Saporin as a Commercial Reagent: Its Uses and Unexpected Impacts in the Biological Sciences—Tools from the Plant Kingdom


 , , and
, , and
Abstract
1. Introduction
1.1. RIPs in Cancer Research
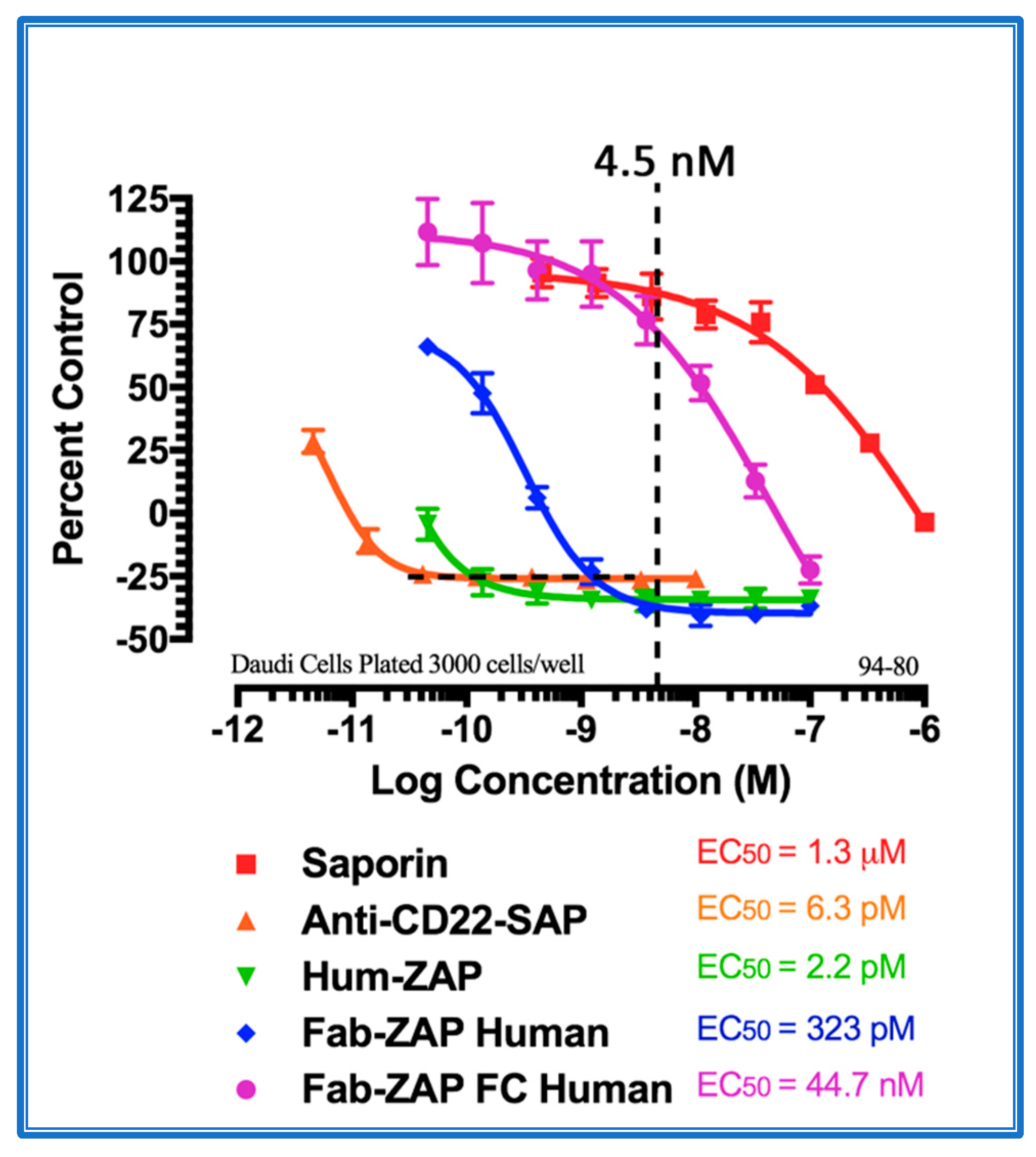
- Potency. In over 27 years of experience producing Saporin conjugates, our laboratory has seen many conjugates that work impressively well, and some that do not. The failings are most often caused by the targeting agent, so it is best to move on to test other targeting agents whether it be another antibody, a ligand, a DNA aptamer, any material that better “sees” the membrane molecule that is the target.
- Non-target activity in vivo. If this occurs in development, it is time to drop the project.
- Controlling stoichiometry. This can be accomplished in one of two ways: DNA construction of a fusion protein, or careful determination and description of the chemistry process for the construction of the conjugation.
- Tumor penetration. The use of saponins appears to be a promising method for increase in penetration and resulting potency [27].
- Manufacturing costs. GMP manufacturing of Saporin conjugates has a high price tag with very few facilities available with the appropriate equipment to produce grams of drug.
1.2. RIPs in Neuroscience Research
- Combined apoptotic body analogues for efficient targeted therapy [41]
- Targeted and intracellular delivery of protein therapeutics by a boronated polymer [42]
- Macropinocytosis-inducible extracellular vesicles modified with antimicrobial protein CAP18-derived cell-penetrating peptides for efficient intracellular delivery [43]
- Nanobody-targeted polymeric nanoparticles [44]
- pH-sensitive coiled-coil peptide-cross-linked hyaluronic acid nanogels [45]
- CD22 ligands on a natural N-glycan scaffold [46]
- Vectorization of biomacromolecules into cells using extracellular vesicles with enhanced internalization [47]
- Soluble T-cell receptors produced in human cells for targeted delivery [48]
2. Targeting Alzheimer’s Disease
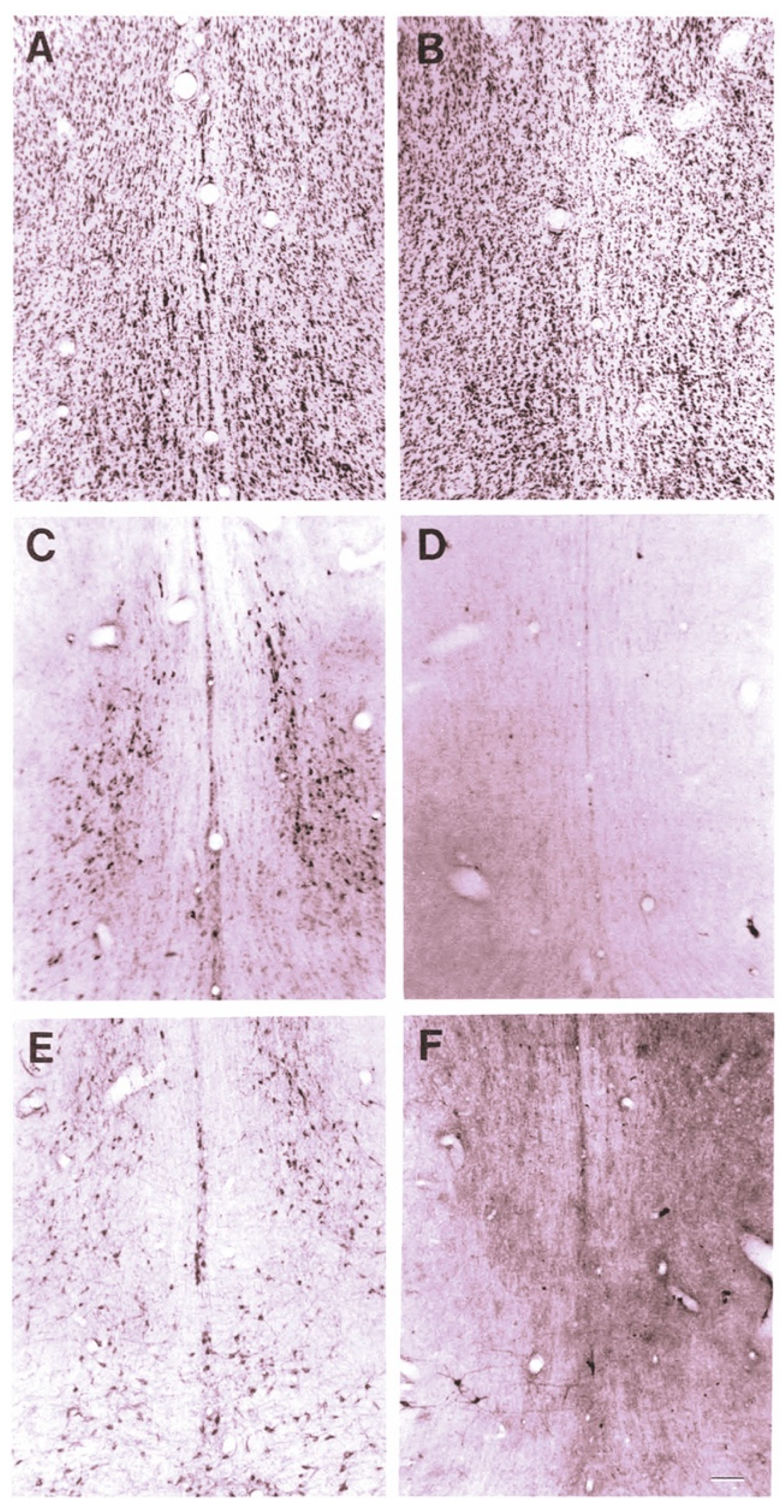
2.1. 192-IgG-SAP
2.2. 192-IgG-SAP Species Specific Alternatives
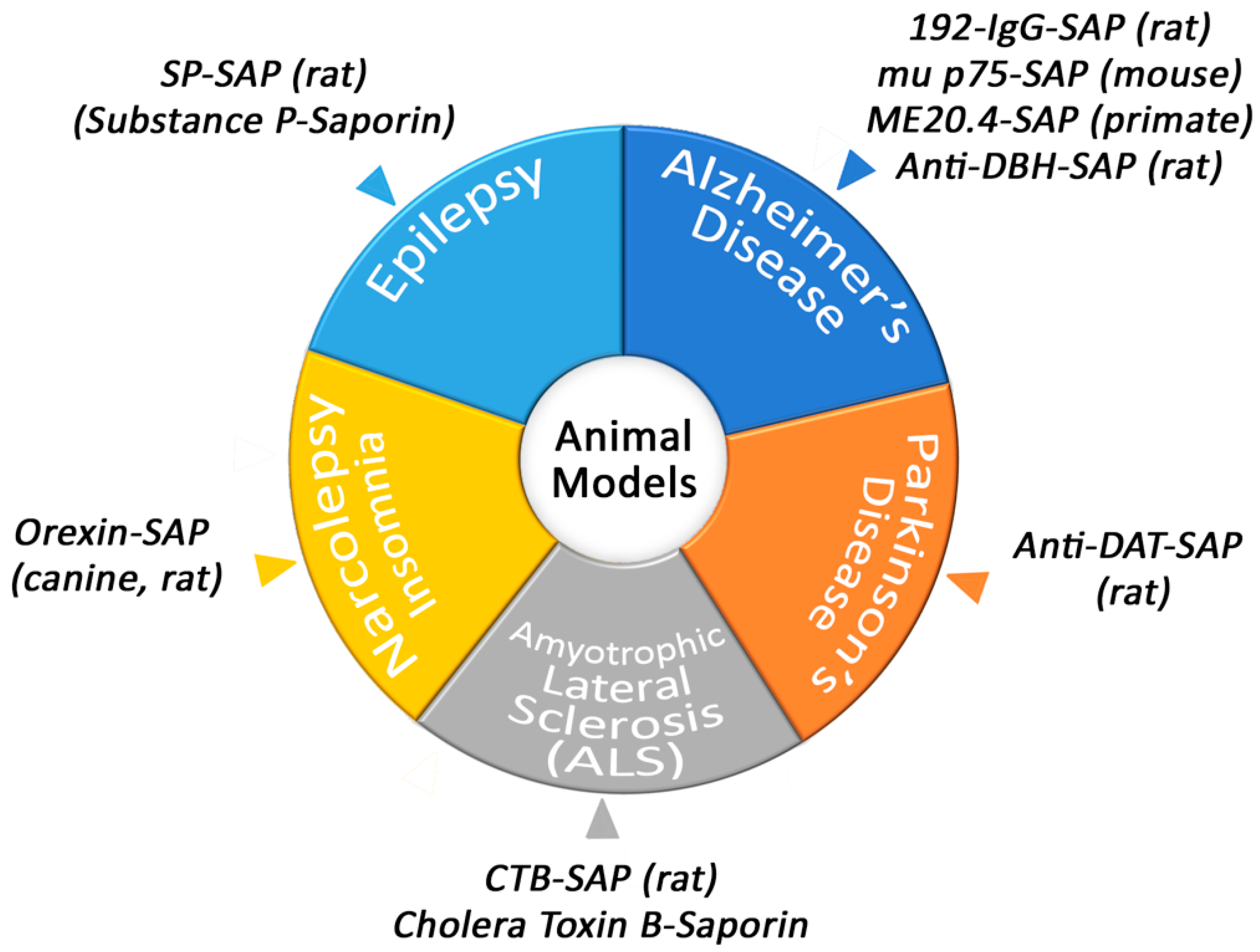
3. Behavior, Disease and Animal Models
3.1. Narcolepsy/Insomnia
3.2. Amyotrophic Lateral Sclerosis
3.3. Parkinson’s Disease
3.4. Itch
3.5. Epilepsy
3.6. Gastroenterology
- Kanai et al. investigated the role of macrophages in a 2,4,6-trinitrobenzene sulfonic acid-induced colitis mouse model [118]. A 20 μg dose of a CD11b antibody conjugated to Saporin (Mac-1-SAP) was administered parenterally in the tail vein. Seven days after treatment, mice showed no evidence of intestinal inflammation.
- In this second study by the Kanai group, Mac-1-SAP was administered as a single intravenous (IV) injection that significantly reduced the amount of intestinal inflammation [119].
- The study by Yamazaki et al. used the same colitis mouse model referenced above and investigated the role of mucosal T cells that express high levels of interleukin-7 receptor (IL-7R) in the development and treatment of chronic colitis. A custom conjugate of an antibody to the IL-7R and Saporin was administered via intraperitoneal (IP) injection once a week for 6 weeks to 20 to 24-week-old mice. Selective elimination of IL-7R-expressing T cells ameliorated established, ongoing colitis [120].
- A rat model of persistent experimental pancreatitis was used to determine the role of descending pathways in the pain caused by pancreatitis [121]. Rats received 1.5-picomolar injections of the targeted toxin Dermorphin–Saporin (a conjugate of the peptide dermorphin and Saporin; MOR-SAP) into each side of the rostral ventromedial medulla. Although the ablation of mu-opioid receptor-expressing neurons by MOR-SAP did not prevent the initial expression of pancreatitis pain, chronic pain was eliminated thereby linking the maintenance of pancreatitis pain to descending pathways. This treatment also prevented the increase of spinal dynorphin content.
- Macrophages perform different functions depending on the tissue type. The specific differentiation that macrophages undergo in response to their environment is called polarization. Criscimanna et al. used a mouse pancreatic lesion model to examine the polarization of macrophages into the two distinct states known, M1 and M2 [122]. Mice received 20 μg of Mac-1-SAP in a tail vein. The results of this study demonstrate that various aspects of macrophage polarization are required for pancreatic regeneration. The authors state that, “Additional study of these processes and signals might lead to new approaches for treating Type I diabetes or pancreatitis”.
- In a study conducted by Wang et al., diabetes was induced in mice by injection of streptozotocin (STZ). In order to investigate the role of macrophages in the development of diabetic encephalopathy, IP injections of Mac-1-SAP were administered twice weekly. Mice receiving Mac-1-SAP had greatly reduced numbers of inflammatory macrophages in the brain without affecting blood glucose, serum insulin, glucose responses or beta cell mass [123].
- CCK-SAP was used to induce neural lesioning of vagal afferent neurons while sparing vagal efferent neurons in the nodose ganglia of rats [124].
- In 2018, Suarez et al. injected CCK-SAP into the nodose ganglia to “eliminate ~80% of GI-derived vagal sensory input to the brain while leaving intact all brain-to-gut vagal motor signaling, and supradiaphragmatic and colonic vagal sensory signaling” [125]. This technique identified a previously unknown role for the gut–brain axis in memory control.
3.7. Noradrenergic Lesioning/Anti-DBH-SAP
3.8. OX7-SAP
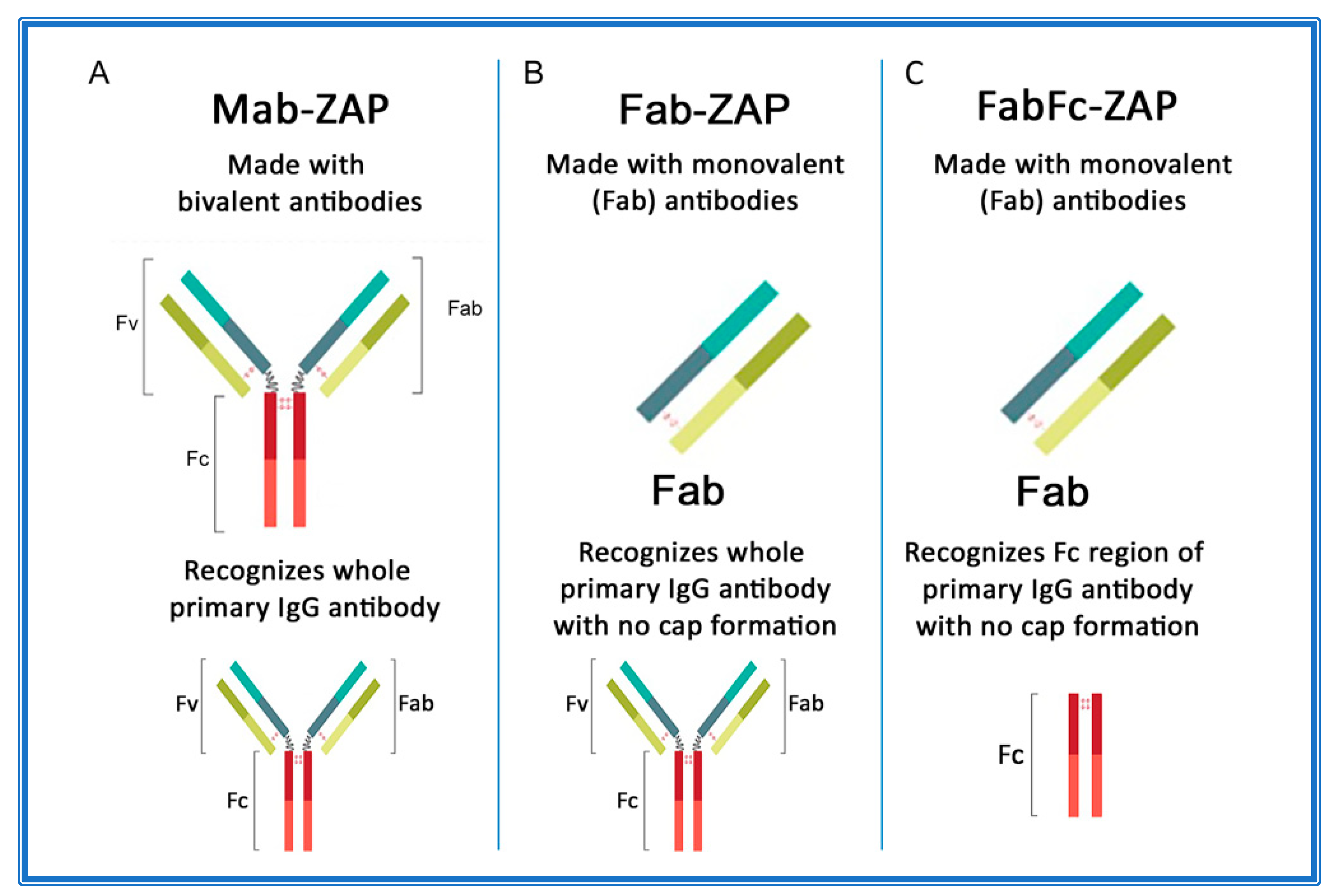
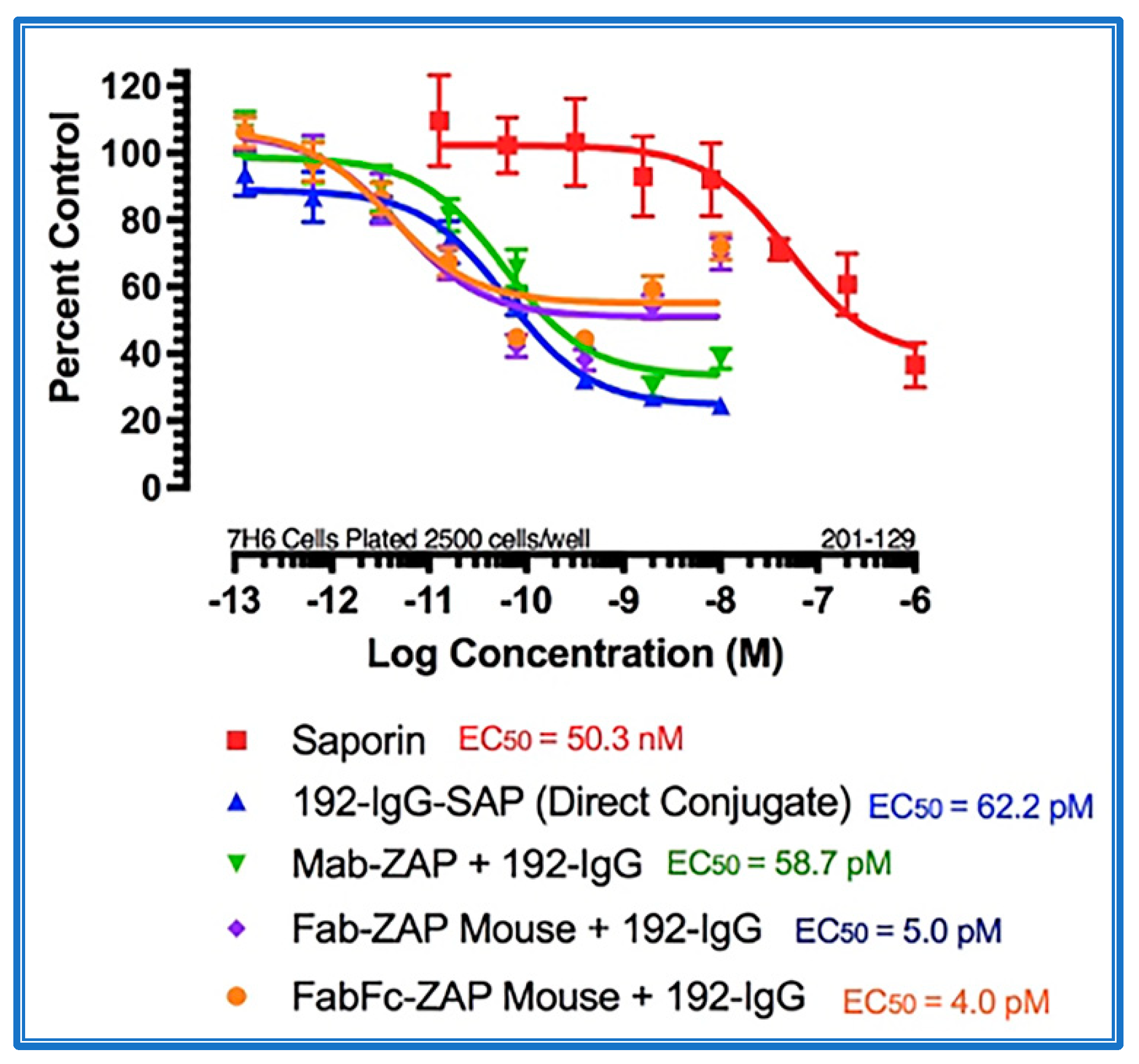
4. Secondary Conjugates and Streptavidin-ZAP
5. Clinical Trial for Cancer Pain
5.1. The Road to Human Clinical Trials
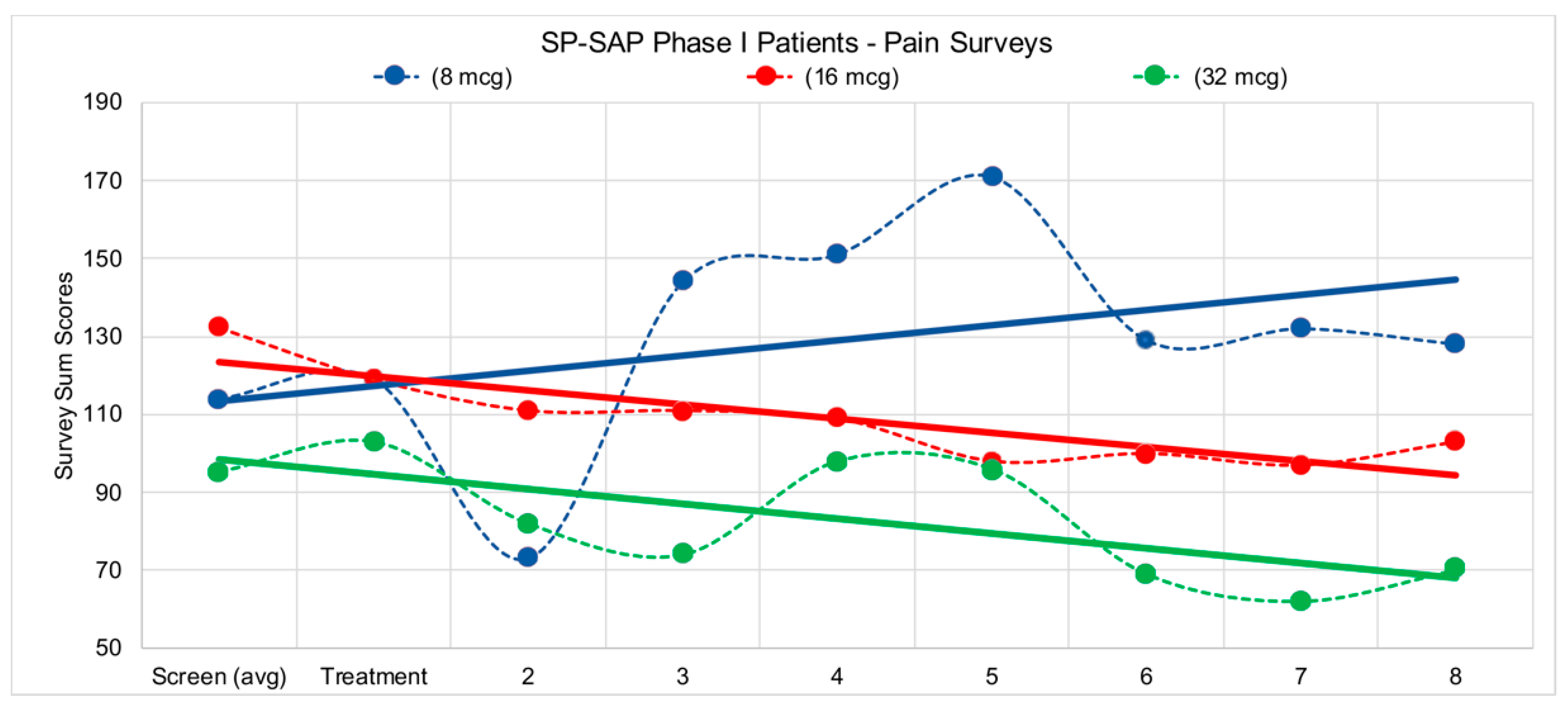
5.2. Human Clinical Trial
5.3. Veterinary Clinical Trial
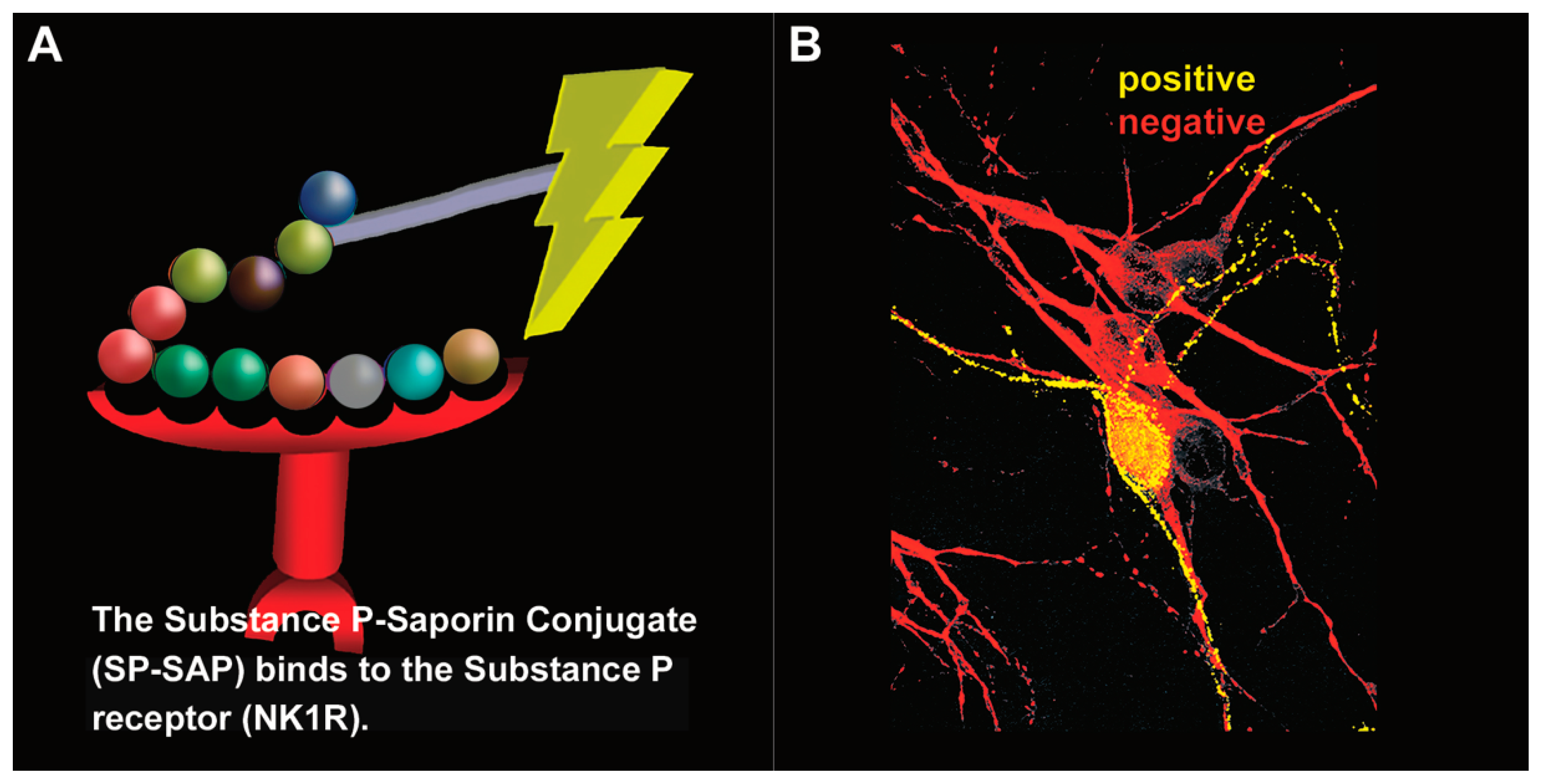
5.4. Preclinical Work Using SP-SAP
- SP-SAP in the upper cervical dorsal horn of adult rats reduced aversion to suprathreshold but not near-threshold levels of oral high intensity pain stimulant (capsaicin) [151].
- SP-SAP eliminated a pivotal component of the spinal circuits involved in triggering central sensitization and hyperalgesia [152].
- SP-SAP removed NK1R+ spinal projection neurons that project to higher brain areas and control spinal excitability—and therefore pain sensitivity—primarily through descending pathways from the brainstem. This implies central sensitization is stopped by SP-SAP [153].
- SP-SAP inhibited high level pain transmission in more complex pathologic pain models [154].
- SP-SAP attenuated the tactile and cold hypersensitivity and abnormal neuronal coding (including spontaneous activity, expansion of receptive field size) seen after spinal nerve ligation [155].
- Ablation of NK1R+ lamina I cells eliminates the ascending limb of a spinal–bulbospinal loop that engages descending facilitation [121].
- Substance P–Saporin decreased the ratio of NK1R+ neurons innervating the disc related to discogenic low back pain. Conclusion: SP-SAP may be a useful tool to investigate the mechanism of discogenic low back pain [156].
- The generation of intrinsic GABAergic transmission in the spinal cord appears dependent on NK1R+ neurons, yet despite the loss of GABAergic inhibitory controls after SP-SAP treatment, the net effect is a decrease in spinal cord excitability. Thus, activation of these cells predominantly drives facilitation (and elimination inhibits sensitization) [157].
- Since the same neuronal population drives descending facilitation and inhibition, the reduced excitability of lamina V/VI WDR (wide dynamic range) neurons seen after loss of NK1R+ neurons by SP-SAP indicates a dominant role of descending facilitation [158].
- Elimination of NK1R+ neurons with SP-SAP ends descending facilitatory pathways that function in central sensitization [159].
- In a dog safety study, data indicate no adverse toxicity at doses up to 10 times those necessary for producing loss of superficial NK1R+ neurons in a large animal model [144].
5.5. Preclinical Work Using Stable Substance P-Saporin (SSP-SAP)
6. A Closing Note—The Pandemic’s Impact on Research
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schwartz, R.S. Paul Ehrlich’s Magic Bullets. N. Engl. J. Med. 2004, 350, 1079–1080. [Google Scholar] [CrossRef]
- Stirpe, F.; Barbieri, L.; Battelli, M.G.; Soria, M.; Lappi, D. Ribosome–Inactivating Proteins from Plants: Present Status and Future Prospects. Bio/Technology 1992, 10, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Bourrie, B.J.P.; Casellas, P.; Blythman, H.E.; Jansen, F.K. Study of the plasma clearance of antibody–ricin-A-chain immunotoxins. Evidence for specific recognition sites on the A chain that mediate rapid clearance of the immunotoxin. JBIC J. Biol. Inorg. Chem. 1986, 155, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Siena, S.; Bregni, M.; Formosa, A.; Martineau, D.; Lappi, D.A.; Bonadonna, G.; Gianni, A.M. Evaluation of anti-human T-lymphocyte saporin immunotoxins potentially useful in human transplantation. Transplantation 1988, 46, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, F.; Flavell, D.J.; Angelucci, F.; Fabbrini, M.S.; Ippoliti, R. Strategies to Improve the Clinical Utility of Saporin-Based Targeted Toxins. Toxins 2018, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Onda, M.; Beers, R.; Xiang, L.; Nagata, S.; Wang, Q.-C.; Pastan, I. An immunotoxin with greatly reduced immunogenicity by identification and removal of B cell epitopes. Proc. Natl. Acad. Sci. USA 2008, 105, 11311–11316. [Google Scholar] [CrossRef]
- Zein, N.; Sinha, A.M.; McGahren, W.J.; Ellestad, G.A. Calicheamicin γ 1 I: An Antitumor Antibiotic That Cleaves Double-Stranded DNA Site Specifically. Science 1988, 240, 1198–1201. [Google Scholar] [CrossRef]
- Bolshakov, A.P.; Stepanichev, M.Y.; Dobryakova, Y.V.; Spivak, Y.S.; Markevich, V.A. Saporin from Saponaria officinalis as a Tool for Experimental Research, Modeling, and Therapy in Neuroscience. Toxins 2020, 12, 546. [Google Scholar] [CrossRef]
- Chaudhary, V.K.; FitzGerald, D.J.; Adhya, S.; Pastan, I. Activity of a recombinant fusion protein between transforming growth factor type alpha and Pseudomonas toxin. Proc. Natl. Acad. Sci. USA 1987, 84, 4538–4542. [Google Scholar] [CrossRef]
- Kreitman, R.J.; Siegall, C.B.; Chaudhary, V.K.; Fitzgerald, D.J.; Pastan, I. Properties of chimeric toxins with two recognition domains: Interleukin 6 and transforming growth factor alpha at different locations in Pseudomonas exotoxin. Bioconjug. Chem. 1992, 3, 63–68. [Google Scholar] [CrossRef]
- Kaplan, G.; Lee, F.; Onda, M.; Kolyvas, E.; Bhardwaj, G.; Baker, D.; Pastan, I. Protection of the Furin Cleavage Site in Low-Toxicity Immunotoxins Based on Pseudomonas Exotoxin A. Toxins 2016, 8, 217. [Google Scholar] [CrossRef]
- Chaudhary, V.K.; Jinno, Y.; FitzGerald, D.; Pastan, I. Pseudomonas extoxin contains a specific sequence at the carboxyl ter-minus that is required for cytotoxicity. Proc. Natl. Acad. Sci. USA 1990, 87, 308–312. [Google Scholar] [CrossRef]
- Mazor, R.; Pastan, I. Immunogenicity of Immunotoxins Containing Pseudomonas Exotoxin A: Causes, Consequences, and Mitigation. Front. Immunol. 2020, 11, 1261. [Google Scholar] [CrossRef]
- Collier, R.J. Effect of diphtheria toxin on protein synthesis: Inactivation of one of the transfer factors. J. Mol. Biol. 1967, 25, 83–98. [Google Scholar] [CrossRef]
- Stirpe, F.; Gasperi-Campani, A.; Barbieri, L.; Falasca, A.; Abbondanza, A.; Stevens, A.W. Ribosome-inactivating proteins from the seeds of Saponaria officinalis L. (soapwort), of Agrostemma githago L. (corn cockle) and of Asparagus officinalis L. (asparagus), and from the latex of Hura crepitans L. (sandbox tree). Biochem. J. 1983, 216, 617–625. [Google Scholar] [CrossRef]
- Thorpe, P.E.; Brown, A.N.F.; Bremner, J.A.G.; Foxwell, B.M.J.; Stirpe, F. An Immunotoxin Composed of Monoclonal Anti-Thy 1.1 Antibody and a Ribosome-Inactivating Protein from Saponaria officinalis: Potent Antitumor Effects In Vitro and In Vivo. JNCI J. Natl. Cancer Inst. 1985, 75, 151–159. [Google Scholar] [CrossRef]
- Siena, S.; Lappi, D.A.; Bregni, M.; Formosa, A.; Villa, S.; Soria, M.; Bonadonna, G.; Gianni, A.M. Synthesis and characterization of an antihuman T-lymphocyte saporin immunotoxin (OKT1-SAP) with in vivo stability into nonhuman primates. Blood 1988, 72, 756–765. [Google Scholar] [CrossRef]
- Glennie, M.J.; Brennand, D.M.; Bryden, F.; McBride, H.M.; Stirpe, F.; Worth, A.T.; Stevenson, G.T. Bispecific F(ab’ gamma)2 antibody for the delivery of saporin in the treatment of lymphoma. J. Immunol. 1988, 141, 3662–3670. [Google Scholar]
- Flavell, D.J.; Noss, A.; Pulford, K.A.F.; Ling, N.; Flavell, S.U. Systemic delivery with 3BIT, a triple combination cocktail of anti-CD19, -CD22, and -CD38-saporin immunotoxins, is curative of human B-cell lymphoma in severe combined immunode-ficient mice. Cancer Res. 1997, 57, 4824–4829. [Google Scholar]
- Polito, L.; Bortolotti, M.; Pedrazzi, M.; Bolognesi, A. Immunotoxins and Other Conjugates Containing Saporin-S6 for Cancer Therapy. Toxins 2011, 3, 697–720. [Google Scholar] [CrossRef]
- French, R.R.; Bell, A.J.; Hamblin, T.J.; Tutt, A.L.; Glennie, M.J. Response of B-cell lymphoma to a combination of bispecific antibodies and saporin. Leuk. Res. 1996, 20, 607–617. [Google Scholar] [CrossRef]
- Stirpe, F.; Derenzini, M.; Barbieri, L.; Farabegoli, F.; Brown, A.N.F.; Knowles, P.P.; Thorpe, P.E. Hepatotoxicity of immunotoxins made with saporin, a ribosome-inactivating protein from Saponaria officinalis. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1987, 53, 259–271. [Google Scholar] [CrossRef]
- Lappi, D.; Matsunami, R.; Martineau, D.; Baird, A. Reducing the Heterogeneity of Chemically Conjugated Targeted Toxins: Homogeneous Basic FGF-Saporin. Anal. Biochem. 1993, 212, 446–451. [Google Scholar] [CrossRef]
- Zuppone, S.; Assalini, C.; Minici, C.; Bertagnoli, S.; Branduardi, P.; Degano, M.; Fabbrini, M.S.; Montorsi, F.; Salonia, A.; Vago, R. The anti-tumoral potential of the saporin-based uPAR-targeting chimera ATF-SAP. Sci. Rep. 2020, 10, 2521. [Google Scholar] [CrossRef]
- Zarovni, N.; Vago, R.; Soldà, T.; Monaco, L.; Fabbrini, M.S. Saporin as a novel suicide gene in anticancer gene therapy. Cancer Gene Ther. 2007, 14, 165–173. [Google Scholar] [CrossRef]
- Mitdank, H.; Tröger, M.; Sonntag, A.; Shirazi, N.A.; Woith, E.; Fuchs, H.; Kobelt, D.; Walther, W.; Weng, A. Suicide nanoplasmids coding for ribosome-inactivating proteins. Eur. J. Pharm. Sci. 2022, 170, 106107. [Google Scholar] [CrossRef]
- Weng, A.; Bachran, C.; Fuchs, H.; Krause, E.; Stephanowitz, H.; Melzig, M. Enhancement of saporin cytotoxicity by Gypsophila saponins—More than stimulation of endocytosis. Chem. Interact. 2009, 181, 424–429. [Google Scholar] [CrossRef]
- Wiley, R.G.; Oeltmann, T.N.; Lappi, D. Immunolesioning: Selective destruction of neurons using immunotoxin to rat NGF receptor. Brain Res. 1991, 562, 149–153. [Google Scholar] [CrossRef]
- Matchynski, J.J.; Lowrance, S.A.; Pappas, C.; Rossignol, J.; Puckett, N.; Sandstrom, M.; Dunbar, G.L. Combinatorial Treatment of Tart Cherry Extract and Essential Fatty Acids Reduces Cognitive Impairments and Inflammation in the mu-p75 Saporin-Induced Mouse Model of Alzheimer’s Disease. J. Med. Food 2013, 16, 288–295. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Parpura, V.; Rodriguez-Arellano, J.; Zorec, R. Astroglia in Alzheimer’s Disease. In Neuroglia in Neurodegen-erative Diseases; Verkhratsky, A., Ho, M., Zorec, R., Parpura, V., Eds.; Springer: Singapore, 2019; Volume 1175, pp. 273–324. [Google Scholar]
- Fisher, C.E.; Sutherland, J.A.; Krause, J.E.; Murphy, J.R.; Leeman, S.E.; Vanderspek, J.C. Genetic construction and properties of a diphtheria toxin-related substance P fusion protein: In vitro destruction of cells bearing substance P receptors. Proc. Natl. Acad. Sci. USA 1996, 93, 7341–7345. [Google Scholar] [CrossRef]
- Wiley, R.; Lappi, D. Destruction of neurokinin-1 receptor expressing cells in vitro and in vivo using substance P-saporin in rats. Neurosci. Lett. 1997, 230, 97–100. [Google Scholar] [CrossRef]
- Mantyh, P.W.; Rogers, S.D.; Honore, P.; Allen, B.J.; Ghilardi, J.R.; Li, J.; Daughters, R.S.; Lappi, D.A.; Wiley, R.G.; Simone, D.A. Inhibition of Hyperalgesia by Ablation of Lamina I Spinal Neurons Expressing the Substance P Receptor. Science 1997, 278, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Nichols, M.L.; Allen, B.J.; Rogers, S.D.; Ghilardi, J.R.; Honore, P.; Luger, N.M.; Finke, M.P.; Li, J.; Lappi, D.A.; Simone, D.A.; et al. Transmission of Chronic Nociception by Spinal Neurons Expressing the Substance P Receptor. Science 1999, 286, 1558–1561. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-G.; Zhao, Z.-Q.; Meng, X.-L.; Yin, J.; Liu, X.-Y.; Chen, Z.-F. Cellular Basis of Itch Sensation. Science 2009, 325, 1531–1534. [Google Scholar] [CrossRef]
- Akiyama, T.; Nguyen, T.; Curtis, E.; Nishida, K.; Devireddy, J.; Delahanty, J.; Carstens, M.I.; Carstens, E. A central role for spinal dorsal horn neurons that express neurokinin-1 receptors in chronic itch. Pain 2015, 156, 1240–1246. [Google Scholar] [CrossRef]
- Carstens, E.E.; Carstens, M.I.; Simons, C.T.; Jinks, S.L. Dorsal horn neurons expressing NK-1 receptors mediate scratching in rats. Neuroreport 2010, 21, 303–308. [Google Scholar] [CrossRef]
- Wrenn, C.C.; Picklo, M.J.; Lappi, D.; Robertson, D.; Wiley, R.G. Central noradrenergic lesioning using anti-DBH-saporin: Anatomical findings. Brain Res. 1996, 740, 175–184. [Google Scholar] [CrossRef]
- Emanuel, A.; Ritter, S. Hindbrain Catecholamine Neurons Modulate the Growth Hormone but Not the Feeding Response to Ghrelin. Endocrinology 2010, 151, 3237–3246. [Google Scholar] [CrossRef]
- Baxter, M.G.; Bucci, D.J.; Gorman, L.K.; Wiley, R.G.; Gallagher, M. Selective immunotoxic lesions of basal forebrain cholinergic cells: Effects on learning and memory in rats. Behav. Neurosci. 1995, 109, 714–722. [Google Scholar] [CrossRef]
- Zhang, K.; Fu, H.; Xing, C.; Luo, Y.; Cheng, F.-F.; Fu, Q.; Huang, Y.; Qiu, L. “Don’t eat me/eat me”-combined apoptotic body analogues for efficient targeted therapy of triple-negative breast cancer. J. Mater. Chem. B 2021, 9, 8472–8479. [Google Scholar] [CrossRef]
- Yan, Y.; Zhou, L.; Sun, Z.; Song, D.; Cheng, Y. Targeted and intracellular delivery of protein therapeutics by a boronated polymer for the treatment of bone tumors. Bioact. Mater. 2022, 7, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, K.; Obuki, M.; Sumi, H.; Klußmann, M.; Morimoto, K.; Nakai, S.; Hashimoto, T.; Fujiwara, D.; Fujii, I.; Yuba, E.; et al. Macropinocytosis-Inducible Extracellular Vesicles Modified with Antimicrobial Protein CAP18-Derived Cell-Penetrating Peptides for Efficient Intracellular Delivery. Mol. Pharm. 2021, 18, 3290–3301. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Jothar, L.; Beztsinna, N.; Van Nostrum, C.F.; Hennink, W.E.; Oliveira, S. Selective Cytotoxicity to HER2 Positive Breast Cancer Cells by Saporin-Loaded Nanobody-Targeted Polymeric Nanoparticles in Combination with Photochemical Internalization. Mol. Pharm. 2019, 16, 1633–1647. [Google Scholar] [CrossRef]
- Ding, L.; Jiang, Y.; Zhang, J.; Klok, H.-A.; Zhong, Z. pH-Sensitive Coiled-Coil Peptide-Cross-Linked Hyaluronic Acid Nanogels: Synthesis and Targeted Intracellular Protein Delivery to CD44 Positive Cancer Cells. Biomacromolecules 2018, 19, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Paulson, J.C. CD22 Ligands on a Natural N-Glycan Scaffold Efficiently Deliver Toxins to B-Lymphoma Cells. J. Am. Chem. Soc. 2017, 139, 12450–12458. [Google Scholar] [CrossRef] [PubMed]
- Nakase, I.; Noguchi, K.; Fujii, I.; Futaki, S. Vectorization of biomacromolecules into cells using extracellular vesicles with en-hanced internalization induced by macropinocytosis. Sci. Rep. 2016, 6, 34937. [Google Scholar] [CrossRef]
- Walseng, E.; Wälchli, S.; Fallang, L.-E.; Yang, W.; Vefferstad, A.; Areffard, A.; Olweus, J. Soluble T-Cell Receptors Produced in Human Cells for Targeted Delivery. PLoS ONE 2015, 10, e0119559. [Google Scholar] [CrossRef]
- Dementia. Available online: https://www.who.int/news-room/fact-sheets/detail/dementia (accessed on 27 July 2021).
- WHO. Global Action Plan on the Public Health Response to Dementia 2017–2025; WHO Document Production Services: Geneva, Switzerland, 2017. [Google Scholar]
- Alzheimer’s Association: Milestones. Available online: https://www.alz.org/alzheimers-dementia/research_progress/milestones (accessed on 15 October 2021).
- Rossner, S.; Roßner, S. Cholinergic immunolesions by 192IgG-saporin—A useful tool to simulate pathogenic aspects of alzheimer’s disease. Int. J. Dev. Neurosci. 1997, 15, 835–850. [Google Scholar] [CrossRef]
- Arendt, T. Impairment in memory function and neurodegenerative changes in the cholinergic basal forebrain system induced by chronic intake of ethanol. Cell Anim. Models Aging Dement. Res. 1994, 44, 173–187. [Google Scholar] [CrossRef]
- Giovannini, M.G.; Scali, C.; Prosperi, C.; Bellucci, A.; Vannucchi, M.G.; Rosi, S.; Pepeu, G.; Casamenti, F. β-Amyloid-Induced Inflammation and Cholinergic Hypofunction in the Rat Brain in Vivo: Involvement of the p38MAPK Pathway. Neurobiol. Dis. 2002, 11, 257–274. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Rodrigues, J.J.; Pivoriunas, A.; Zorec, R.; Semyanov, A. Astroglial atrophy in Alzheimer’s disease. Pflüg. Arch. Eur. J. Physiol. 2019, 471, 1247–1261. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.; Maloney, A.J. Selective loss of central cholinergic neurons in Alzheimer’s disease. Lancet 1976, 308, 1403. [Google Scholar] [CrossRef]
- Blanco-Centurion, C.; Xu, M.; Murillo-Rodriguez, E.; Gerashchenko, D.; Shiromani, A.M.; Salin-Pascual, R.J.; Hof, P.R.; Shiromani, P.J. Adenosine and Sleep Homeostasis in the Basal Forebrain. J. Neurosci. 2006, 26, 8092–8100. [Google Scholar] [CrossRef]
- Wrenn, C.C.; Wiley, R.G. The behavioral functions of the cholinergic basal forebrain: Lessons from 192 IgG-SAPORIN. Int. J. Dev. Neurosci. 1998, 16, 595–602. [Google Scholar] [CrossRef]
- Bigl, V.; Schliebs, R. Simulation of cortical cholinergic deficits–a novel experimental approach to study pathogenetic aspects of Alzheimer’s disease. Focus Extrapyramidal Dysfunct. 1998, 54, 237–247. [Google Scholar] [CrossRef]
- Wiley, R.; Berbos, T.; Deckwerth, T.; Johnson, E.; Lappi, D. Destruction of the cholinergic basal forebrain using immunotoxin to rat NGF receptor: Modeling the cholinergic degeneration of Alzheimer’s disease. J. Neurol. Sci. 1995, 128, 157–166. [Google Scholar] [CrossRef]
- Waite, J.; Chen, A.; Wardlow, M.; Wiley, R.; Lappi, D.; Thal, L. 192 immunoglobulin G-saporin produces graded behavioral and biochemical changes accompanying the loss of cholinergic neurons of the basal forebrain and cerebellar Purkinje cells. Neuroscience 1995, 65, 463–476. [Google Scholar] [CrossRef]
- Lee, M.; Chrobak, J.; Sik, A.; Wiley, R.; Buzsaki, G. Hippocampal theta activity following selective lesion of the septal cholinergic systeM. Neuroscience 1994, 62, 1033–1047. [Google Scholar] [CrossRef]
- Holley, L.A.; Wiley, R.G.; Lappi, D.A.; Sarter, M. Cortical cholinergic deafferentation following the intracortical infusion of 192 IgG-saporin: A quantitative histochemical study. Brain Res. 1994, 663, 277–286. [Google Scholar] [CrossRef]
- Ohtake, T.; Heckers, S.; Wiley, R.; Lappi, D.; Mesulam, M.-M.; Geula, C. Retrograde degeneration and colchicine protection of basal forebrain cholinergic neurons following hippocampal injections of an immunotoxin against the p75 nerve growth factor receptor. Neuroscience 1997, 78, 123–133. [Google Scholar] [CrossRef]
- Torres, E.; Perry, T.; Blokland, A.; Wilkinson, L.; Wiley, R.; Lappi, D.; Dunnett, S. Behavioural, histochemical and biochemical consequences of selective immunolesions in discrete regions of the basal forebrain cholinergic system. Neuroscience 1994, 63, 95–122. [Google Scholar] [CrossRef]
- Levey, A.I.; Edmunds, S.M.; Hersch, S.M.; Wiley, R.G.; Heilman, C.J. Light and electron microscopic study of m2 muscarinic acetylcholine receptor in the basal forebrain of the rat. J. Comp. Neurol. 1995, 351, 339–356. [Google Scholar] [CrossRef]
- Ho, N.F.; Han, S.P.; Dawe, G.S. Effect of voluntary running on adult hippocampal neurogenesis in cholinergic lesioned mice. BMC Neurosci. 2009, 10, 57. [Google Scholar] [CrossRef]
- Heckers, S.; Ohtake, T.; Wiley, R.; Lappi, D.; Geula, C.; Mesulam, M. Complete and selective cholinergic denervation of rat neocortex and hippocampus but not amygdala by an immunotoxin against the p75 NGF receptor. J. Neurosci. 1994, 14, 1271–1289. [Google Scholar] [CrossRef]
- Steckler, T.; Keith, A.B.; Wiley, R.G.; Sahgal, A. Cholinergic lesions by 192 IgG-saporin and short-term recognition memory: Role of the septohippocampal projection. Neuroscience 1995, 66, 101–114. [Google Scholar] [CrossRef]
- Cassel, J.-C.; Gaurivaud, M.; Lazarus, C.; Bertrand, F.; Galani, R.; Jeltsch, H. Grafts of fetal septal cells after cholinergic immunotoxic denervation of the hippocampus: A functional dissociation between dorsal and ventral implantation sites. Neuroscience 2002, 113, 871–882. [Google Scholar] [CrossRef]
- Garcia-Alloza, M.; Zaldua, M.N.; Diez-Ariza, M.; Marcos, B.B.; Lasheras, B.; Gil-Bea, F.J.; Ramirez, M.J. Effect of Selective Cholinergic Denervation on the Serotonergic System: Implications for Learning and Memory. J. Neuropathol. Exp. Neurol. 2006, 65, 1074–1081. [Google Scholar] [CrossRef]
- Rodríguez, J.J.; Noristani, H.; Verkhratsky, A. The serotonergic system in ageing and Alzheimer’s disease. Prog. Neurobiol. 2012, 99, 15–41. [Google Scholar] [CrossRef]
- Cho, J.S.; Lee, J.; Jeong, D.U.; Kim, H.W.; Chang, W.S.; Moon, J.; Chang, J.W. Effect of Placenta-Derived Mesenchymal Stem Cells in a Dementia Rat Model via Microglial Mediation: A Comparison between Stem Cell Transplant Methods. Yonsei Med. J. 2018, 59, 406–415. [Google Scholar] [CrossRef]
- Shin, J.; Kong, C.; Lee, J.; Choi, B.Y.; Sim, J.; Koh, C.S.; Park, M.; Na, Y.C.; Suh, S.W.; Chang, W.S.; et al. Focused ultrasound-induced blood-brain barrier opening improves adult hippocampal neurogenesis and cognitive function in a cholinergic degeneration dementia rat model. Alzheimer’s Res. Ther. 2019, 11, 1–15. [Google Scholar] [CrossRef]
- Jeong, D.U.; Lee, J.E.; Lee, S.E.; Chang, W.S.; Kim, S.J.; Chang, J.W. Improvements in Memory after Medial Septum Stimulation Are Associated with Changes in Hippocampal Cholinergic Activity and Neurogenesis. BioMed Res. Int. 2014, 2014, 568587. [Google Scholar] [CrossRef] [PubMed]
- Giannoni, P.; Fossati, S.; Marcello, E.; Claeysen, S. Editorial: Identification of Multiple Targets in the Fight Against Alzheimer’s Disease. Front. Aging Neurosci. 2020, 12, 169. [Google Scholar] [CrossRef] [PubMed]
- Cerovic, M.; Forloni, G.; Balducci, C. Neuroinflammation and the Gut Microbiota: Possible Alternative Therapeutic Targets to Counteract Alzheimer’s Disease? Front. Aging Neurosci. 2019, 11, 284. [Google Scholar] [CrossRef]
- Heneka, M.T.; Carson, M.J.; El Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef]
- Lambert, M.P.; Barlow, A.K.; Chromy, B.A.; Edwards, C.; Freed, R.; Liosatos, M.; Morgan, T.E.; Rozovsky, I.; Trommer, B.; Viola, K.L.; et al. Diffusible, nonfibrillar ligands derived from A 1-42 are potent central nervous system neurotoxins. Proc. Natl. Acad. Sci. USA 1998, 95, 6448–6453. [Google Scholar] [CrossRef]
- McGaughy, J.; Everitt, B.; Robbins, T.; Sarter, M. The role of cortical cholinergic afferent projections in cognition: Impact of new selective immunotoxins. Behav. Brain Res. 2000, 115, 251–263. [Google Scholar] [CrossRef]
- Beach, T.G.; Potter, E.P.; Kuo, Y.-M.; Emmerling, M.R.; Durham, A.R.; Webster, S.D.; Walker, D.G.; Sue, I.L.; Scott, S.; Layne, K.J.; et al. Cholinergic deafferentation of the rabbit cortex: A new animal model of Aβ deposition. Neurosci. Lett. 2000, 283, 9–12. [Google Scholar] [CrossRef]
- Ferreira, G.; Meurisse, M.; Tillet, Y.; Lévy, F. Distribution and co-localization of choline acetyltransferase and p75 neurotrophin receptors in the sheep basal forebrain: Implications for the use of a specific cholinergic immunotoxin. Neuroscience 2001, 104, 419–439. [Google Scholar] [CrossRef]
- Berger-Sweeney, J.; Stearns, N.A.; Murg, S.L.; Floerke-Nashner, L.R.; Lappi, D.; Baxter, M.G. Selective Immunolesions of Cholinergic Neurons in Mice: Effects on Neuroanatomy, Neurochemistry, and Behavior. J. Neurosci. 2001, 21, 8164–8173. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, D.R.; Dong, H.; Chen, L.; Lu, J.; Lazarus, M.; Cherasse, Y.; Chen, G.H.; Qu, W.M.; Huang, Z.L. Medial Para-brachial Nucleus Is Essential in Controlling Wakefulness in Rats. Front. Neurosci. 2021, 15, 645877. [Google Scholar] [CrossRef]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and Orexin Receptors: A Family of Hypothalamic Neuropeptides and G Protein-Coupled Receptors that Regulate Feeding Behavior. Cell 1998, 92, 573–585. [Google Scholar] [CrossRef]
- de Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.-B.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.F.; Gautvik, V.T.; Bartlett, F.S., III; et al. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc. Natl. Acad. Sci. USA 1998, 95, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Faraco, J.; Li, R.; Kadotani, H.; Rogers, W.; Lin, X.; Qiu, X.; de Jong, P.J.; Nishino, S.; Mignot, E. The Sleep Disorder Canine Narcolepsy Is Caused by a Mutation in the Hypocretin (Orexin) Receptor 2 Gene. Cell 1999, 98, 365–376. [Google Scholar] [CrossRef]
- Gerashchenko, D.; Kohls, M.D.; Greco, M.; Waleh, N.S.; Salin-Pascual, R.; Kilduff, T.; Lappi, D.; Shiromani, P.J. Hypocretin-2-Saporin Lesions of the Lateral Hypothalamus Produce Narcoleptic-Like Sleep Behavior in the Rat. J. Neurosci. 2001, 21, 7273–7283. [Google Scholar] [CrossRef] [PubMed]
- Murillo-Rodríguez, E.; Millán-Aldaco, D.; Palomero-Rivero, M.; Morales-Lara, D.; Mechoulam, R.; Drucker-Colín, R. Cannabidiol Partially Blocks the Excessive Sleepiness in Hypocretindeficient Rats: Preliminary Data. CNS Neurol. Disord. Drug Targets 2019, 18, 705–712. [Google Scholar] [CrossRef]
- Nirogi, R.; Shinde, A.K.; Mohammed, A.R.; Badange, R.K.; Reballi, V.; Bandyala, T.R.; Saraf, S.K.; Bojja, K.; Manchineella, S.; Achanta, P.K.; et al. Discovery and Development of N-[4-(1-Cyclobutylpiperidin-4-yloxy)phenyl]-2-(morpholin-4-yl)acetamide Dihydrochloride (SUVN-G3031): A Novel, Potent, Selective, and Orally Active Histamine H3 Receptor Inverse Agonist with Robust Wake-Promoting Activity. J. Med. Chem. 2019, 62, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Gerashchenko, D.; Blanco-Centurion, C.; Miller, J.; Shiromani, P. Insomnia following hypocretin2-saporin lesions of the substantia nigra. Neuroscience 2006, 137, 29–36. [Google Scholar] [CrossRef]
- Eikermann, M.; Vetrivelan, R.; Grosse-Sundrup, M.; Henry, M.E.; Hoffmann, U.; Yokota, S.; Saper, C.B.; Chamberlin, N.L. The ventrolateral preoptic nucleus is not required for isoflurane general anesthesia. Brain Res. 2011, 1426, 30–37. [Google Scholar] [CrossRef]
- Lind, L.A.; Murphy, E.R.; Lever, T.E.; Nichols, N.L. Hypoglossal Motor Neuron Death Via Intralingual CTB–saporin (CTB–SAP) Injections Mimic Aspects of Amyotrophic Lateral Sclerosis (ALS) Related to Dysphagia. Neuroscience 2018, 390, 303–316. [Google Scholar] [CrossRef]
- Fargo, K.N.; Sengelaub, D.R. Exogenous testosterone prevents motoneuron atrophy induced by contralateral motoneuron depletion. J. Neurobiol. 2004, 60, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Fargo, K.N.; Sengelaub, D.R. Testosterone manipulation protects motoneurons from dendritic atrophy after contralateral motoneuron depletion. J. Comp. Neurol. 2004, 469, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Gulino, R.; Vicario, N.; Giunta, M.A.S.; Spoto, G.; Calabrese, G.; Vecchio, M.; Gulisano, M.; Leanza, G.; Parenti, R. Neuro-muscular Plasticity in a Mouse Neurotoxic Model of Spinal Motoneuronal Loss. Int. J. Mol. Sci. 2019, 20, 1500. [Google Scholar] [CrossRef] [PubMed]
- Leanza, G.; Gulino, R.; Zorec, R. Noradrenergic Hypothesis Linking Neurodegeneration-Based Cognitive Decline and As-troglia. Front. Mol. Neurosci. 2018, 11, 254. [Google Scholar] [CrossRef] [PubMed]
- Gulino, R. Neuroplasticity and Repair in Rodent Neurotoxic Models of Spinal Motoneuron Disease. Neural Plast. 2016, 2016, 2769735. [Google Scholar] [CrossRef]
- Gulino, R.; Gulisano, M. Noggin and Sonic hedgehog are involved in compensatory changes within the motoneuron-depleted mouse spinal cord. J. Neurol. Sci. 2013, 332, 102–109. [Google Scholar] [CrossRef]
- Gulino, R.; Forte, S.; Parenti, R.; Gulisano, M. TDP-43 as a Modulator of Synaptic Plasticity in a Mouse Model of Spinal Motoneuron Degeneration. CNS Neurol. Disord. Drug Targets 2015, 14, 55–60. [Google Scholar] [CrossRef]
- Borkowski, L.F.; Nichols, N.L. Differential mechanisms are required for phrenic long-term facilitation over the course of motor neuron loss following CTB-SAP intrapleural injections. Exp. Neurol. 2020, 334, 113460. [Google Scholar] [CrossRef]
- Borkowski, L.F.; Smith, C.L.; Keilholz, A.N.; Nichols, N.L. Divergent receptor utilization is necessary for phrenic long-term facilitation over the course of motor neuron loss following CTB-SAP intrapleural injections. J. Neurophysiol. 2021, 126, 709–722. [Google Scholar] [CrossRef]
- Nichols, N.L.; Craig, T.A.; Tanner, M.A. Phrenic long-term facilitation following intrapleural CTB-SAP-induced respiratory motor neuron death. Respir. Physiol. Neurobiol. 2018, 256, 43–49. [Google Scholar] [CrossRef]
- Nichols, N.L.; Vinit, S.; Bauernschmidt, L.; Mitchell, G.S. Respiratory function after selective respiratory motor neuron death from intrapleural CTB–saporin injections. Exp. Neurol. 2015, 267, 18–29. [Google Scholar] [CrossRef]
- Borkowski, L.F.; Keilholz, A.N.; Smith, C.L.; Canda, K.A.; Nichols, N.L. Nonsteroidal anti-inflammatory drug (ketoprofen) delivery differentially impacts phrenic long-term facilitation in rats with motor neuron death induced by intrapleural CTB-SAP injections. Exp. Neurol. 2021, 347, 113892. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.A.; Lever, T.E.; Nichols, N.L. Tongue and hypoglossal morphology after intralingual cholera toxin B–saporin injection. Muscle Nerve 2021, 63, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Lujan, H.L.; Palani, G.; Peduzzi, J.D.; DiCarlo, S.E. Targeted ablation of mesenteric projecting sympathetic neurons reduces the hemodynamic response to pain in conscious, spinal cord-transected rats. Am. J. Physiol. Integr. Comp. Physiol. 2010, 298, R1358–R1365. [Google Scholar] [CrossRef] [PubMed]
- Lujan, H.L.; Palani, G.; Chen, Y.; Peduzzi, J.D.; Dicarlo, S.E. Targeted ablation of cardiac sympathetic neurons reduces resting, reflex and exercise-induced sympathetic activation in conscious rats. Am. J. Physiol. Circ. Physiol. 2009, 296, H1305–H1311. [Google Scholar] [CrossRef][Green Version]
- Lujan, H.L.; Palani, G.; Zhang, L.; Dicarlo, S.E. Targeted ablation of cardiac sympathetic neurons reduces the susceptibility to ischemia-induced sustained ventricular tachycardia in conscious rats. Am. J. Physiol. Circ. Physiol. 2010, 298, H1330–H1339. [Google Scholar] [CrossRef]
- Chew, C.; Sengelaub, D.R. Neuroprotective Effects of Exercise on the Morphology of Somatic Motoneurons Following the Death of Neighboring Motoneurons. Neurorehabilit. Neural Repair 2019, 33, 656–667. [Google Scholar] [CrossRef]
- Meredith, G.E.; Sonsalla, P.K.; Chesselet, M.-F. Animal models of Parkinson’s disease progression. Acta Neuropathol. 2008, 115, 385–398. [Google Scholar] [CrossRef]
- Wiley, R.G.; Harrison, M.B.; Levey, A.I.; Lappi, D. Destruction of midbrain dopaminergic neurons by using immunotoxin to dopamine transporter. Cell. Mol. Neurobiol. 2003, 23, 839–850. [Google Scholar] [CrossRef]
- Weisshaar, E. Epidemiology of Itch. Immunodefic. Ski. 2016, 50, 5–10. [Google Scholar] [CrossRef]
- Mishra, S.K.; Hoon, M.A. The Cells and Circuitry for Itch Responses in Mice. Science 2013, 340, 968–971. [Google Scholar] [CrossRef]
- Turski, W.A.; Cavalheiro, E.; Schwarz, M.; Czuczwar, S.J.; Kleinrok, Z.; Turski, L. Limbic seizures produced by pilocarpine in rats: Behavioural, electroencephalographic and neuropathological study. Behav. Brain Res. 1983, 9, 315–335. [Google Scholar] [CrossRef]
- Curia, G.; Longo, D.; Biagini, G.; Jones, R.S.; Avoli, M. The pilocarpine model of temporal lobe epilepsy. J. Neurosci. Methods 2008, 172, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Chun, E.; Bumanglag, A.V.; Burke, S.N.; Sloviter, R.S. Targeted hippocampal GABA neuron ablation by Stable Substance P–saporin causes hippocampal sclerosis and chronic epilepsy in rats. Epilepsia 2019, 60, e52–e57. [Google Scholar] [CrossRef] [PubMed]
- Kanai, T.; Watanabe, M.; Okazawa, A.; Sato, T.; Yamazaki, M.; Okamoto, S.; Ishii, H.; Totsuka, T.; Iiyama, R.; Okamoto, R.; et al. Macrophage-derived IL-18-mediated intestinal inflammation in the murine model of Crohn’s disease. Gastroenterology 2001, 121, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Kanai, T.; Uraushihara, K.; Totsuka, T.; Okazawa, A.; Hibi, T.; Oshima, S.; Miyata, T.; Nakamura, T.; Watanabe, M. Macrophage-derived IL-18 targeting for the treatment of Crohn’s disease. Curr. Drug Targets Inflamm. Allergy 2003, 2, 131–136. [Google Scholar] [CrossRef]
- Yamazaki, M.; Yajima, T.; Tanabe, M.; Fukui, K.; Okada, E.; Okamoto, R.; Oshima, S.; Nakamura, T.; Kanai, T.; Uehira, M.; et al. Mucosal T cells expressing high levels of IL-7 receptor are potential targets for treatment of chronic colitis. J. Immunol. 2003, 171, 1556–1563. [Google Scholar] [CrossRef]
- Vera-Portocarrero, L.P.; Zhang, E.T.; Ossipov, M.H.; Xie, J.Y.; King, T.; Lai, J.; Porreca, F. Descending facilitation from the rostral ventromedial medulla maintains nerve injury-induced central sensitization. Neuroscience 2006, 140, 1311–1320. [Google Scholar] [CrossRef]
- Criscimanna, A.; Coudriet, G.M.; Gittes, G.K.; Piganelli, J.D.; Esni, F. Activated macrophages create lineage-specific microenvironments for pancreatic acinar- and β-cell regeneration in mice. Gastroenterology 2014, 147, 1106–1118.e11. [Google Scholar] [CrossRef]
- Wang, B.; Miao, Y.; Zhao, Z.; Zhong, Y. Inflammatory Macrophages Promotes Development of Diabetic Encephalopathy. Cell Physiol. Biochem. 2015, 36, 1142–1150. [Google Scholar] [CrossRef]
- Diepenbroek, C.; Quinn, D.; Stephens, R.; Zollinger, B.; Anderson, S.; Pan, A.; de Lartigue, G. Validation and characterization of a novel method for selective vagal deafferentation of the gut. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G342–G352. [Google Scholar] [CrossRef]
- Suarez, A.N.; Hsu, T.M.; Liu, C.M.; Noble, E.E.; Cortella, A.M.; Nakamoto, E.M.; Hahn, J.D.; de Lartigue, G.; Kanoski, S.E. Gut vagal sensory signaling regulates hippocampus function through multi-order pathways. Nat. Commun. 2018, 9, 2181. [Google Scholar] [CrossRef] [PubMed]
- Schreihofer, A.M.; Stornetta, R.L.; Guyenet, P.G. Regulation of sympathetic tone and arterial pressure by rostral ventrolateral medulla after depletion of C1 cells in rat. J. Physiol. 2000, 529, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Schreihofer, A.M.; Guyenet, P.G. Role of presympathetic C1 neurons in the sympatholytic and hypotensive effects of clonidine in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R1753–R1762. [Google Scholar] [CrossRef] [PubMed]
- Schreihofer, A.; Guyenet, P.G. Sympathetic reflexes after depletion of bulbospinal catecholaminergic neurons with anti-DbH-saporin. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R729–R742. [Google Scholar] [CrossRef]
- Katsurada, K.; Nakata, M.; Saito, T.; Zhang, B.; Maejima, Y.; Nandi, S.S.; Sharma, N.M.; Patel, K.P.; Kario, K.; Yada, T. Central Glucagon-like Peptide-1 Receptor Signaling via Brainstem Catecholamine Neurons Counteracts Hypertension in Spontane-ously Hypertensive Rats. Sci. Rep. 2019, 9, 12986. [Google Scholar] [CrossRef]
- I’Anson, H.; Sundling, L.A.; Roland, S.M.; Ritter, S. Immunotoxic Destruction of Distinct Catecholaminergic Neuron Populations Disrupts the Reproductive Response to Glucoprivation in Female Rats. Endocrinology 2003, 144, 4325–4331. [Google Scholar] [CrossRef]
- Dudkin, K.N.; Chueva, L.V.; Makarov, F.N.; Bich, T.G.; Roer, A.E. Impairments in working memory and decision-taking pro-cesses in monkeys in a model of Alzheimer’s disease. Neurosci. Behav. Physiol. 2005, 35, 281–289. [Google Scholar] [CrossRef]
- Wiley, R.G.; Kline, I.R. Neuronal lesioning with axonally transported toxins. J. Neurosci. Methods 2000, 103, 73–82. [Google Scholar] [CrossRef]
- Mavrogenis, A.F.; Pavlakis, K.; Stamatoukou, A.; Papagelopoulos, P.J.; Theoharis, S.; Ζhang, Z.; Soucacos, P.N.; Zoubos, A.B. Intraneural OX7-saporin for neuroma-in-continuity in a rat model. Eur. J. Orthop. Surg. Traumatol. 2013, 23, 263–272. [Google Scholar] [CrossRef]
- Wiley, R.G.; Lappi, D.A. (Eds.) Molecular Surgery with Targeted Toxins; Humana Press: Totowa, NJ, USA, 2005; Volume 1, p. 311. [Google Scholar]
- Polito, L.; Bortolotti, M.; Mercatelli, D.; Battelli, M.G.; Bolognesi, A. Saporin-S6: A Useful Tool in Cancer Therapy. Toxins 2013, 5, 1698–1722. [Google Scholar] [CrossRef]
- Till, M.; May, R.D.; Uhr, J.W.; Thorpe, E.P.; Vitetta, E.S. An assay that predicts the ability of monoclonal antibodies to form potent ricin A chain-containing immunotoxins. Cancer Res. 1988, 48, 1119–1123. [Google Scholar] [PubMed]
- Kohls, M.D.; Lappi, D. Mab-Zap: A Tool for Evaluating Antibody Efficacy for Use in an Immunotoxin. Biotechniques 2000, 28, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Singer, S.J.; Nicolson, G.L. The Fluid Mosaic Model of the Structure of Cell Membranes. Science 1972, 175, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Lampert, P.W.; Joseph, B.S.; Oldstone, M.B. Antibody-induced capping of measles virus antigens on plasma membrane studied by electron microscopy. J. Virol. 1975, 15, 1248–1255. [Google Scholar] [CrossRef]
- Unanue, E.R.; Katz, D.H. Immunogenicity of macrophage-bound antigens: The requirement for hapten and carrier determinants to be on the same molecule for t and b lymphocyte collaboration. Eur. J. Immunol. 1973, 3, 559–563. [Google Scholar] [CrossRef]
- Loor, F.; Forni, L.; Pernis, B. The dynamic state of the lymphocyte membrane. Factors affecting the distribution and turnover of surface immunoglobulins. Eur. J. Immunol. 1972, 2, 203–212. [Google Scholar] [CrossRef]
- Taylor, R.B.; Duffus, W.P.H.; Raff, M.C.; De Petris, S. Redistribution and Pinocytosis of Lymphocyte Surface Immunoglobulin Molecules Induced by Anti-Immunoglobulin Antibody. Nat. New Biol. 1971, 233, 225–229. [Google Scholar] [CrossRef]
- Lu, R.-M.; Hwang, Y.-C.; Liu, I.-J.; Lee, C.-C.; Tsai, H.-Z.; Li, H.-J.; Wu, H.-C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1–30. [Google Scholar] [CrossRef]
- Allen, J.W.; Mantyh, P.W.; Horais, K.; Tozier, N.; Rogers, S.D.; Ghilardi, J.R.; Cizkova, D.; Grafe, M.R.; Richter, P.; Lappi, D.A.; et al. Safety Evaluation of Intrathecal Substance P-Saporin, a Targeted Neurotoxin, in Dogs. Toxicol. Sci. 2006, 91, 286–298. [Google Scholar] [CrossRef]
- Wiese, A.J.; Rathbun, M.; Butt, M.T.; Malkmus, S.A.; Richter, P.J.; Osborn, K.G.; Xu, Q.; Veesart, S.L.; Steinauer, J.J.; Higgins, D.; et al. Intrathecal Substance P-Saporin in the Dog. Anesthesiology 2013, 119, 1163–1177. [Google Scholar] [CrossRef]
- Frankel, A.E.; Nymeyer, H.; Lappi, D.; Higgins, D.; Ahn, C.; Noe, C. Preliminary results from a phase I study of substance P-saporin in terminal cancer patients with intractable pain. J. Clin. Oncol. 2014, 32, 191. [Google Scholar] [CrossRef]
- Noe, C. SP-SAP Human Clinical Trial for Cancer Pain–An Anesthesiologist’s Point of View. Target. Trends 2015, 16, 1–8. [Google Scholar]
- Hayashida, K. Substance P-saporin for bone cancer pain in dogs: Can man’s best friend solve the lost in translation problem in analgesic development? Anesthesiology 2013, 119, 999–1000. [Google Scholar] [CrossRef] [PubMed]
- ATS. Otis Gets His Wag Back. 2013. Available online: https://youtu.be/y7zzr2Sj1RM (accessed on 21 February 2022).
- Brown, D.C.; Agnello, K. Intrathecal substance P-saporin in the dog: Efficacy in bone cancer pain. Anesthesiology 2013, 119, 1178–1185. [Google Scholar] [CrossRef]
- Simons, C.T.; Gogineni, A.G.; Iodi Carstens, M.; Carstens, E. Reduced aversion to oral capsaicin following neurotoxic de-struction of superficial medullary neurons expressing NK-1 receptors. Brain Res. 2002, 945, 139–143. [Google Scholar] [CrossRef]
- Khasabov, S.G.; Rogers, S.D.; Ghilardi, J.R.; Peters, C.; Mantyh, P.W.; Simone, N.A. Spinal Neurons that Possess the Substance P Receptor Are Required for the Development of Central Sensitization. J. Neurosci. 2002, 22, 9086–9098. [Google Scholar] [CrossRef]
- Suzuki, R.; Morcuende, S.; Webber, M.; Hunt, S.P.; Dickenson, A.H. Superficial NK1-expressing neurons control spinal ex-citability through activation of descending pathways. Nat. Neurosci. 2002, 5, 1319–1326. [Google Scholar] [CrossRef]
- Vierck, C.; Kline, R.; Wiley, R. Intrathecal substance p–saporin attenuates operant escape from nociceptive thermal stimuli. Neuroscience 2003, 119, 223–232. [Google Scholar] [CrossRef]
- Suzuki, R.; Rahman, W.; Rygh, L.J.; Webber, M.; Hunt, S.; Dickenson, A.H. Spinal-supraspinal serotonergic circuits regulating neuropathic pain and its treatment with gabapentin. Pain 2005, 117, 292–303. [Google Scholar] [CrossRef]
- Ohtori, S.; Inoue, G.; Koshi, T.; Ito, T.; Doya, H.; Moriya, H.; Takahashi, K. Substance P-Saporin Down-Regulates Substance P Receptor Immunoreactive Sensory Dorsal Root Ganglion Neurons Innervating the Lumbar Intervertebral Discs in Rats. Spine 2006, 31, 2987–2991. [Google Scholar] [CrossRef]
- Rahman, W.; Sikandar, S.; Suzuki, R.; Hunt, S.P.; Dickenson, A.H. Superficial NK1 expressing spinal dorsal horn neurones modulate inhibitory neurotransmission mediated by spinal GABA(A) receptors. Neurosci. Lett. 2007, 419, 278–283. [Google Scholar] [CrossRef]
- Rahman, W.; Suzuki, R.; Hunt, S.P.; Dickenson, A.H. Selective ablation of dorsal horn NK1 expressing cells reveals a modu-lation of spinal alpha2-adrenergic inhibition of dorsal horn neurones. Neuropharmacology 2008, 54, 1208–1214. [Google Scholar] [CrossRef] [PubMed]
- Rivat, C.; Vera-Portocarrero, L.; Ibrahim, M.; Mata, M.; Stagg, N.; De Felice, M.; Porreca, F.; Malan, T. Spinal NK1 recep-tor-expressing neurons and descending pathways support fentanyl-induced pain hypersensitivity in a rat model of postoper-ative pain. Eur. J. Neurosci. 2009, 28, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.L.; Sloviter, R.S. Focal inhibitory interneuron loss and principal cell hyperexcitability in the rat hippocampus after microinjection of a neurotoxic conjugate of saporin and a peptidase-resistant analog of Substance P. J. Comp. Neurol. 2001, 436, 127–152. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Germanson, T.P.; Guyenet, P.G. Depressor and tachypneic responses to chemical stimulation of the ventral respir-atory group are reduced by ablation of neurokinin-1 receptor-expressing neurons. J. Neurosci. 2002, 22, 3755–3764. [Google Scholar] [CrossRef]
- Truitt, W.A.; Coolen, L.M. Identification of a Potential Ejaculation Generator in the Spinal Cord. Science 2002, 297, 1566–1569. [Google Scholar] [CrossRef]
- Stornetta, R.L.; Rosin, D.L.; Wang, H.; Sevigny, C.P.; Weston, M.C.; Guyenet, P.G. A group of glutamatergic interneurons expressing high levels of both neurokinin-1 receptors and somatostatin identifies the region of the pre-Bötzinger complex. J. Comp. Neurol. 2003, 455, 499–512. [Google Scholar] [CrossRef]
- Seki, S.; Erickson, K.A.; Seki, M.; Nishizawa, O.; Igawa, Y.; Ogawa, T.; De Groat, W.C.; Chancellor, M.B.; Yoshimura, N. Elimination of rat spinal neurons expressing neurokinin 1 receptors reduces bladder overactivity and spinal c-fos expression induced by bladder irritation. Am. J. Physiol. Physiol. 2005, 288, F466–F473. [Google Scholar] [CrossRef]
- Truitt, W.A.; Sajdyk, T.J.; Dietrich, A.D.; Oberlin, B.; McDougle, C.J.; Shekhar, A. From anxiety to autism: Spectrum of abnormal social behaviors modeled by progressive disruption of inhibitory neuronal function in the basolateral amygdala in Wistar rats. Psychopharmacology 2007, 191, 107–118. [Google Scholar] [CrossRef]
- Takakura, A.C.; Moreira, T.S.; Stornetta, R.L.; West, G.; Gwilt, J.M.; Guyenet, P.G. Selective lesion of retrotrapezoid Phox2b-expressing neurons raises the apnoeic threshold in rats. J. Physiol. 2008, 586, 2975–2991. [Google Scholar] [CrossRef]
- Darmani, N.A.; Wang, Y.; Abad, J.; Ray, A.P.; Thrush, G.R.; Ramirez, J. Utilization of the least shrew as a rapid and selective screening model for the antiemetic potential and brain penetration of substance P and NK1 receptor antagonists. Brain Res. 2008, 1214, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Xu, W.; Wang, J.; Ali, S.F.; Angulo, J.A. The neurokinin-1 receptor modulates the methamphetamine-induced striatal apoptosis and nitric oxide formation in mice. J. Neurochem. 2009, 111, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Truitt, W.A.; Johnson, P.L.; Dietrich, A.D.; Fitz, S.D.; Shekhar, A. Anxiety-like behavior is modulated by a discrete subpopu-lation of interneurons in the basolateral amygdala. Neuroscience 2009, 160, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Zipancic, I.; Calcagnotto, M.E.; Piquer-Gil, M.; Mello, L.E.; Álvarez-Dolado, M. Transplant of GABAergic Precursors Restores Hippocampal Inhibitory Function in a Mouse Model of Seizure Susceptibility. Cell Transplant. 2010, 19, 549–564. [Google Scholar] [CrossRef]
- Wilkinson, K.A.; Fu, Z.; Powell, F.L. Ventilatory effects of substance P-saporin lesions in the nucleus tractus solitarii of chron-ically hypoxic rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R343–R350. [Google Scholar] [CrossRef]
- Abbott, S.B.; Kanbar, R.; Bochorishvili, G.; Coates, M.B.; Stornetta, R.L.; Guyenet, P.G. C1 neurons excite locus coeruleus and A5 noradrenergic neurons along with sympathetic outflow in rats. J. Physiol. 2012, 590, 2897–2915. [Google Scholar] [CrossRef]
- Feetham, C.H.; Barrett-Jolley, R. NK1-receptor-expressing paraventricular nucleus neurones modulate daily variation in heart rate and stress-induced changes in heart rate variability. Physiol. Rep. 2014, 2, e12207. [Google Scholar] [CrossRef]
- Cordier, D.; Gerber, A.; Kluba, C.; Bauman, A.; Hutter, G.; Mindt, T.; Mariani, L. Expression of Different Neurokinin-1 Receptor (NK1R) Isoforms in Glioblastoma Multiforme: Potential Implications for Targeted Therapy. Cancer Biother. Radiopharm. 2014, 29, 221–226. [Google Scholar] [CrossRef]
- Fu, C.; Xue, J.; Wang, R.; Chen, J.; Ma, L.; Liu, Y.; Wang, X.; Guo, F.; Zhang, Y.; Zhang, X.; et al. Chemosensitive Phox2b-expressing neurons are crucial for hypercapnic ventilatory response in the nucleus tractus solitarius. J. Physiol. 2017, 595, 4973–4989. [Google Scholar] [CrossRef]
- Souza, G.; Stornetta, R.L.; Stornetta, D.S.; Abbott, S.B.G.; Guyenet, P.G. Contribution of the Retrotrapezoid Nucleus and Ca-rotid Bodies to Hypercapnia- and Hypoxia-induced Arousal from Sleep. J. Neurosci. 2019, 39, 9725–9737. [Google Scholar] [CrossRef]
- Díaz, H.S.; Andrade, D.C.; Toledo, C.; Pereyra, K.V.; Schwarz, K.G.; Díaz-Jara, E.; Lucero, C.; Arce-Álvarez, A.; Schultz, H.D.; Silva, J.N.; et al. Episodic stimulation of central chemoreceptor neurons elicits disordered breathing and autonomic dysfunction in volume overload heart failure. Am. J. Physiol. Cell. Mol. Physiol. 2020, 318, L27–L40. [Google Scholar] [CrossRef] [PubMed]
- Acton, D.; Ren, X.; Di Costanzo, S.; Dalet, A.; Bourane, S.; Bertocchi, I.; Eva, C.; Goulding, M. Spinal Neuropeptide Y1 Receptor-Expressing Neurons Form an Essential Excitatory Pathway for Mechanical Itch. Cell Rep. 2019, 28, 625–639.e6. [Google Scholar] [CrossRef] [PubMed]
- Driessen, A.K.; McGovern, A.E.; Behrens, R.; Moe, A.A.K.; Farrell, M.J.; Mazzone, S.B. A role for neurokinin 1 receptor expressing neurons in the paratrigeminal nucleus in bradykinin-evoked cough in guinea-pigs. J. Physiol. 2020, 598, 2257–2275. [Google Scholar] [CrossRef] [PubMed]
- Ming, X.; Prasad, N.; Thulasi, V.; Elkins, K.; Shivamurthy, V.K.N. Possible contribution of cerebellar disinhibition in epilepsy. Epilepsy Behav. 2021, 118, 107944. [Google Scholar] [CrossRef]
- Arslan, I.; Akgul, H.; Kara, M. Saporin, a Polynucleotide–Adenosine Nucleosidase, May Be an Efficacious Therapeutic Agent for SARS-CoV-2 Infection. SLAS Discov. Adv. Sci. Drug Discov. 2021, 26, 330–335. [Google Scholar] [CrossRef]
- Citores, L.; Iglesias, R.; Ferreras, J. Antiviral Activity of Ribosome-Inactivating Proteins. Toxins 2021, 13, 80. [Google Scholar] [CrossRef]
- Krivdova, G.; Kira, C.; Neller, M.; Parikh, B.A.; Hudak, K.A. Antiviral and Antifungal Properties of RIPs. In Ribosome Inacti-vating Proteins: Ricin and Related Proteins; Stirpe, F., Lappi, D.A., Eds.; John Wiley & Sons: Pondicherry, India, 2014; pp. 198–211. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area of Injection | Lesioned Cells | Reference |
|---|---|---|
| Intrapleural | Respiratory Motoneurons | [101,102,103,104,105] |
| Intralingual | Hypoglossal Motoneurons | [93,106] |
| Celiac Ganglia | Mesenteric Projecting Sympathetic Neurons | [107] |
| Bilateral Stellate Ganglia | Cardiac Sympathetic Neurons | [107,108,109] |
| Gastrocnemius Muscle | Spinal Motoneurons | [96] |
| Bulbocavernosus Muscle | Spinal Nucleus of the Bulbocavernosus Motoneurons | [94,95] |
| Vastus Medialis Muscle | Quadricep Motoneurons | [110] |
| Targeting Agent | Size |
|---|---|
| Antibody: Whole IgG | 160 kDa |
| Antibody: F(ab’)2 | 110 kDa |
| Antibody: F(ab) | 55 kDa |
| Antibody: single-chain variable fragment (scFv) | 28 kDa |
| Lectin (e.g., Isolectin B4) | 28 kDa |
| Growth Factor (e.g., Fibroblast Growth Factor (FGF) | 16.5 kDa |
| RNA Aptamers | 13–17 kDa |
| Peptides (e.g., Epidermal Growth Factor (EGF) | 2–6 kDa |
| Extracellular vesicles (EVs) | |
| Quantum Dots |
| Year | Application | Citation |
|---|---|---|
| 2001 | Focal inhibitory interneuron loss and principal cell hyperexcitability in the rat hippocampus after microinjection of a neurotoxic conjugate of Saporin and a peptidase-resistant analog of Substance P. | [160] |
| 2002 | Depressor and tachypneic responses to chemical stimulation of the ventral respiratory group are reduced by ablation of neurokinin-1 receptor-expressing neurons. | [161] |
| 2002 | Identification of a potential ejaculation generator in the spinal cord. | [162] |
| 2003 | A group of glutamatergic interneurons expressing high levels of both neurokinin-1 receptors and somatostatin identifies the region of the pre-Bötzinger complex. | [163] |
| 2005 | Elimination of rat spinal neurons expressing neurokinin 1 receptors reduces bladder overactivity and spinal c-fos expression induced by bladder irritation. | [164] |
| 2007 | From anxiety to autism: spectrum of abnormal social behaviors modeled by progressive disruption of inhibitory neuronal function in the basolateral amygdala in Wistar rats. | [165] |
| 2008 | Selective lesion of retrotrapezoid Phox2b-expressing neurons raises the apnoeic threshold in rats. | [166] |
| 2008 | Utilization of the least shrew as a rapid and selective screening model for the antiemetic potential and brain penetration of substance P and NK1 receptor antagonists. | [167] |
| 2009 | The neurokinin-1 receptor modulates the methamphetamine-induced striatal apoptosis and nitric oxide formation in mice. | [168] |
| 2009 | Anxiety-like behavior is modulated by a discrete subpopulation of interneurons in the basolateral amygdala. | [169] |
| 2010 | Transplant of GABAergic precursors restores hippocampal inhibitory function in a mouse model of seizure susceptibility. | [170] |
| 2011 | Ventilatory effects of Substance P-Saporin lesions in the nucleus tractus solitarius of chronically hypoxic rats. | [171] |
| 2012 | C1 neurons excite locus coeruleus and A5 noradrenergic neurons along with sympathetic outflow in rats. | [172] |
| 2014 | NK1-receptor-expressing paraventricular nucleus neurons modulate daily variation in heart rate and stress-induced changes in heart rate variability. | [173] |
| 2014 | Expression of different neurokinin-1 receptor (NK1R) isoforms in glioblastoma multiforme: potential implications for targeted therapy. | [174] |
| 2017 | Chemosensitive Phox2b-expressing neurons are crucial for hypercapnic ventilatory response in the nucleus tractus solitarius. | [175] |
| 2019 | contribution of the retrotrapezoid nucleus and carotid bodies to hypercapnia- and hypoxia-induced arousal from sleep. | [176] |
| 2019 | Episodic stimulation of central chemoreflex elicits long-term breathing disorders and autonomic imbalance in heart failure rats. | [177] |
| 2019 | Targeted hippocampal GABA neuron ablation by Stable Substance P-Saporin causes hippocampal sclerosis and chronic epilepsy in rats. (A new, stable model of temporal lobe epilepsy.) | [117] |
| 2019 | Spinal neuropeptide Y1 receptor-expressing neurons form an essential excitatory pathway for mechanical itch. | [178] |
| 2020 | A role for neurokinin-1 receptor expressing neurons in the paratrigeminal nucleus in bradykinin-evoked cough in guinea-pigs. | [179] |
| 2021 | Possible contribution of cerebellar disinhibition in epilepsy. | [180] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ancheta, L.R.; Shramm, P.A.; Bouajram, R.; Higgins, D.; Lappi, D.A. Saporin as a Commercial Reagent: Its Uses and Unexpected Impacts in the Biological Sciences—Tools from the Plant Kingdom. Toxins 2022, 14, 184. https://doi.org/10.3390/toxins14030184
Ancheta LR, Shramm PA, Bouajram R, Higgins D, Lappi DA. Saporin as a Commercial Reagent: Its Uses and Unexpected Impacts in the Biological Sciences—Tools from the Plant Kingdom. Toxins. 2022; 14(3):184. https://doi.org/10.3390/toxins14030184
Chicago/Turabian StyleAncheta, Leonardo R., Patrick A. Shramm, Raschel Bouajram, Denise Higgins, and Douglas A. Lappi. 2022. "Saporin as a Commercial Reagent: Its Uses and Unexpected Impacts in the Biological Sciences—Tools from the Plant Kingdom" Toxins 14, no. 3: 184. https://doi.org/10.3390/toxins14030184
APA StyleAncheta, L. R., Shramm, P. A., Bouajram, R., Higgins, D., & Lappi, D. A. (2022). Saporin as a Commercial Reagent: Its Uses and Unexpected Impacts in the Biological Sciences—Tools from the Plant Kingdom. Toxins, 14(3), 184. https://doi.org/10.3390/toxins14030184