The Preferential Therapeutic Potential of Chlorella vulgaris against Aflatoxin-Induced Hepatic Injury in Quail

 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Growth Performance
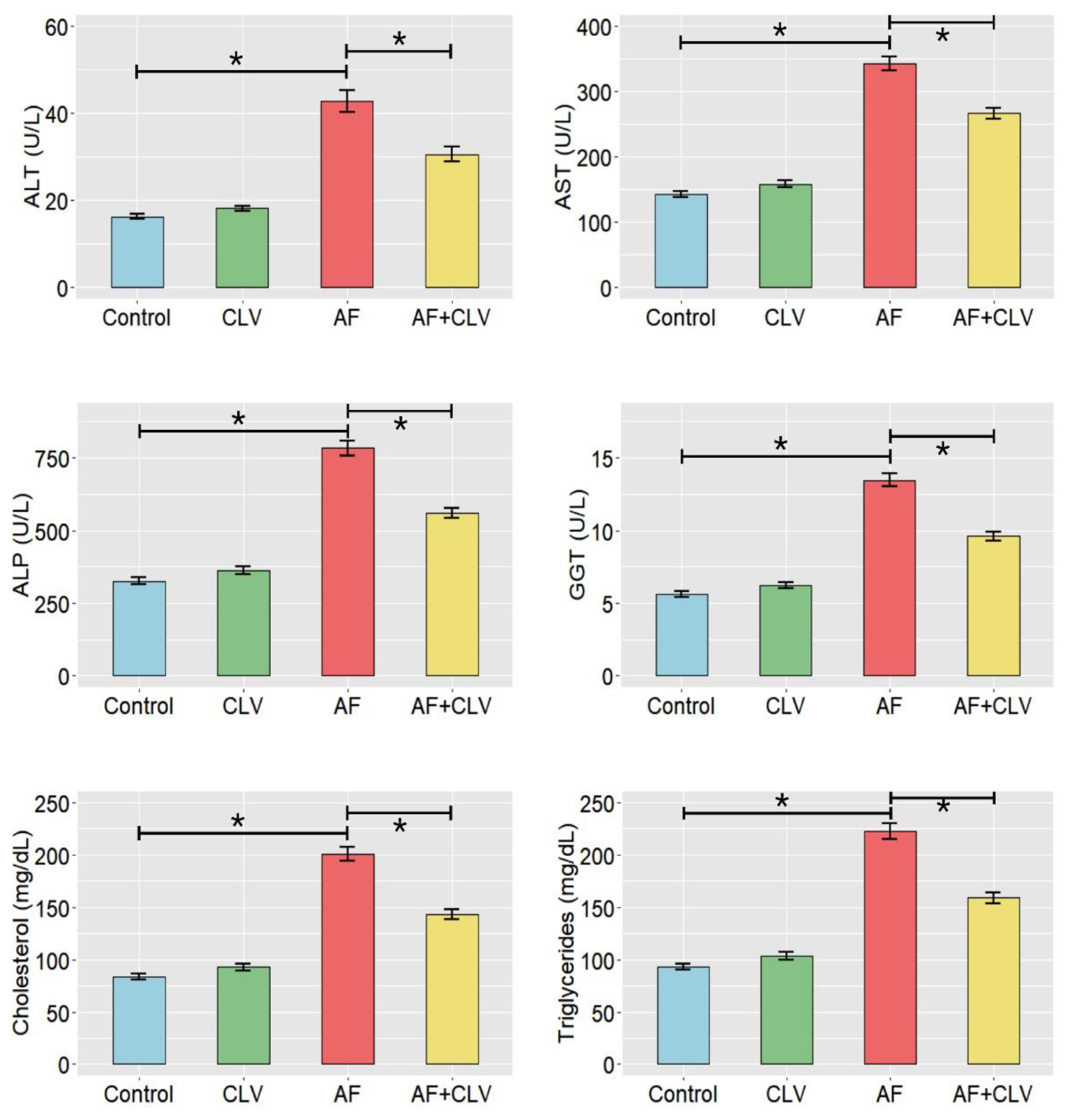
2.2. Changes in Liver Function Indices and Lipid Profile
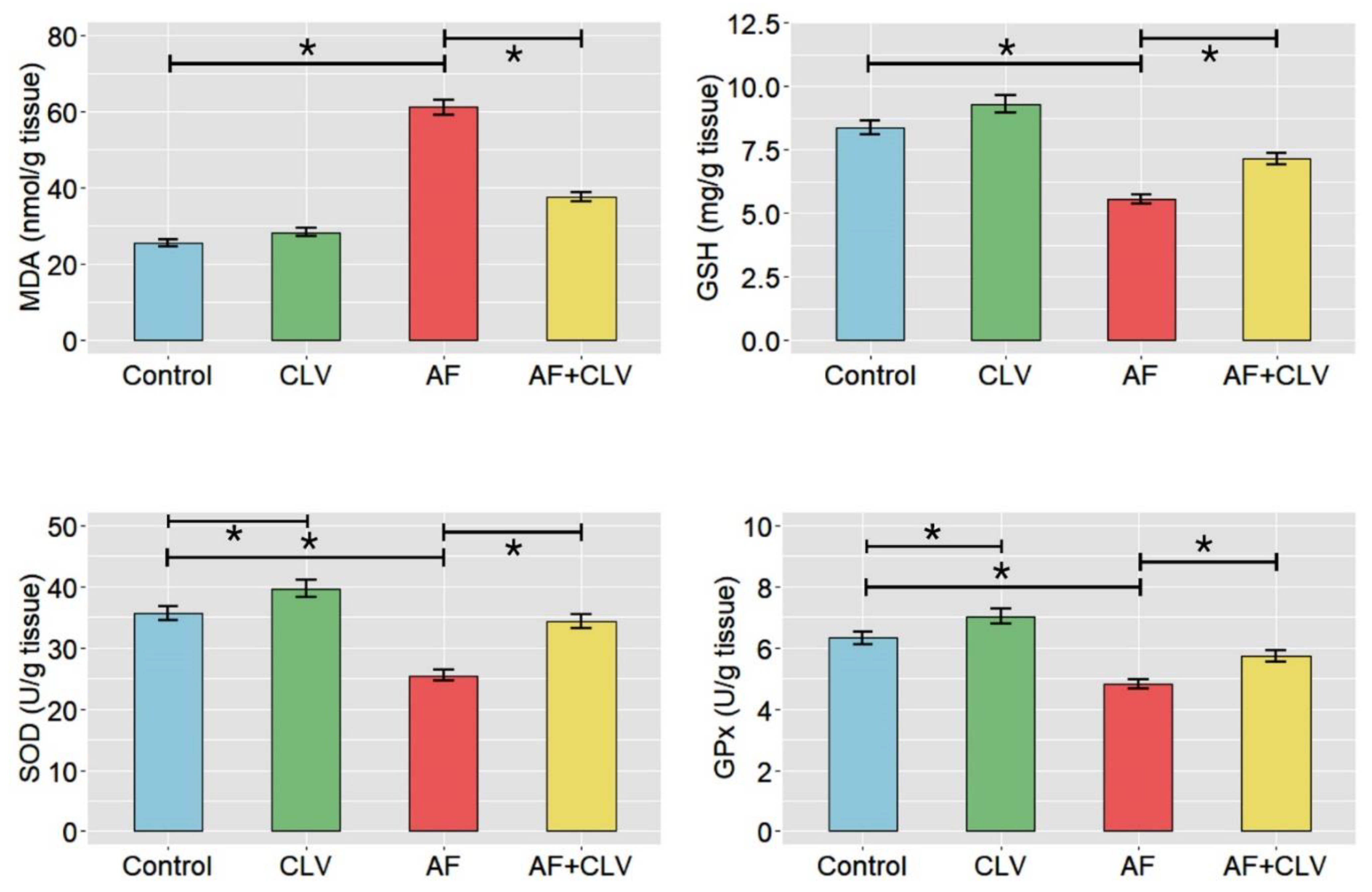
2.3. Changes in Hepatic Oxidant/Antioxidant Hemostasis
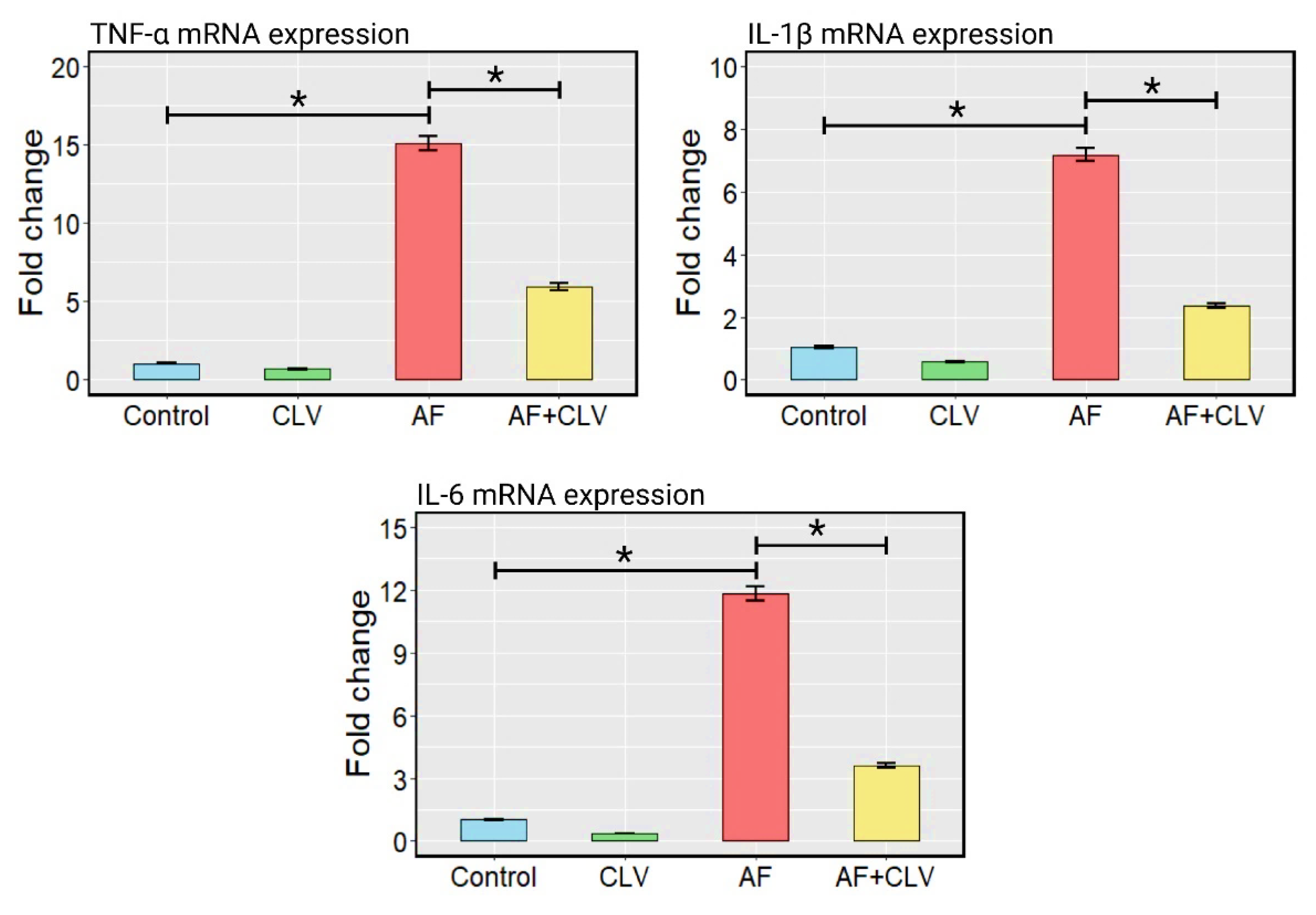
2.4. Changes in Inflammatory Cytokine mRNA Expressions
2.5. Changes in Meat Nutritive Value after C. vulgaris and/or Aflatoxin Exposure
2.6. Impact of C. vulgaris on Aflatoxin Residues in Liver Tissue and Meat
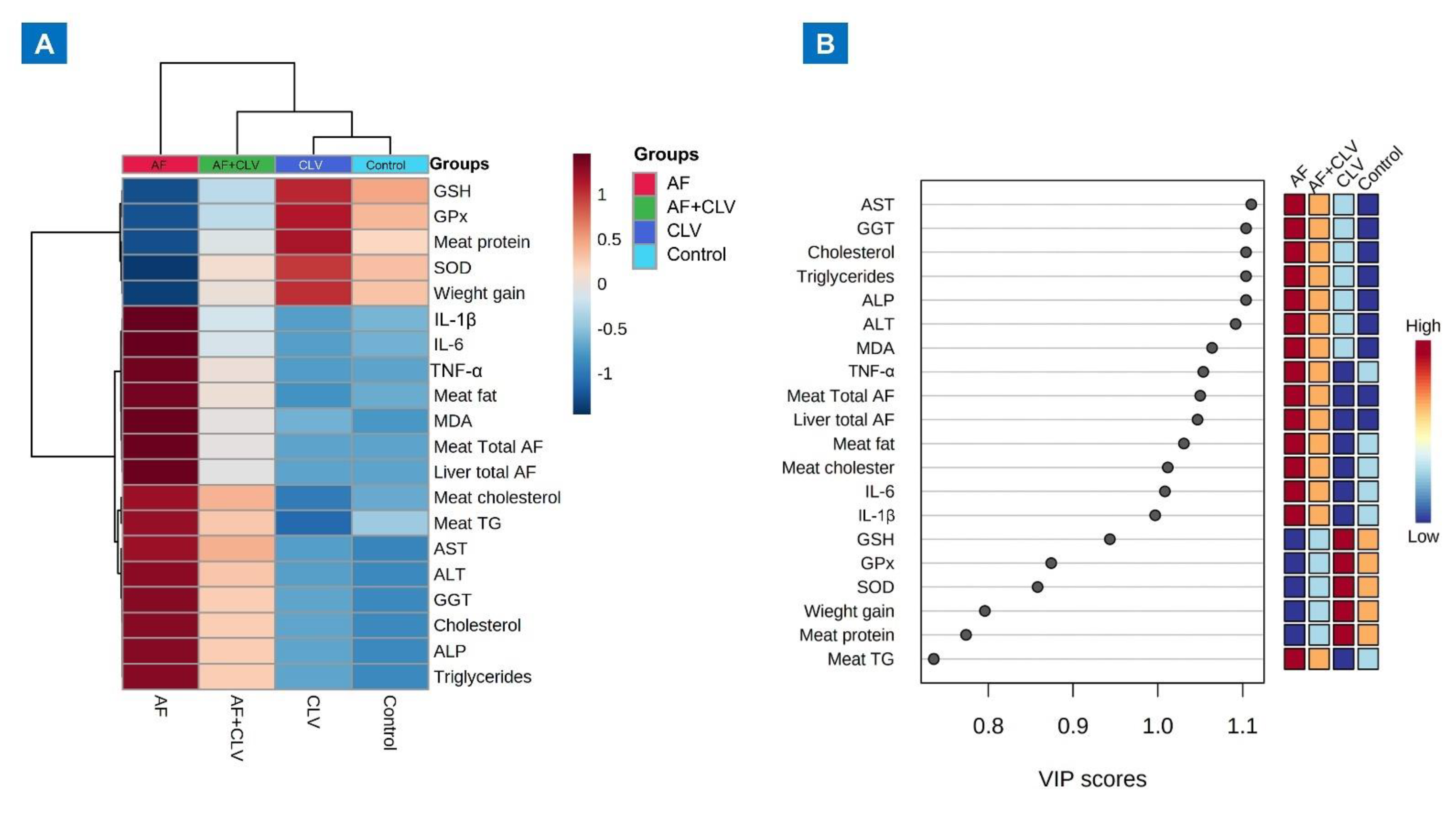
2.7. Hierarchical Clustering Heatmap and Variable Importance in Projection (VIP) Score
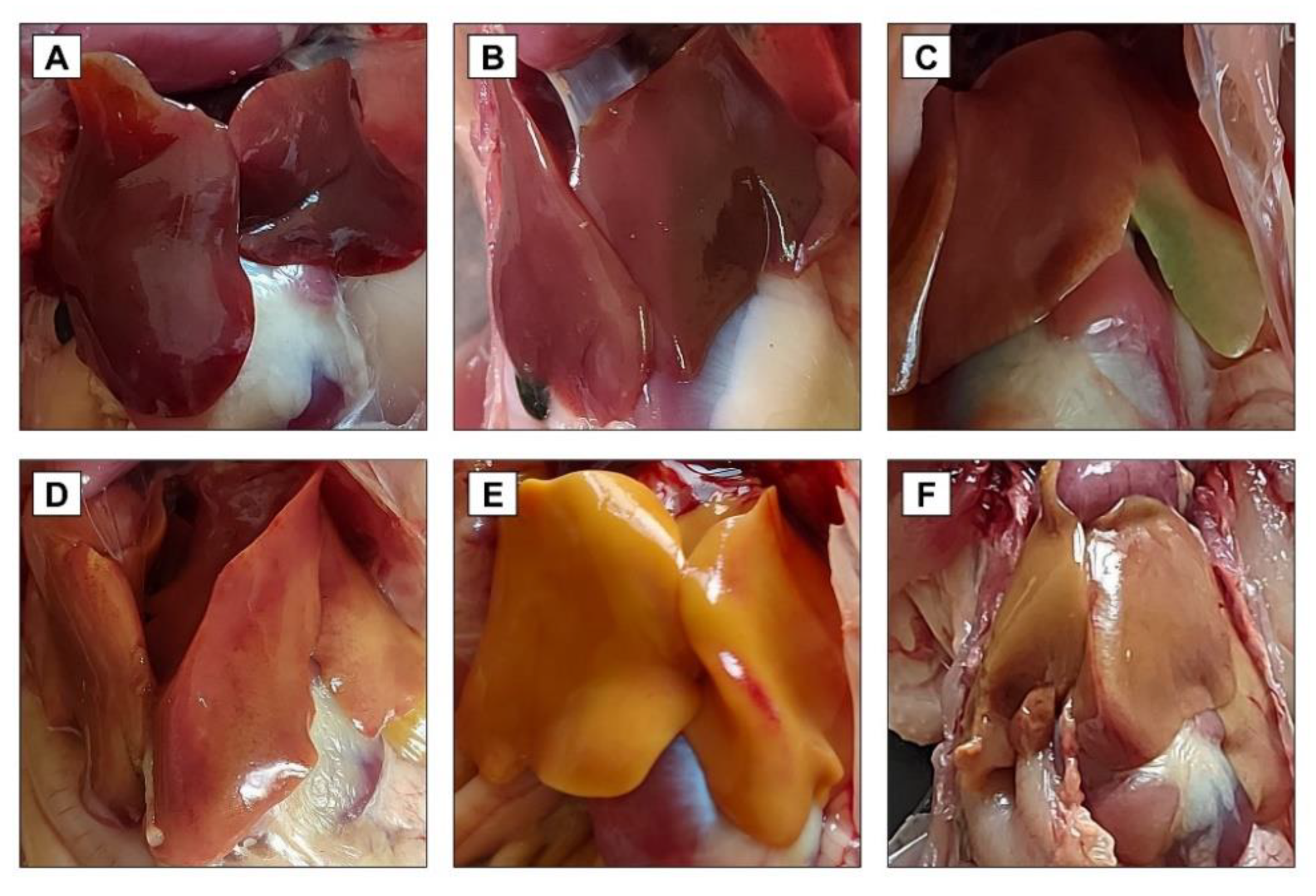
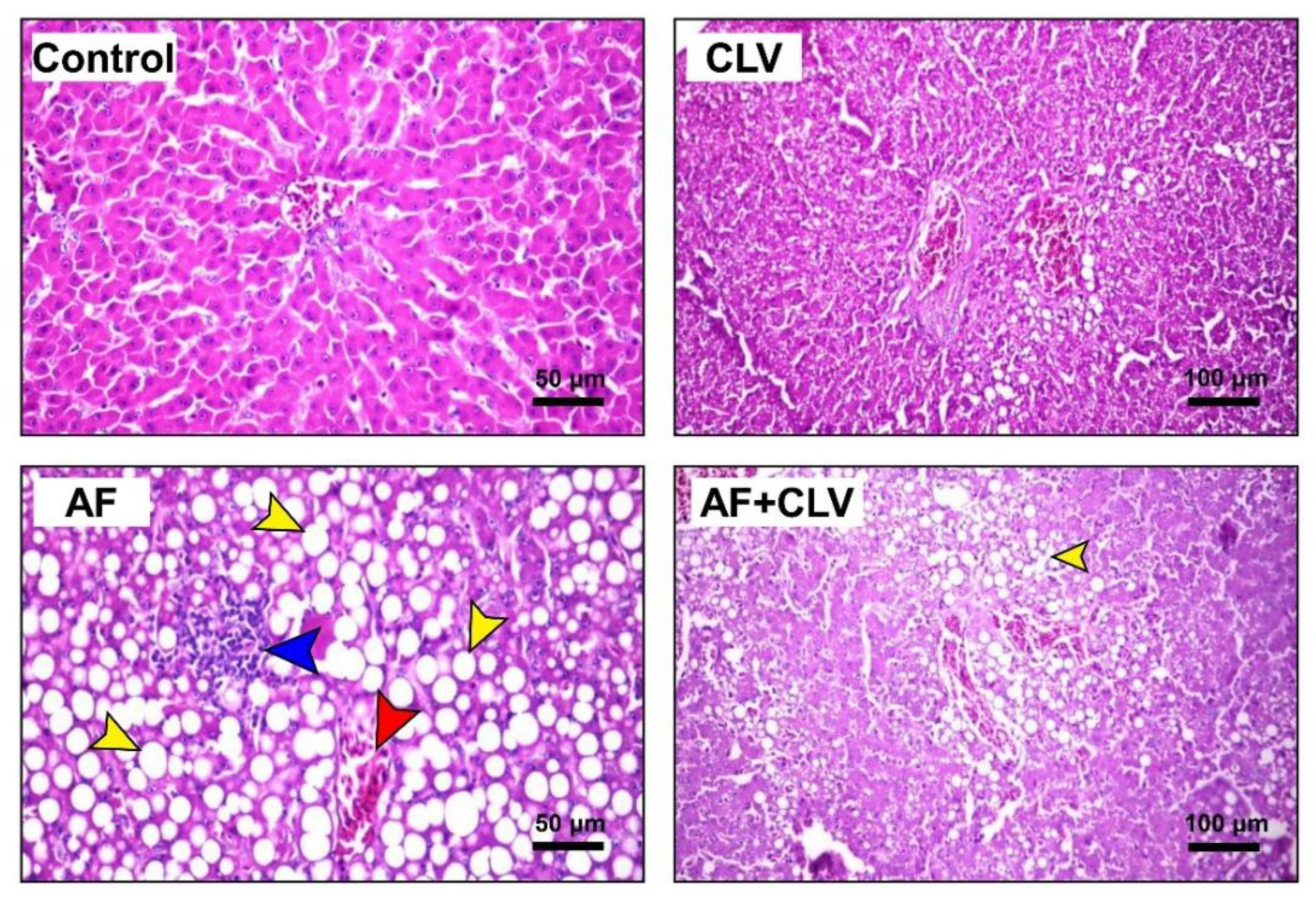
2.8. Clinical, Postmortem, and Histopathological Examination
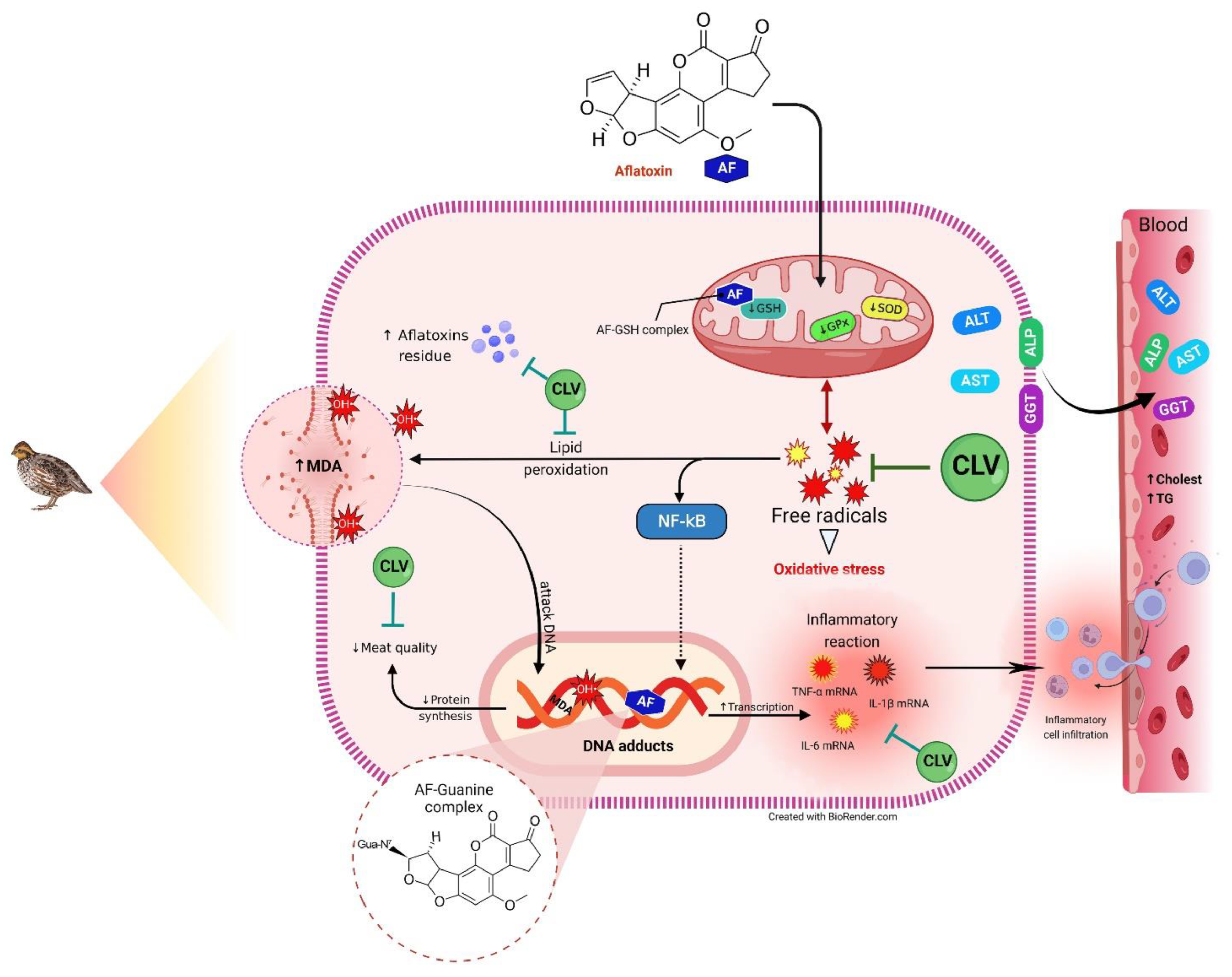
3. Discussion
4. Conclusions
5. Materials and Methods
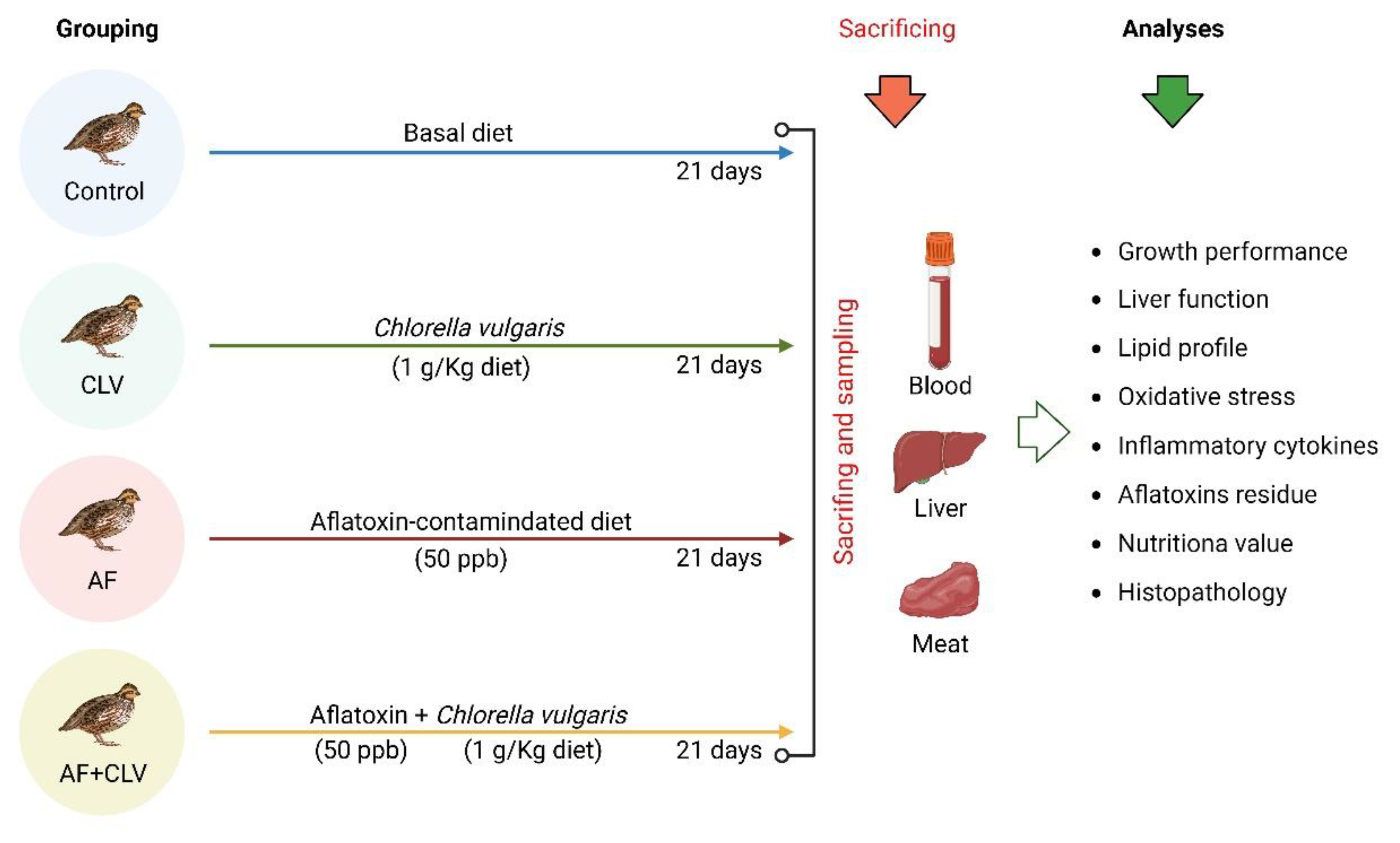
5.1. Experimental Design
5.2. Growth Performance Assessment
5.3. Biochemical Analyses
5.4. Quantitative Real-Time PCR (qRT-PCR)
5.5. Evaluation of Meat Nutritive Value
5.6. Aflatoxin Residues in Liver Tissue and Meat
5.7. Postmortem and Histopathological Examination
5.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, M.; Jiao, P.; Wang, X.; Sun, Y.; Liang, G.; Xie, X.; Zhang, Y. Evaluation of Growth Performance, Nitrogen Balance and Blood Metabolites of Mutton Sheep Fed an Ammonia-Treated Aflatoxin B1-Contaminated Diet. Toxins 2022, 14, 361. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; El-Rayes, S.M.; Khalil, W.F.; Abdeen, A.; Abdelkader, A.; Youssef, M.; Maher, Z.M.; Ibrahim, A.N.; Abdelrahman, S.M.; Ibrahim, S.F.; et al. Arabic Gum Could Alleviate the Aflatoxin B1-Provoked Hepatic Injury in Rat: The Involvement of Oxidative Stress, Inflammatory, and Apoptotic Pathways. Toxins 2022, 14, 605. [Google Scholar] [CrossRef] [PubMed]
- Medina, Á.; González-Jartín, J.M.; Sainz, M.J. Impact of Global Warming on Mycotoxins. Curr. Opin. Food Sci. 2017, 18, 76–81. [Google Scholar] [CrossRef]
- Saleemi, M.K.; Ashraf, K.; Gul, S.T.; Naseem, M.N.; Sajid, M.S.; Mohsin, M.; He, C.; Zubair, M.; Khan, A. Toxicopathological Effects of Feeding Aflatoxins B1 in Broilers and Its Ameliosration with Indigenous Mycotoxin Binder. Ecotoxicol. Environ. Saf. 2020, 187, 109712. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human Aflatoxicosis in Developing Countries: A Review of Toxicology, Exposure, Potential Health Consequences, and Interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef] [PubMed]
- da Silva Cardoso, V.; Vermelho, A.B.; Ribeiro de Lima, C.A.; Mendes de Oliveira, J.; Freire de Lima, M.E.; Pinto da Silva, L.H.; Direito, G.M.; Miranda Danelli, M.d.G. Antigenotoxic Effect of Piperine in Broiler Chickens Intoxicated with Aflatoxin B1. Toxins 2016, 8, 316. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.; Dawood, M.A.O.; AlKahtane, A.A.; Abdeen, A.; Abdel-Latif, H.M.R.; Senousy, H.H.; Aleya, L.; Alkahtani, S. Spirulina platensis Mediated the Biochemical Indices and Antioxidative Function of Nile Tilapia (Oreochromis niloticus) Intoxicated with Aflatoxin B1. Toxicon 2020, 184, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.M.; Abdeen, A.; Jalouli, M.; Abdelkader, A.; Megahed, A.; Alkahtane, A.; Almeer, R.; Alhoshani, N.M.; Al-Johani, N.S.; Alkahtani, S.; et al. Fucoidan Supplementation Modulates Hepato-Renal Oxidative Stress and DNA Damage Induced by Aflatoxin B1 Intoxication in Rats. Sci. Total Environ. 2021, 768, 144781. [Google Scholar] [CrossRef]
- Bhatti, S.A.; Khan, M.Z.; Saleemi, M.K.; Saqib, M.; Khan, A.; Ul-Hassan, Z. Protective Role of Bentonite against Aflatoxin B1-and Ochratoxin A-Induced Immunotoxicity in Broilers. J. Immunotoxicol. 2017, 14, 66–76. [Google Scholar] [CrossRef]
- Yilmaz, S.; Bag, H. Aflatoxin B1: Mechanism, Oxidative Stress, and Effects on Animal Health. Insights Vet. Sci. 2022, 6, 17–24. [Google Scholar] [CrossRef]
- Zhu, F.; Zhao, X.; Li, J.; Guo, L.; Bai, L.; Qi, X. A New Compound Trichomicin Exerts Antitumor Activity through STAT3 Signaling Inhibition. Biomed. Pharmacother. 2020, 121, 109608. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, R.; Xia, S.; Wei, G.; Ishfaq, M.; Zhang, Y.; Zhang, X. Protective Role of Curcumin on Aflatoxin B1-Induced TLR4/RIPK Pathway Mediated-Necroptosis and Inflammation in Chicken Liver. Ecotoxicol. Environ. Saf. 2022, 233, 113319. [Google Scholar] [CrossRef]
- Kolosova, A.; Stroka, J. Evaluation of the Effect of Mycotoxin Binders in Animal Feed on the Analytical Performance of Standardised Methods for the Determination of Mycotoxins in Feed. Food Addit. Contam. Part A 2012, 29, 1959–1971. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, F.; Zhou, X.; Liu, M.; Zang, H.; Liu, X.; Shan, A.; Feng, X. Alleviation of Oral Exposure to Aflatoxin B1-Induced Renal Dysfunction, Oxidative Stress, and Cell Apoptosis in Mice Kidney by Curcumin. Antioxidants 2022, 11, 1082. [Google Scholar] [CrossRef]
- Palade, L.M.; Dore, M.I.; Marin, D.E.; Rotar, M.C.; Taranu, I. Assessment of Food By-Products’ Potential for Simultaneous Binding of Aflatoxin B1 and Zearalenone. Toxins 2021, 13, 2. [Google Scholar] [CrossRef]
- Andrade, L.M.; Andrade, C.J.; Dias, M.; Nascimento, C.; Mendes, M.A. Chlorella and Spirulina Microalgae as Sources of Functional Foods. Nutraceuticals Food Suppl. 2018, 6, 45–58. [Google Scholar]
- Abdelnour, S.A.; Sheiha, A.M.; Taha, A.E.; Swelum, A.A.; Alarifi, S.; Alkahtani, S.; Ali, D.; AlBasher, G.; Almeer, R.; Falodah, F. Impacts of Enriching Growing Rabbit Diets with Chlorella Vulgaris Microalgae on Growth, Blood Variables, Carcass Traits, Immunological and Antioxidant Indices. Animals 2019, 9, 788. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.E.; Naguib, M.M.; Higazy, A.M.; Sultan, Y.Y.; Marrez, D.A. Effect of Substitution Soybean by Blue Green Alga Spirulina platensis on Performance and Meat Quality of Growing Rabbits. Am. J. Food Technol. 2017, 12, 51–59. [Google Scholar] [CrossRef]
- Abdelhamid, F.M.; Elshopakey, G.E.; Aziza, A.E. Ameliorative Effects of Dietary Chlorella vulgaris and β-Glucan against Diazinon-Induced Toxicity in Nile Tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2020, 96, 213–222. [Google Scholar] [CrossRef]
- Wen, Z.G.; Du, Y.K.; Xie, M.; Li, X.M.; Wang, J.D.; Yang, P.L. Effects of Low-Protein Diets on Growth Performance and Carcass Yields of Growing French Meat Quails (France coturnix coturnix). Poult. Sci. 2017, 96, 1364–1369. [Google Scholar] [CrossRef]
- Chen, X.; Abdallah, M.F.; Grootaert, C.; Rajkovic, A. Bioenergetic Status of the Intestinal and Hepatic Cells after Short Term Exposure to Fumonisin B1 and Aflatoxin B1. Int. J. Mol. Sci. 2022, 23, 6945. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Liu, Y.; Guo, Y.; Ma, Q.; Ji, C.; Zhao, L. Transcriptional Profiling of Aflatoxin B1-Induced Oxidative Stress and Inflammatory Response in Macrophages. Toxins 2021, 13, 401. [Google Scholar] [CrossRef] [PubMed]
- Aleissa, M.S.; Alkahtani, S.; Abd Eldaim, M.A.; Ahmed, A.M.; Bungǎu, S.G.; Almutairi, B.; Bin-Jumah, M.; Alkahtane, A.A.; Alyousif, M.S.; Abdel-Daim, M.M.; et al. Fucoidan Ameliorates Oxidative Stress, Inflammation, DNA Damage, and Hepatorenal Injuries in Diabetic Rats Intoxicated with Aflatoxin B1. Oxid. Med. Cell. Longev. 2020, 2020, 9316751. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, S.; Kaya, E.; Kisacam, M.A. The Effect on Oxidative Stress of Aflatoxin and Protective Effect of Lycopene on Aflatoxin Damage. Aflatoxin-Control Anal. Detect. Health Risks 2017, 30, 67–90. [Google Scholar] [CrossRef]
- Aboubakr, M.; Elshafae, S.M.; Abdelhiee, E.Y.; Fadl, S.E.; Soliman, A.; Abdelkader, A.; Abdel-Daim, M.M.; Bayoumi, K.A.; Baty, R.S.; Elgendy, E.; et al. Antioxidant and Anti-Inflammatory Potential of Thymoquinone and Lycopene Mitigate the Chlorpyrifos-Induced Toxic Neuropathy. Pharmaceuticals 2021, 14, 940. [Google Scholar] [CrossRef]
- Allam, A.; Abdeen, A.; Devkota, H.P.; Ibrahim, S.S.; Youssef, G.; Soliman, A.; Abdel-Daim, M.M.; Alzahrani, K.J.; Shoghy, K.; Ibrahim, S.F.; et al. N-Acetylcysteine Alleviated the Deltamethrin-Induced Oxidative Cascade and Apoptosis in Liver and Kidney Tissues. Int. J. Environ. Res. Public Health 2022, 19, 638. [Google Scholar] [CrossRef]
- Abdelnaby, A.; Abdel-Aleem, N.; Mansour, A.; Abdelkader, A.; Ibrahim, A.N.; Sorour, S.M.; Elgendy, E.; Bayoumi, H.; Abdelrahman, S.M.; Ibrahim, S.F.; et al. The Combination of Tamarindus indica and Coenzyme Q10 Can Be a Potential Therapy Preference to Attenuate Cadmium-Induced Hepatorenal Injury. Front. Pharmacol. 2022, 13, 2716. [Google Scholar] [CrossRef]
- Abdeen, A.; Samir, A.; Elkomy, A.; Aboubaker, M.; Habotta, O.A.; Gaber, A.; Alsanie, W.F.; Abdullah, O.; Elnoury, H.A.; Baioumy, B.; et al. The Potential Antioxidant Bioactivity of Date Palm Fruit against Gentamicin-Mediated Hepato-Renal Injury in Male Albino Rats. Biomed. Pharmacother. 2021, 143, 112154. [Google Scholar] [CrossRef]
- Sakamoto, M.I.; Murakami, A.E.; Fernandes, A.M.; Ospina-Rojas, I.C.; Nunes, K.C.; Hirata, A.K. Performance and Serum Biochemical Profile of Japanese Quail Supplemented with Silymarin and Contaminated with Aflatoxin B1. Poult. Sci. 2018, 97, 159–166. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, S.; Zhao, C.; Zhou, Y.; Zhang, L.; Liu, H.; Zhou, P.; Li, S.; Fu, L.; Zheng, Z.; et al. Genome-Scale CRISPR Knockout Screening Identifies BACH1 as a Key Regulator of Aflatoxin B1-Induced Oxidative Damage. Antioxidants 2022, 11, 1787. [Google Scholar] [CrossRef]
- Mahmoud, E.A.; El-Sayed, B.M.; Mahsoub, Y.H.; Neamat-Allah, A.N.F. Effect of Chlorella vulgaris Enriched Diet on Growth Performance, Hemato-Immunological Responses, Antioxidant and Transcriptomics Profile Disorders Caused by Deltamethrin Toxicity in Nile Tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2020, 102, 422–429. [Google Scholar] [CrossRef]
- Zahran, E.; Awadin, W.; Risha, E.; Khaled, A.A.; Wang, T. Dietary Supplementation of Chlorella vulgaris Ameliorates Chronic Sodium Arsenite Toxicity in Nile Tilapia Oreochromis Niloticus as Revealed by Histopathological, Biochemical and Immune Gene Expression Analysis. Fish. Sci. 2019, 85, 199–215. [Google Scholar] [CrossRef]
- Jahanian, E.; Mahdavi, A.H.; Asgary, S.; Jahanian, R.; Tajadini, M.H. Effect of Dietary Supplementation of Mannanoligosaccharides on Hepatic Gene Expressions and Humoral and Cellular Immune Responses in Aflatoxin-Contaminated Broiler Chicks. Prev. Vet. Med. 2019, 168, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, Q.-G.; Zhao, L.-H.; Wei, H.; Duan, G.-X.; Zhang, J.-Y.; Ji, C. Effects of Lipoic Acid on Immune Function, the Antioxidant Defense System, and Inflammation-Related Genes Expression of Broiler Chickens Fed Aflatoxin Contaminated Diets. Int. J. Mol. Sci. 2014, 15, 5649–5662. [Google Scholar] [CrossRef] [PubMed]
- Soontornchaiboon, W.; Joo, S.S.; Kim, S.M. Anti-Inflammatory Effects of Violaxanthin Isolated from Microalga Chlorella ellipsoidea in RAW 264.7 Macrophages. Biol. Pharm. Bull. 2012, 35, 1137–1144. [Google Scholar] [CrossRef]
- Lin, L.; Fu, P.; Chen, N.; Gao, N.; Cao, Q.; Yue, K.; Xu, T.; Zhang, C.; Zhang, C.; Liu, F. Total Flavonoids of Rhizoma drynariae Protect Hepatocytes against Aflatoxin B1-Induced Oxidative Stress and Apoptosis in Broiler Chickens. Ecotoxicol. Environ. Saf. 2022, 230, 113148. [Google Scholar] [CrossRef]
- Amritha, N.; Revathi, K.; Babu, M. Effect Of Green Algae (Chlorella vulgaris) On The Production Performance Of Japanese Quails (Coturnix coturnix japonica). In Proceedings of the Recent Advances in Life Sciences, Ethiraj College for Women, Chennai, India, 11–13 February 2016; p. 59. [Google Scholar]
- Halle, I.; Janczyk, P.; Freyer, G.; Souffrant, W.B. Effect of Microalgae Chlorella vulgaris on Laying Hen Performance. Arch. Zootech. 2009, 12, 5–13. [Google Scholar]
- Sikiru, A.B.; Arangasamy, A.; Alemede, I.C.; Guvvala, P.R.; Egena, S.S.A.; Ippala, J.R.; Bhatta, R. Chlorella vulgaris Supplementation Effects on Performances, Oxidative Stress and Antioxidant Genes Expression in Liver and Ovaries of New Zealand White Rabbits. Heliyon 2019, 5, e02470. [Google Scholar] [CrossRef]
- Khani, M.; Soltani, M.; Shamsaie Mehrjan, M.; Foroudi, F.; Ghaeni, M. The Effects of Chlorella vulgaris Supplementation on Growth Performance, Blood Characteristics, and Digestive Enzymes in Koi (Cyprinus carpio). Iran. J. Fish. Sci. 2017, 16, 832–843. [Google Scholar]
- Lamminen, M.; Halmemies-Beauchet-Filleau, A.; Kokkonen, T.; Jaakkola, S.; Vanhatalo, A. Different Microalgae Species as a Substitutive Protein Feed for Soya Bean Meal in Grass Silage Based Dairy Cow Diets. Anim. Feed Sci. Technol. 2019, 247, 112–126. [Google Scholar] [CrossRef]
- El-Bahr, S.; Shousha, S.; Shehab, A.; Khattab, W.; Ahmed-Farid, O.; Sabike, I.; El-Garhy, O.; Albokhadaim, I.; Albosadah, K. Effect of Dietary Microalgae on Growth Performance, Profiles of Amino and Fatty Acids, Antioxidant Status, and Meat Quality of Broiler Chickens. Animals 2020, 10, 761. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.S.; Elsabagh, R.; Allam, A.; Youssef, G.; Fadl, S.E.; Abdelhiee, E.Y.; Alkafafy, M.; Soliman, A.; Aboubakr, M. Bioremediation Role of Spirulina platensis against Deltamethrin-Mediated Toxicity and Its Chemical Residues in Chicken Meat. Environ. Sci. Pollut. Res. 2021, 28, 56188–56198. [Google Scholar] [CrossRef] [PubMed]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Microalgae as Feed Ingredients for Livestock Production and Meat Quality: A Review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Suzuki, K.; Okada, H.; Itoh, T.; Tada, T.; Mase, M.; Nakamura, K.; Kubo, M.; Tsukamoto, K. Association of Increased Pathogenicity of Asian H5N1 Highly Pathogenic Avian Influenza Viruses in Chickens with Highly Efficient Viral Replication Accompanied by Early Destruction of Innate Immune Responses. J. Virol. 2009, 83, 7475–7486. [Google Scholar] [CrossRef]
- Chen, H.; Yan, F.F.; Hu, J.Y.; Wu, Y.; Tucker, C.M.; Green, A.R.; Cheng, H.W. Immune Response of Laying Hens Exposed to 30 Ppm Ammonia for 25 Weeks. Int. J. Poult. Sci. 2017, 16, 139–146. [Google Scholar] [CrossRef]
- Samy, A.A.; El-Enbaawy, M.I.; El-Sanousi, A.A.; Abd El-Wanes, S.A.; Ammar, A.M.; Hikono, H.; Saito, T. In-Vitro Assessment of Differential Cytokine Gene Expression in Response to Infections with Egyptian Classic and Variant Strains of Highly Pathogenic H5N1 Avian Influenza Virus. Int. J. Vet. Sci. Med. 2015, 3, 1–8. [Google Scholar] [CrossRef]
- Anderson, S. Determination of Fat, Moisture, and Protein in Meat and Meat Products by Using the FOSS FoodScan Near-Infrared Spectrophotometer with FOSS Artificial Neural Network Calibration Model and Associated Database: Collaborative Study. J. AOAC Int. 2007, 90, 1073–1083. [Google Scholar] [CrossRef]
- El-medany, S.A.; El-Reffaei, W.H.M. Evaluation Canola Meal on Growing Rabbits; Nutritionally and on Their Nutritional Meat Quality. J. Food Nutr. Res. 2015, 3, 220–234. [Google Scholar] [CrossRef]
- Abdel-Monem, M.; Ragab, M.M.; Ali, S.H.; Salah, N.; Hassan, M.H.; Amro, F.H. Detection of Aflatoxins in Meat by Modified HPLC Method. Egypt. J. Chem. Environ. Health 2015, 1, 945–954. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Experimental Groups | |||
|---|---|---|---|---|
| Control | CLV | AF | AF+CLV | |
| BW (g) | 265.7 ± 27.5 | 286.7 ± 34.3 | 218.0 ± 16.5 * | 257.7 ± 25.5 # |
| BWG (g) | 15.67 ± 2.3 | 36.67 ± 6.8 | (-)32 ± 16 * | 7.67 ± 4.2 # |
| FCR | 3.82 ± 0.04 | 3.18 ± 0.19 * | 5.15 ± 0.13 * | 4.54 ± 0.31 * |
| Mortality % | 0 | 0 | 26.67% | 3.33% |
| Survival rate (%) | 100% | 100% | 73.33% | 96.67% |
| Parameters | Experimental Groups | |||
|---|---|---|---|---|
| Control | CLV | AF | AF+CLV | |
| Nutritive value | ||||
| Protein% | 22.76 ± 0.56 | 24.94 ± 0.67 * | 19.54 ± 0.47 * | 22.19 ± 0.48 # |
| Fat% | 0.57 ± 0.01 | 0.53 ± 0.01 | 1.07 ± 0.03 * | 0.74 ± 0.02 *# |
| Cholesterol (mg/g) | 20.66 ± 0.51 | 19.44 ± 0.52 | 27.74 ± 0.66 * | 24.65 ± 0.53 *# |
| Triacylglycerols (mg/100 g) | 58.75 ± 1.45 | 56.79 ± 1.52 | 63.56 ± 1.52 * | 60.77 ± 1.31 |
| AF residue (ppb) | ||||
| Liver | ND | ND | 121.54± 2.60 * | 43.32 ± 0.81 *# |
| Meat | ND | ND | 76.98± 1.65 * | 28.21 ± 0.53 *# |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elbasuni, S.S.; Ibrahim, S.S.; Elsabagh, R.; Nada, M.O.; Elshemy, M.A.; Ismail, A.K.; Mansour, H.M.; Ghamry, H.I.; Ibrahim, S.F.; Alsaati, I.; et al. The Preferential Therapeutic Potential of Chlorella vulgaris against Aflatoxin-Induced Hepatic Injury in Quail. Toxins 2022, 14, 843. https://doi.org/10.3390/toxins14120843
Elbasuni SS, Ibrahim SS, Elsabagh R, Nada MO, Elshemy MA, Ismail AK, Mansour HM, Ghamry HI, Ibrahim SF, Alsaati I, et al. The Preferential Therapeutic Potential of Chlorella vulgaris against Aflatoxin-Induced Hepatic Injury in Quail. Toxins. 2022; 14(12):843. https://doi.org/10.3390/toxins14120843
Chicago/Turabian StyleElbasuni, Sawsan S., Samar S. Ibrahim, Rasha Elsabagh, Mai O. Nada, Mona A. Elshemy, Ayman K. Ismail, Heba M. Mansour, Heba I. Ghamry, Samah F. Ibrahim, Ilhaam Alsaati, and et al. 2022. "The Preferential Therapeutic Potential of Chlorella vulgaris against Aflatoxin-Induced Hepatic Injury in Quail" Toxins 14, no. 12: 843. https://doi.org/10.3390/toxins14120843
APA StyleElbasuni, S. S., Ibrahim, S. S., Elsabagh, R., Nada, M. O., Elshemy, M. A., Ismail, A. K., Mansour, H. M., Ghamry, H. I., Ibrahim, S. F., Alsaati, I., Abdeen, A., & Said, A. M. (2022). The Preferential Therapeutic Potential of Chlorella vulgaris against Aflatoxin-Induced Hepatic Injury in Quail. Toxins, 14(12), 843. https://doi.org/10.3390/toxins14120843