 (29%) | 70 ± 1 (71%) |
| WC1/1-small | 39 ± 1 (26%) | 67 ± 1 (74%) | 41 ± 1 (28%) | 71 ± 1 (72%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holmes, M.J.; Lewis, R.J. Bimodal Cell Size and Fusing Cells Observed in a Clonal Culture of the Ciguatoxin-Producing Benthic Dinoflagellate Gambierdiscus (WC1/1). Toxins 2022, 14, 767. https://doi.org/10.3390/toxins14110767
Holmes MJ, Lewis RJ. Bimodal Cell Size and Fusing Cells Observed in a Clonal Culture of the Ciguatoxin-Producing Benthic Dinoflagellate Gambierdiscus (WC1/1). Toxins. 2022; 14(11):767. https://doi.org/10.3390/toxins14110767
Chicago/Turabian StyleHolmes, Michael J., and Richard J. Lewis. 2022. "Bimodal Cell Size and Fusing Cells Observed in a Clonal Culture of the Ciguatoxin-Producing Benthic Dinoflagellate Gambierdiscus (WC1/1)" Toxins 14, no. 11: 767. https://doi.org/10.3390/toxins14110767
APA StyleHolmes, M. J., & Lewis, R. J. (2022). Bimodal Cell Size and Fusing Cells Observed in a Clonal Culture of the Ciguatoxin-Producing Benthic Dinoflagellate Gambierdiscus (WC1/1). Toxins, 14(11), 767. https://doi.org/10.3390/toxins14110767