Targeted Proteomics Analysis of Staphylococcal Superantigenic Toxins in Menstrual Fluid from Women with Menstrual Toxic Shock Syndrome (mTSS)

 , , and
, , and
Abstract
1. Introduction
2. Results
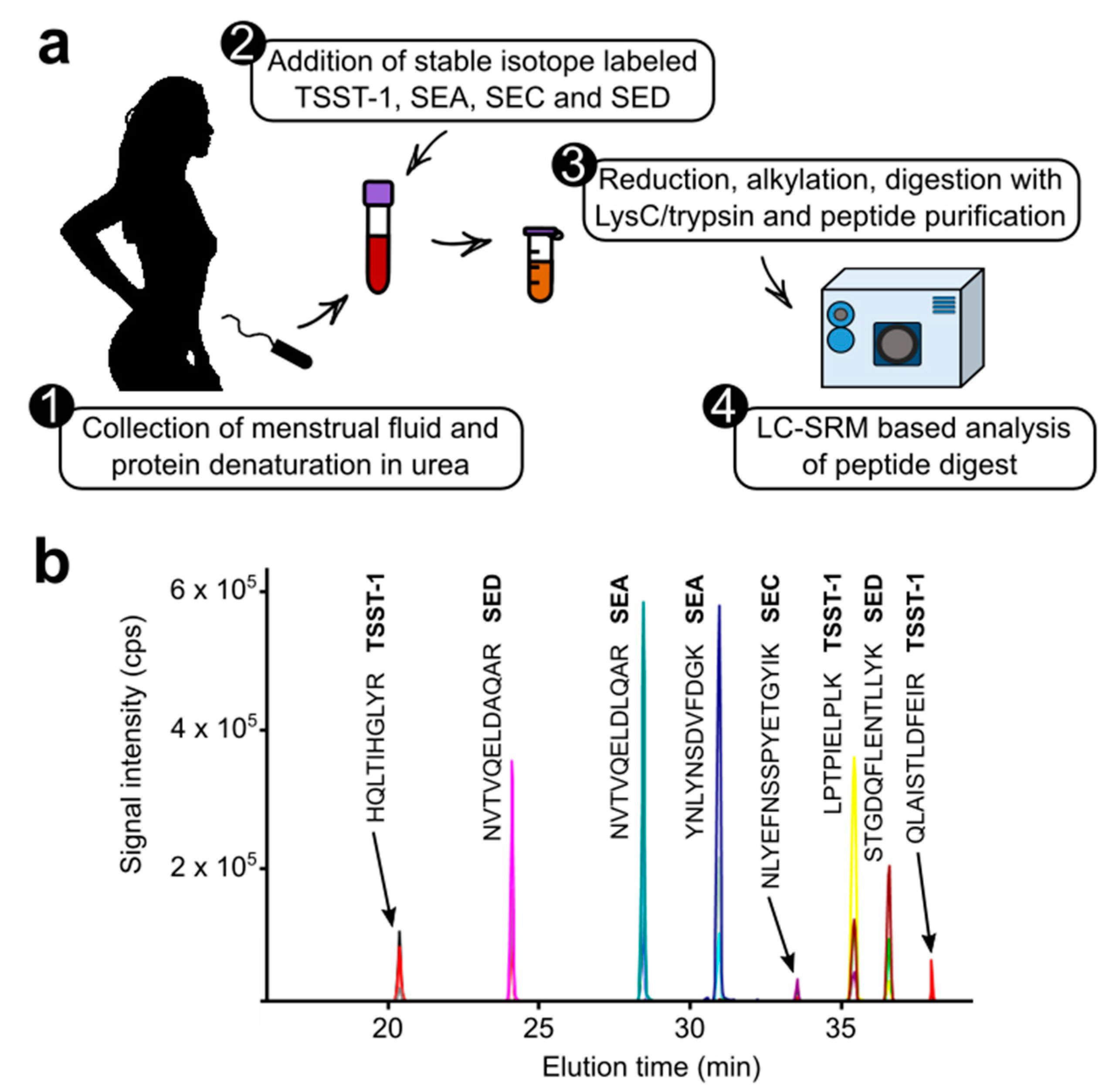
2.1. Development of the Targeted Proteomic Assay
2.2. Investigation of Clinical Samples
3. Discussion
4. Materials and Methods
4.1. Ethical Review of the Study
4.2. Subjects
4.3. Sample Collection and Microbiological Characterization
4.4. Production of Full-Length Stable Isotope-Labeled Toxins
4.5. Biochemical Preparation of Menstrual Fluids
4.6. LC-SRM Analysis
4.7. LC-SRM Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schlievert, P.M.; Davis, C.C. Device-Associated Menstrual Toxic Shock Syndrome. Clin. Microbiol. Rev. 2020, 33, e00032-19. [Google Scholar] [CrossRef] [PubMed]
- Billon, A.; Gustin, M.P.; Tristan, A.; Benet, T.; Berthiller, J.; Gustave, C.A.; Vanhems, P.; Lina, G. Association of characteristics of tampon use with menstrual toxic shock syndrome in France. EClinicalMedicine 2020, 21, 100308. [Google Scholar] [CrossRef] [PubMed]
- Parsonnet, J.; Hansmann, M.A.; Delaney, M.L.; Modern, P.A.; Dubois, A.M.; Wieland-Alter, W.; Wissemann, K.W.; Wild, J.E.; Jones, M.B.; Seymour, J.L.; et al. Prevalence of toxic shock syndrome toxin 1-producing Staphylococcus aureus and the presence of antibodies to this superantigen in menstruating women. J. Clin. Microbiol. 2005, 43, 4628–4634. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, A.R.; Salgado-Pabon, W.; Kohler, P.L.; Horswill, A.R.; Leung, D.Y.; Schlievert, P.M. Staphylococcal and streptococcal superantigen exotoxins. Clin. Microbiol. Rev. 2013, 26, 422–447. [Google Scholar] [CrossRef]
- Thomas, D.; Dauwalder, O.; Brun, V.; Badiou, C.; Ferry, T.; Etienne, J.; Vandenesch, F.; Lina, G. Staphylococcus aureus superantigens elicit redundant and extensive human Vbeta patterns. Infect. Immun. 2009, 77, 2043–2050. [Google Scholar] [CrossRef]
- Ferry, T.; Thomas, D.; Perpoint, T.; Lina, G.; Monneret, G.; Mohammedi, I.; Chidiac, C.; Peyramond, D.; Vandenesch, F.; Etienne, J. Analysis of superantigenic toxin Vbeta T-cell signatures produced during cases of staphylococcal toxic shock syndrome and septic shock. Clin. Microbiol. Infect. 2008, 14, 546–554. [Google Scholar] [CrossRef]
- Low, D.E. Toxic shock syndrome: Major advances in pathogenesis, but not treatment. Crit. Care Clin. 2013, 29, 651–675. [Google Scholar] [CrossRef]
- Lappin, E.; Ferguson, A.J. Gram-positive toxic shock syndromes. Lancet Infect. Dis. 2009, 9, 281–290. [Google Scholar] [CrossRef]
- Chiaruzzi, M.; Barbry, A.; Muggeo, A.; Tristan, A.; Jacquemond, I.; Badiou, C.; Cluzeau, L.; Bourdeau, S.; Durand, T.; Engelmann, A.; et al. Vaginal Tampon Colonization by Staphylococcus aureus in Healthy Women. Appl. Environ. Microbiol. 2020, 86, e01249-20. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, B.; Prinz, M.; Siegel, D. Proteomic analysis of menstrual blood. Mol. Cell Proteom. 2012, 11, 1024–1035. [Google Scholar] [CrossRef]
- Farrah, T.; Deutsch, E.W.; Omenn, G.S.; Campbell, D.S.; Sun, Z.; Bletz, J.A.; Mallick, P.; Katz, J.E.; Malmstrom, J.; Ossola, R.; et al. A high-confidence human plasma proteome reference set with estimated concentrations in PeptideAtlas. Mol. Cell Proteom. 2011, 10, M110 006353. [Google Scholar] [CrossRef]
- Bryk, A.H.; Wisniewski, J.R. Quantitative Analysis of Human Red Blood Cell Proteome. J. Proteome Res. 2017, 16, 2752–2761. [Google Scholar] [CrossRef] [PubMed]
- Vaswani, K.; Ashman, K.; Reed, S.; Salomon, C.; Sarker, S.; Arraztoa, J.A.; Perez-Sepulveda, A.; Illanes, S.E.; Kvaskoff, D.; Mitchell, M.D.; et al. Applying SWATH Mass Spectrometry to Investigate Human Cervicovaginal Fluid During the Menstrual Cycle. Biol. Reprod. 2015, 93, 39. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Notermans, S.; Timmermans, P.; Nagel, J. Interaction of staphylococcal protein A in enzyme-linked immunosorbent assays for detecting staphylococcal antigens. J. Immunol. Methods 1982, 55, 35–41. [Google Scholar] [CrossRef]
- Rosten, P.M.; Bartlett, K.H.; Chow, A.W. Detection and quantitation of toxic shock syndrome toxin 1 in vitro and in vivo by noncompetitive enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1987, 25, 327–332. [Google Scholar] [CrossRef]
- Brun, V.; Masselon, C.; Garin, J.; Dupuis, A. Isotope dilution strategies for absolute quantitative proteomics. J. Proteom. 2009, 72, 740–749. [Google Scholar] [CrossRef]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef]
- Brun, V.; Dupuis, A.; Adrait, A.; Marcellin, M.; Thomas, D.; Court, M.; Vandenesch, F.; Garin, J. Isotope-labeled protein standards: Toward absolute quantitative proteomics. Mol. Cell Proteom. 2007, 6, 2139–2149. [Google Scholar] [CrossRef]
- Wisniewski, J.R.; Mann, M. Consecutive proteolytic digestion in an enzyme reactor increases depth of proteomic and phosphoproteomic analysis. Anal. Chem. 2012, 84, 2631–2637. [Google Scholar] [CrossRef]
- Jacquemond, I.; Muggeo, A.; Lamblin, G.; Tristan, A.; Gillet, Y.; Bolze, P.A.; Bes, M.; Gustave, C.A.; Rasigade, J.P.; Golfier, F.; et al. Complex ecological interactions of Staphylococcus aureus in tampons during menstruation. Sci. Rep. 2018, 8, 9942. [Google Scholar] [CrossRef] [PubMed]
- Warren, L.A.; Shih, A.; Renteira, S.M.; Seckin, T.; Blau, B.; Simpfendorfer, K.; Lee, A.; Metz, C.N.; Gregersen, P.K. Analysis of menstrual effluent: Diagnostic potential for endometriosis. Mol. Med. 2018, 24, 1. [Google Scholar] [CrossRef] [PubMed]
- Adrait, A.; Lebert, D.; Trauchessec, M.; Dupuis, A.; Louwagie, M.; Masselon, C.; Jaquinod, M.; Chevalier, B.; Vandenesch, F.; Garin, J.; et al. Development of a Protein Standard Absolute Quantification (PSAQ) assay for the quantification of Staphylococcus aureus enterotoxin A in serum. J. Proteom. 2012, 75, 3041–3049. [Google Scholar] [CrossRef] [PubMed]
- Gilquin, B.; Jaquinod, M.; Louwagie, M.; Kieffer-Jaquinod, S.; Kraut, A.; Ferro, M.; Becher, F.; Brun, V. A proteomics assay to detect eight CBRN-relevant toxins in food. Proteomics 2017, 17, 1600357. [Google Scholar] [CrossRef] [PubMed]
- Schlievert, P.M.; Nemeth, K.A.; Davis, C.C.; Peterson, M.L.; Jones, B.E. Staphylococcus aureus exotoxins are present in vivo in tampons. Clin. Vaccine Immunol. 2010, 17, 722–727. [Google Scholar] [CrossRef]
- Vojtov, N.; Ross, H.F.; Novick, R.P. Global repression of exotoxin synthesis by staphylococcal superantigens. Proc. Natl. Acad. Sci. USA 2002, 99, 10102–10107. [Google Scholar] [CrossRef]
- Patot, S.; Imbert, P.R.; Baude, J.; Martins Simoes, P.; Campergue, J.B.; Louche, A.; Nijland, R.; Bes, M.; Tristan, A.; Laurent, F.; et al. The TIR Homologue Lies near Resistance Genes in Staphylococcus aureus, Coupling Modulation of Virulence and Antimicrobial Susceptibility. PLoS Pathog. 2017, 13, e1006092. [Google Scholar] [CrossRef]
- Lebert, D.; Dupuis, A.; Garin, J.; Bruley, C.; Brun, V. Production and use of stable isotope-labeled proteins for absolute quantitative proteomics. Methods Mol. Biol. 2011, 753, 93–115. [Google Scholar] [CrossRef]
- Louwagie, M.; Kieffer-Jaquinod, S.; Dupierris, V.; Coute, Y.; Bruley, C.; Garin, J.; Dupuis, A.; Jaquinod, M.; Brun, V. Introducing AAA-MS, a rapid and sensitive method for amino acid analysis using isotope dilution and high-resolution mass spectrometry. J. Proteome Res. 2012, 11, 3929–3936. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Subject Number | mTSS 1 | Superantigenic Toxin Genes Present 2 | LC-SRM Detection of TSST-1 3 | Detection of SEA, SEC, SED 3 |
|---|---|---|---|---|
| 1 | Yes | tst, sea | 460 ng/mL | Not detected |
| 2 | Yes | tst, sea | Not detected | Not detected |
| 3 | Yes | tst, sea | 80 ng/mL | Not detected |
| 4 | Yes | tst, sec, sed | 40 ng/mL | SEC detected but not quantifiable |
| 5 | Yes | tst, sea | 70 ng/mL | Not detected |
| 6 | Yes | tst | 10 ng/mL | Not detected |
| 7 | No | tst, sed | Not detected | Not detected |
| 8 | No | tst | Not detected | Not detected |
| 9 | No | tst, sea | Not detected | Not detected |
| 10 | No | tst | Not detected | Not detected |
| 11 | No | tst, sea | Not detected | Not detected |
| 12 | No | tst, sea | Not detected | Not detected |
| 13 | No | tst | Not detected | Not detected |
| 14 | No | tst | Not detected | Not detected |
| 15 | No | tst, sea | Not detected | Not detected |
| 16 | No | tst, sea | Not detected | Not detected |
| 17 | No | tst, sea | Not detected | Not detected |
| 18 | No | tst | Not detected | Not detected |
| 19 | No | tst, sec, sed | Not determined (signal contamination 4) | SEC 1.07 µg/mL |
| 20 | No | tst, sea | 330 ng/mL | Not detected |
| 21 | No | tst | Not detected | Not detected |
| 22 | No | tst, sea | Not detected | Not detected |
| 23 | No | sea, sed | Not detected | Not detected |
| 24 | No | sea, sed | Not detected | Not detected |
| 25 | No | sec | Not detected | Not detected |
| 26 | No | sec | Not detected | Not detected |
| 27 | No | sea | Not detected | Not detected |
| 28 | No | sea | Not detected | Not detected |
| S. aureus Toxin | UniProt Reference | Signature Peptide 1 | Fragment Ion | SRM Transitions 2 | Collision Energy (eV) | |
|---|---|---|---|---|---|---|
| Q1 m/z | Q3 m/z | |||||
| TSST-1 | P06886 | LPTPIELPLK | +2y7 | 560.9 | 809.5 | 29.8 |
| +2y9 + 2 | 560.9 | 504.3 | 29.8 | |||
| +2y8 | 560.9 | 910.6 | 29.8 | |||
| LPTPIELPL [13C6,15N2] K | +2y7 | 564.9 | 817.5 | 29.8 | ||
| +2y9 + 2 | 564.9 | 508.3 | 29.8 | |||
| +2y8 | 564.9 | 918.6 | 29.8 | |||
| QLAISTLDFEIR | +3y5 | 469.3 | 679.3 | 25.7 | ||
| +3y4 | 469.3 | 564.3 | 25.7 | |||
| +3y3 | 469.3 | 417.2 | 25.7 | |||
| QLAISTLDFEI [13C6,15N4] R | +3y5 | 472.6 | 689.3 | 25.7 | ||
| +3y4 | 472.6 | 574.3 | 25.7 | |||
| +3y3 | 472.6 | 427.2 | 25.7 | |||
| HQLTQIHGLYR | +3y4 | 455.9 | 508.3 | 25.2 | ||
| +3y5 | 455.9 | 645.4 | 25.2 | |||
| +3y6 | 455.9 | 758.4 | 25.2 | |||
| HQLTQIHGLY [13C6,15N4] R | +3y4 | 459.3 | 518.3 | 25.2 | ||
| +3y5 | 459.3 | 655.4 | 25.2 | |||
| +3y6 | 459.3 | 768.4 | 25.2 | |||
| SEA | NVTVQELDLQAR | +2y8 | 693.4 | 972.5 | 33.8 | |
| +2y7 | 693.4 | 844.5 | 33.8 | |||
| +2y6 | 693.4 | 715.4 | 33.8 | |||
| NVTVQELDLQA [13C6,15N4] R | +2y8 | 698.4 | 982.5 | 33.8 | ||
| +2y7 | 698.4 | 854.5 | 33.8 | |||
| +2y6 | 698.4 | 725.4 | 33.8 | |||
| YNLYNSDVFDGK | +2y8 | 717.8 | 881.4 | 34.7 | ||
| +2y7 | 717.8 | 767.4 | 34.7 | |||
| +2y6 | 717.8 | 680.3 | 34.7 | |||
| YNLYNSDVFDG [13C6,15N2] K | +2y8 | 721.8 | 889.4 | 34.7 | ||
| +2y7 | 721.8 | 775.4 | 34.7 | |||
| +2y6 | 721.8 | 688.3 | 34.7 | |||
| SEC | NLYEFNSSPYETGYIK | +2y8 | 963.0 | 970.5 | 43.5 | |
| +2y5 | 963.0 | 581.3 | 43.5 | |||
| +2y8 + 2 | 963.0 | 485.7 | 43.5 | |||
| NLYEFNSSPYETGYI [13C6] K | +2y8 | 966.0 | 976.5 | 43.5 | ||
| +2y5 | 966.0 | 587.3 | 43.5 | |||
| +2y8 + 2 | 966.0 | 488.8 | 43.5 | |||
| SED | STGDQFLENTLLYK | +2y7 | 814.9 | 880.5 | 38.2 | |
| +2y6 | 814.9 | 751.4 | 38.2 | |||
| +2y5 | 814.9 | 637.4 | 38.2 | |||
| STGDQFLENTLLY [13C6] K | +2y7 | 817.9 | 886.5 | 38.2 | ||
| +2y6 | 817.9 | 757.4 | 38.2 | |||
| +2y5 | 817.9 | 643.4 | 38.2 | |||
| NVTVQELDAQAR | +2y8 | 672.4 | 930.5 | 33.0 | ||
| +2y7 | 672.4 | 802.4 | 33.0 | |||
| +2y5 | 672.4 | 560.3 | 33.0 | |||
| NVTVQELDAQA [13C6] R | +2y8 | 675.4 | 936.5 | 33.0 | ||
| +2y7 | 675.4 | 808.4 | 33.0 | |||
| +2y5 | 675.4 | 566.3 | 33.0 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Courçon, M.; Badiou, C.; Louwagie, M.; Etievant, S.; Jaquinod, M.; Lina, G.; Brun, V. Targeted Proteomics Analysis of Staphylococcal Superantigenic Toxins in Menstrual Fluid from Women with Menstrual Toxic Shock Syndrome (mTSS). Toxins 2022, 14, 886. https://doi.org/10.3390/toxins14120886
Courçon M, Badiou C, Louwagie M, Etievant S, Jaquinod M, Lina G, Brun V. Targeted Proteomics Analysis of Staphylococcal Superantigenic Toxins in Menstrual Fluid from Women with Menstrual Toxic Shock Syndrome (mTSS). Toxins. 2022; 14(12):886. https://doi.org/10.3390/toxins14120886
Chicago/Turabian StyleCourçon, Marie, Cédric Badiou, Mathilde Louwagie, Sibyle Etievant, Michel Jaquinod, Gérard Lina, and Virginie Brun. 2022. "Targeted Proteomics Analysis of Staphylococcal Superantigenic Toxins in Menstrual Fluid from Women with Menstrual Toxic Shock Syndrome (mTSS)" Toxins 14, no. 12: 886. https://doi.org/10.3390/toxins14120886
APA StyleCourçon, M., Badiou, C., Louwagie, M., Etievant, S., Jaquinod, M., Lina, G., & Brun, V. (2022). Targeted Proteomics Analysis of Staphylococcal Superantigenic Toxins in Menstrual Fluid from Women with Menstrual Toxic Shock Syndrome (mTSS). Toxins, 14(12), 886. https://doi.org/10.3390/toxins14120886