Abstract
The neurotoxic alkaloid β-N-methyl-amino-l-alanine (BMAA) and related isomers, including N-(2-aminoethyl glycine) (AEG), β-amino-N-methyl alanine (BAMA), and 2,4-diaminobutyric acid (DAB), have been reported previously in cyanobacterial samples. However, there are conflicting reports regarding their occurrence in surface waters. In this study, we evaluated the impact of amending lake water samples with trichloroacetic acid (0.1 M TCA) on the detection of BMAA isomers, compared with pre-existing protocols. A sensitive instrumental method was enlisted for the survey, with limits of detection in the range of 5–10 ng L−1. Higher detection rates and significantly greater levels (paired Wilcoxon’s signed-rank tests, p < 0.001) of BMAA isomers were observed in TCA-amended samples (method B) compared to samples without TCA (method A). The overall range of B/A ratios was 0.67–8.25 for AEG (up to +725%) and 0.69–15.5 for DAB (up to +1450%), with absolute concentration increases in TCA-amended samples of up to +15,000 ng L−1 for AEG and +650 ng L−1 for DAB. We also documented the trends in the occurrence of BMAA isomers for a large breadth of field-collected lakes from Brazil, Canada, France, Mexico, and the United Kingdom. Data gathered during this overarching campaign (overall, n = 390 within 45 lake sampling sites) indicated frequent detections of AEG and DAB isomers, with detection rates of 30% and 43% and maximum levels of 19,000 ng L−1 and 1100 ng L−1, respectively. In contrast, BAMA was found in less than 8% of the water samples, and BMAA was not found in any sample. These results support the analyses of free-living cyanobacteria, wherein BMAA was often reported at concentrations of 2–4 orders of magnitude lower than AEG and DAB. Seasonal measurements conducted at two bloom-impacted lakes indicated limited correlations of BMAA isomers with total microcystins or chlorophyll-a, which deserves further investigation.
Keywords:
lake water; β-N-methyl-amino-l-alanine (BMAA); 2,4-diaminobutyric acid (DAB); N-(2-aminoethyl) glycine (AEG); trichloroacetic acid (TCA); temporal trends Key Contribution:
In this study, we evaluated the BMAA isomers in lake water with two protocol variations and observed that previously published procedures that did not involve TCA addition may have led to lower levels of extracellular concentrations of BMAA isomers, compared with TCA-amended samples.
1. Introduction
Cyanobacteria produce a host of secondary metabolites, some of which can present toxic effects in plants, invertebrates, and vertebrates during acute or chronic exposures [1]. The neurotoxin β-N-methyl-amino-L-alanine (BMAA) is a nonproteinogenic amino acid suspected to be implicated in the etiology of neurodegenerative diseases [2,3,4,5,6,7,8]. It was originally discovered in the Pacific Island of Guam, where it was produced by the cyanobacterium Nostoc sp. in endosymbiosis within the coralloid roots of cycads [9]. BMAA exposure (through the consumption of cycad seed flour and flying fox bats that bioconcentrated the toxin) was suggested as one of the possible factors for the high incidence rates of amyotrophic lateral sclerosis and endemic neuronal diseases observed there. BMAA and some of its structural isomers, including N-(2-aminoethyl) glycine (AEG), β-amino-N-methylalanine (BAMA), and 2,4-diaminobutyric acid (DAB), may also be produced in aquatic ecosystems during cyanobacterial harmful algal blooms (CyanoHABs). Phytoplanktonic phyla other than cyanobacteria, including ochrophyta (photosynthetic heterokonts, e.g., diatoms) and myzozoa (e.g., dinoflagellates), may also be capable of biosynthesis [10]. Hence, accidental ingestion of contaminated surface water during recreational activities and the consumption of contaminated drinking water, fish, shellfish [11], and spirulina food supplements [12] are other potential human exposure routes.
BMAA typically occurs at low–moderate levels in free-living cyanobacteria (ng g−1 to µg g−1 dry weight) [10,13,14,15,16] and presumably at much lower levels in bulk surface waters (extracellular dissolved, ng L−1 to µg L−1) [15,17,18,19]. This implies that BMAA and its isomers may escape detection in environmental waters. In addition, few studies have targeted environmental waters for BMAA monitoring, compared with biological samples [10].
A trace analysis of BMAA can be complex to implement, as described in several reviews [13,20,21]. The re-evaluation of pre-existing analytical methods has also demonstrated the potential for QA/QC failures [22,23]. Difficult chromatographic retention of underivatized BMAA and its resolution from co-occurring isomers (e.g., AEG, BAMA, and DAB) are well-known issues, but there exist other pitfalls. For instance, Roy-Lachapelle et al. [19] showed that dissolved AEG, BMAA, and DAB could suffer significant losses (>60%) with certain types of filtration materials, including glass fiber filters (GFF). As a basic amino acid, BMAA could also adhere to the surface of other glassware materials [21], potentially resulting in underreporting of concentrations. There are also cases where overreporting may be observed. Faassen et al. [22] evaluated a commercial enzyme linked immunosorbent assay (ELISA) and showed that the kit was not suitable for BMAA screening in surface waters. Recoveries were as high as 400% in spiked samples, and the ELISA kit yielded positive BMAA results in nearly all field samples (max of 300 µg L−1 for pond water), contrasting with no detections in the corresponding split samples submitted to LC-MS/MS [22]. A similar issue was recently noted by Zhang et al. [24] during the analysis of BMAA in surface runoff waters by the two instrumental techniques. The false positive results sometimes obtained with ELISA in natural waters [22,24] and other complex matrixes [25] were likely due to interferences with co-occurring matrix components. A critical review of the BMAA literature also noted serious flaws in the validation and reporting of analytical procedures [13]. The use of unsuited or insufficiently documented analytical methods may partly explain the BMAA controversy, i.e., the current lack of consensus over the widespread presence of BMAA in aquatic ecosystems [13,21].
Another critical aspect relates to the fact that BMAA can exist in different forms. For instance, for solid samples, free, soluble bound, and precipitated bound BMAA fractions have been investigated [26]. Faassen et al. confirmed that solid biological samples (cycad seeds, seafood, and exposed cladocerans) pre-treated with trichloroacetic acid (TCA) and hydrochloric acid (HCl) had BMAA concentrations greatly surpassing the free BMAA fraction [26]. Lage et al. found higher recoveries of BMAA from Spirulina powder (Anthrospira fusiformis) with an aqueous solution of 0.1 M TCA, compared with solvent mixtures without TCA (e.g., methanol/water or methanol/acetone) [27]. In the previous workflows targeting BMAA in surface water, the samples typically were not amended with strong acids [15,19,28,29]. Although SPE (on-line or off-line) was used as a pre-concentration step, BMAA was rarely detected in lake water [19,29]. BMAA isomers might be bound with metals, organic colloids, or cell residues in water, and this may lead to underestimation using the currently available procedures for surface water samples. Whether amending the lake samples with strong acid would yield different results has not been previously explored.
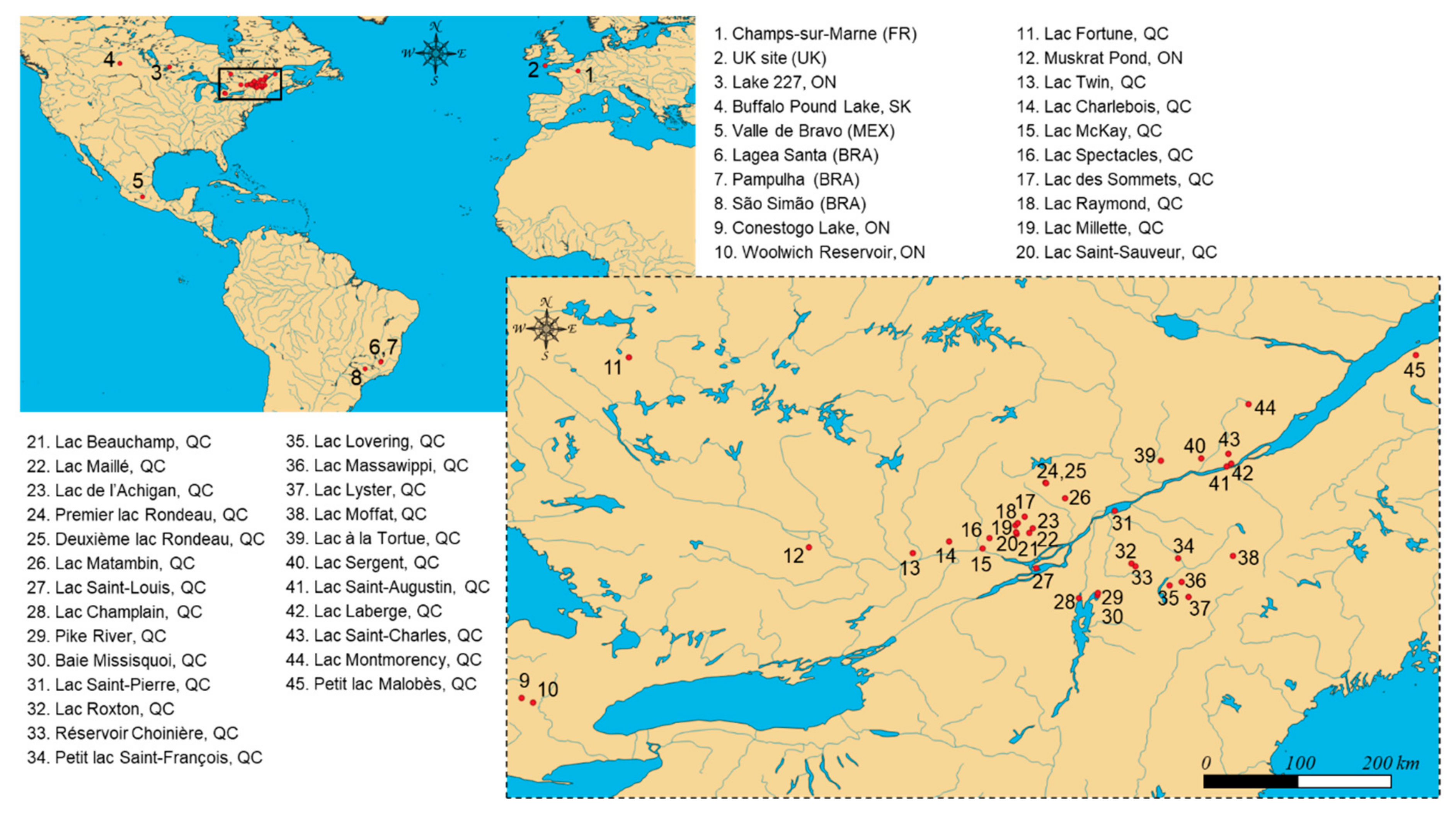
The large number of lakes targeted for cyanotoxin analysis within the framework of the ATRAPP project (Algal Blooms, Treatment, Risk Assessment, Prediction, and Prevention) provided an opportunity to address the knowledge gap. We also aimed to confirm whether BMAA and its isomers would be widely occurring in freshwater lakes impacted by harmful cyanobacterial blooms. In the first step, field-collected surface water samples were processed by two different methods to evaluate the influence of 0.1 M TCA amendment (method A: without TCA; method B: with TCA). A sensitive instrumental method relying on the on-line enrichment of derivatized BMAA/isomers and liquid chromatography high-resolution mass spectrometry was applied [29]. In the second step, we used method B (with TCA) to examine the presence of AEG, BAMA, BMAA, and DAB in a larger number of environmental water samples. The monitoring included 390 samples from 45 lakes and reservoirs of Brazil, Canada, France, Mexico, and the United Kingdom (Figure 1). To the best of our knowledge, this is the first study to report on the occurrence of BMAA isomers in lake samples at such a large spatial scale.
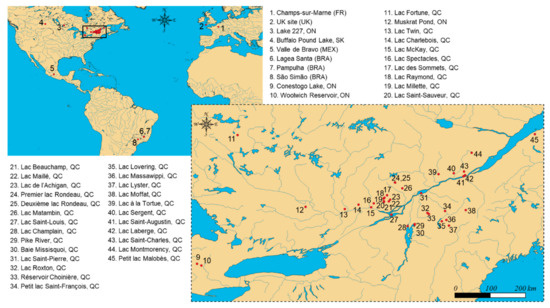
Figure 1.
Geographical location of the 45 sampling sites monitored in the present survey, including lakes and reservoirs of Brazil (BRA), Canada (Ontario (ON), Québec (QC), and Saskatchewan (SK)), France (FR), Mexico (MEX), and the United Kingdom (UK). Sampling was conducted at different locations within some lakes to account for local variations of CyanoHABs (e.g., sites #4, #5, #9, #28, #30, and #34), different water column depths (e.g., site #4), or different days during seasonal high-intensity sampling (e.g., sites #4 and #34), leading to an overall number of n = 390 environmental water samples for analysis.
2. Results and Discussion
2.1. Influence of TCA Addition
Surface water samples were analyzed using two sample preparation methods: A: without TCA amendment [19,29] and B: with 0.1 M TCA [26,30,31].
Of 158 surface water samples analyzed with both methods (Supplementary Material Table S3), 69 samples (43%) were positive for at least one of the four isomers with method A, while higher detection rates were obtained with the addition of 0.1 M TCA (119/158 positive samples or 75%). Compound-dependent detection rates were also lower using method A (AEG 19%, BAMA 3%, and DAB 35%) than method B (AEG 42%, BAMA 15%, and DAB 62%). Interestingly, BMAA itself was not detected in any of the surface water samples using either method (Supplementary Material Table S3), contrasting with the relatively widespread occurrence of its isomers AEG and DAB. In a previous study [29], we detected BMAA in only one sample (out of 82) using the same analytical approach as method A.
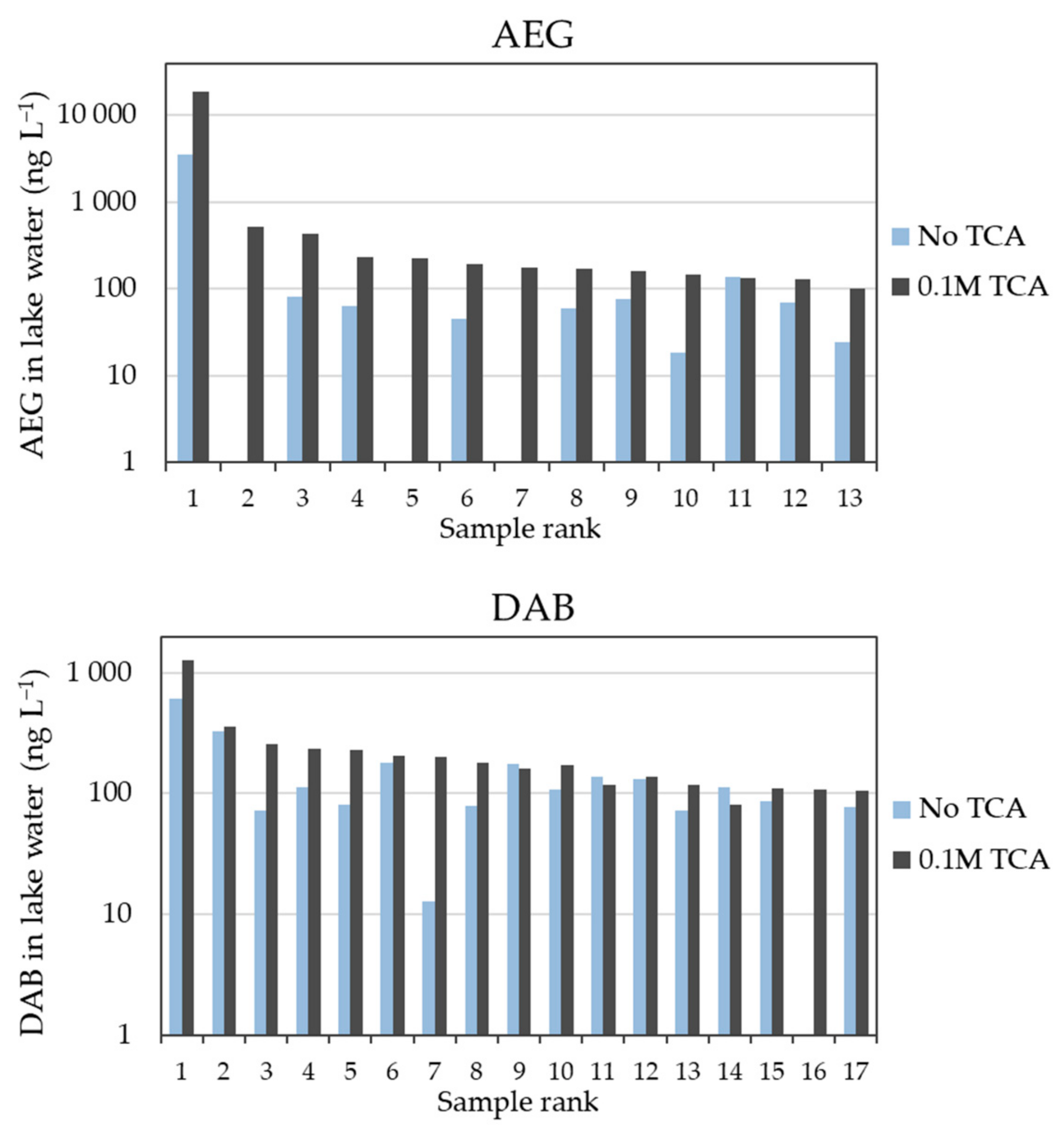
The concentrations of BMAA isomers were significantly greater using method B vs. method A (paired Wilcoxon’s signed-rank tests, p-values: pAEG and pDAB < 0.001). When a concentration value could be simultaneously returned by both methods (matching pairs calculation method), concentration ratios (B/A) greater than 1 were frequently observed. Samples with concentrations greater than 100 ng L−1 (by either method) almost systematically followed the B > A trend (Figure 2), with median ratios of 3.9 (i.e., +290% with TCA amendment) and 1.5 (+50%) for AEG and DAB, respectively. At the site location with the highest AEG concentration (Petit lac Saint-François), amending the sample with TCA led to an increase of about 5.3-times (+430%) the determined concentration (19,000 vs. 3600 ng L−1).
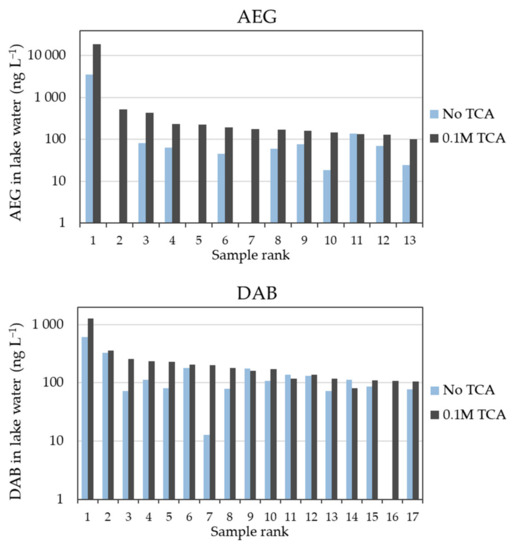
Figure 2.
Quantified concentrations (ng L−1) of AEG and DAB in surface water samples with or without 0.1 M TCA, for those samples with concentrations greater than 100 ng L−1 (in the figure x-axis, samples are ranked according to decreasing AEG and DAB concentrations by either method). Due to the ranges of concentrations in environmental samples from different locations, a logarithmic scale was applied to the y-axis. The AEG data below the detection limit of 5 ng L−1 for three samples (rank: 2, 5, and 7) without TCA added; DAB data below the detection limit of 10 ng L−1 for one sample (rank: 16) without TCA added.
Median ratios higher than their unity were also observed for the full scope of samples without concentration thresholds (i.e., median B/A of 2.4 and 1.3 for AEG and DAB, respectively). The overall range of B/A ratios were 0.67–8.25 for AEG (i.e., up to +725% with TCA) and 0.69–15.5 for DAB (i.e., up to +1450% with TCA). While a slightly negative influence (i.e., down to −33%) could be considered to fall within the analytical variation, the large increase observed at several locations could not. In addition, the use of internal standardization (BMAA-d3 spiked before derivatization) and matrix-matched calibration for each protocol likely corrected for possible instrumental variations [29].
Based on the above, we can conclude that TCA addition had a significant influence on detection rates and concentrations of BMAA isomers in lake water. To the best of our knowledge, this is the first study to report on this type of impact for water samples. The complexation of amino acids with metallic or organic ligands may be reduced under the low pH conditions of method B (pH < 1) [32] compared with method A (circumneutral pH). Low pH conditions could also favor the precipitation or denaturation of medium- and long-chain peptides [33] (e.g., originated from cellular debris) that could otherwise engage in hydrogen bond interactions. This could explain the different detection trends of BMAA isomers between the two methods. The magnitude of the increase with the TCA method was also slightly higher for samples with greater concentrations of BMAA isomers, which could reflect the enhanced sequestration potential in the more complex water bloom samples unless treated with method B. The addition of TCA may be recommended for future studies targeting dissolved BMAA in ambient water, while stronger conditions (e.g., involving additional hydrolysis with 6 M HCl) would be required for biotic tissues and lyophilized cyanobacterial samples [26].
2.2. Occurrence Trends of BMAA Isomers in 390 Lake Samples
Overall, 390 surface water samples from 45 different locations (Figure 1) were analyzed for BMAA isomers, using the 0.1 M TCA method. The associated descriptive statistics, including detection rates and concentration ranges, are summarized in Table 1. Detailed concentrations per sample and aggregated literature data from an additional 136 samples (Table 2) are also provided in the Supporting Excel data.

Table 1.
Descriptive statistics, including detection frequency (% of samples >LOD) and concentration ranges (min–max and average, ng L−1) of BMAA isomers in surface water samples (n = 390, TCA method). Site locations with concentrations punctually surpassing 100 ng L−1 are also indicated (maximum observed concentration parenthetically noted). PLSF: Petit lac Saint-Francois; BPL: Buffalo Pound Lake.
Table 2.
Overview of literature data reporting BMAA and its isomers in water samples of freshwater (lakes, rivers, and reservoirs) and saltwater (seawater) environments, including instrumental methods, limits of detection (LOD, ng L−1), type and number of field-collected samples, and concentration ranges (min-max of positive samples, ng L−1). Sample-specific concentration data of the present study and aggregated literature are also provided in the Supporting Excel data.
Of 390 samples, 222 (57%) were positive to at least one of the BMAA isomers. AEG and DAB were the most recurrently detected isomers (30% and 43% of samples with hits), while BAMA was found in only 7.4% of samples and BMAA was not found above the method detection limit (LOD of 10 ng L−1) in any sample. AEG and DAB co-occurred in 16% of the samples, while 6% of the samples had detectable levels for the three isomers simultaneously (AEG, BAMA, and DAB). The number of lakes with positive detections was 21/45 for AEG and 15/45 for DAB. BAMA was found at five site locations only (Buffalo Pound Lake, Conestogo Lake, Lac Saint-Augustin, Petit lac Saint-François, and Woolwich Reservoir).
Ten locations presented maximum concentrations of AEG and/or DAB above 100 ng L−1: Buffalo Pound Lake, Conestogo Lake, Lac Beauchamp, Lac Fortune, Lac Millette, Lac Saint-Augustin, Lac Saint-Pierre, Missisquoi Bay, Petit lac Saint-François, and the UK site. The maximum observed concentrations for each isomer were 19,000 ng L−1 for AEG (Petit lac Saint-François), 1100 ng L−1 for DAB (Petit lac Saint-François), and 56 ng L−1 for BAMA (Buffalo Pound Lake) (Table 1). The maximum concentrations of BMAA isomers in the μg L−1 range agreed with previous surveys of CyanoHAB impacted lakes and reservoirs in Canada [17,29] and the United States [18] (Table 2). Detailed concentrations per sample and aggregated literature data from an additional 136 samples (Table 2) are also provided in the Supporting Excel data.
Relatively high concentrations of AEG and DAB in some samples and the concomitant lack of BMAA detection agreed with a previous survey of lakes and rivers from Eastern Canada [29]. The results could also be related to a monitoring survey of cyanobacterial blooms in Lake Winnipeg, Manitoba, Canada [16]. In that study, the levels of AEG and DAB found in cyanobacterial samples (averages of 2120 and 170 μg g−1) were 2–3 orders of magnitude higher than those of BMAA (average of 4.05 μg g−1) [16]. A similar trend was observed in marine mats of the Arabian Gulf (Khor Al Adaid) and Australian cyanobacterial blooms, with AEG and/or DAB often 3–4 orders of magnitude higher than BMAA [15,34]. If BMAA was present in surface water from our survey, concentrations 2–4 orders of magnitude lower than AEG or DAB would fall below the detection threshold (10 ng L−1). While a few studies did report detectable levels of BMAA in lake water samples [18,19] (see also Table 2), the present study and other surveys [15,16,29,34] suggest that AEG and DAB isomers may be more prevalent than BMAA. However, monitoring of a much higher number of aquatic ecosystems would be needed for confirmation.
2.3. Monitoring of Two CyanoHAB Impacted Lakes during the Bloom Season
Monitoring conducted at two sites allowed us to examine time trends of BMAA isomers during the bloom season (Buffalo Pound Lake; BPL) or throughout the entire year (Petit lac Saint-François; PLSF, also referred to as Lake Tomcod in the literature). The selection of sampling sites for this study was aligned with previous reports of relatively high AEG/DAB levels at these two locations, compared with a range of other Canadian lakes from New Brunswick, Nova Scotia, Ontario, Québec, and Saskatchewan [16,19,29]. Both BPL and PLSF have also been experiencing significant cyanobacterial bloom-related stressors [35,36].
BPL is a shallow polymictic lake of an elongated shape (~29 km × 1 km, max. depth of 5.8 m) located in the Canadian prairies. It also serves as a major drinking water supply for the cities of Moose Jaw and Regina [37], representing 25% of Saskatchewan’s population. Occurrences of harmful cyanobacterial algal blooms in BPL and the related poisoning of dogs, cattle, and poultry were reported as early as the 1960s [37]. The BPL campaign included 49 water samples collected between 29 May and 3 October 2019, examined for BMAA isomers and other parameters (Figure 3).
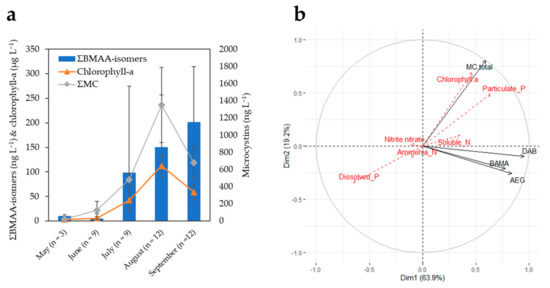
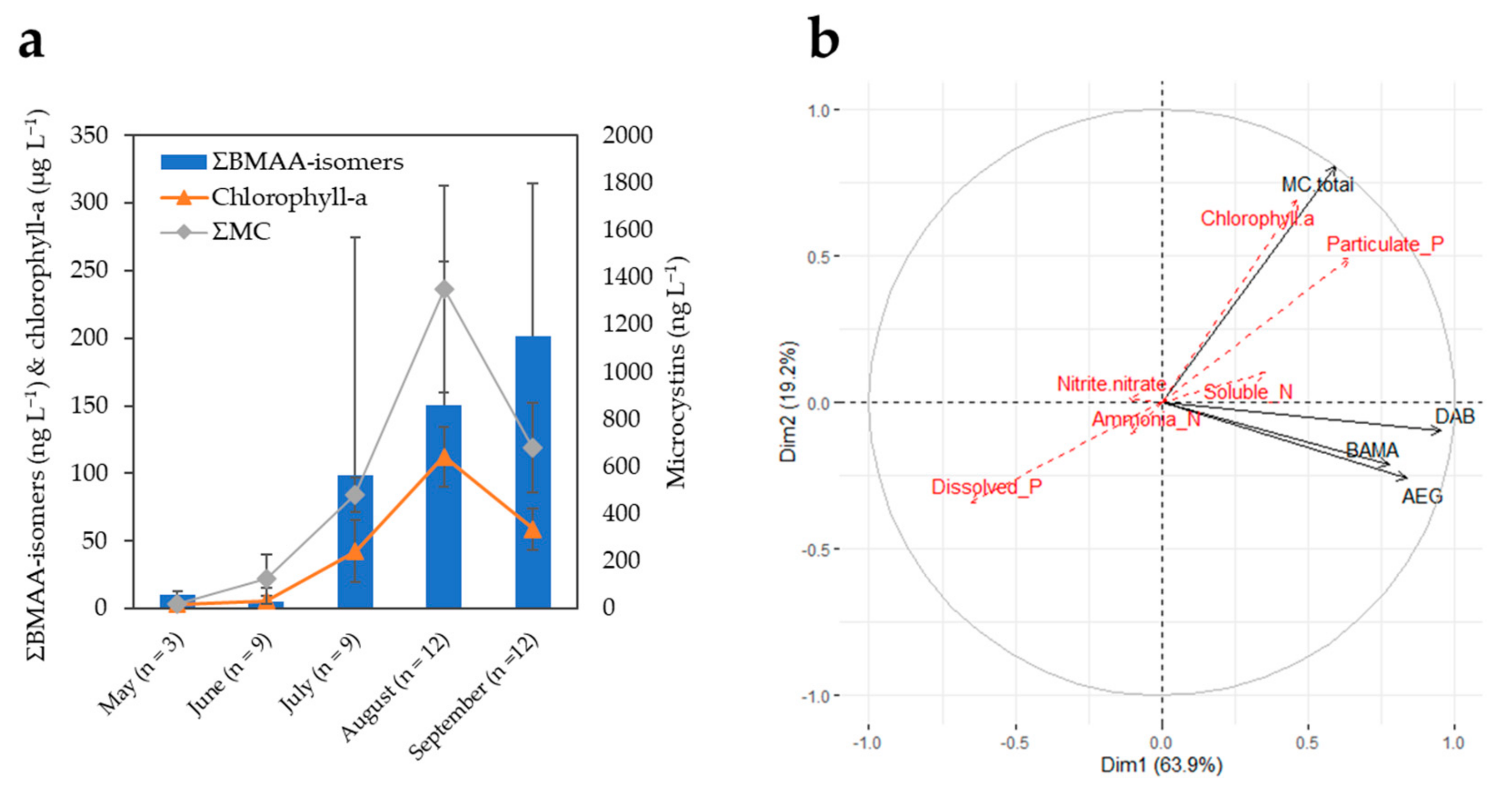
Figure 3.
(a) Monthly averages of summed BMAA isomers (ng L−1), chlorophyll-a (μg L−1), and total microcystins (ng L−1) (MMPB method) in Buffalo Pound Lake during the 2019 sampling season. Error bars are the corresponding standard deviations. Summed BMAA isomers and chlorophyll-a are plotted against the primary (left) y-axis, while the secondary (right) y-axis is used for total microcystins. (The different y-axis scales are for visualization purposes only, and they do not refer to scaled toxicities). (b) Principal component analysis (PCA) correlation circle of active variables (total MCs, AEG, BAMA, and DAB) and additional environmental variables superimposed on the plot (red font dotted arrows). To deal with different units/scales between environmental variables, data were transformed (center-reduced) prior to performing the PCA. Temperature data were not available for all dates and, therefore, could not be included in the PCA.
The monthly concentrations of summed BMAA isomers (ΣBMAA-isomers: AEG + BAMA + DAB) in BPL water samples are plotted in Figure 3a. The average concentrations of BMAA isomers remained at or below detection limits in the late spring months (~6–10 ng L−1), increased by about 10–20 times in July–August (~100–150 ng L−1) concomitantly with total microcystins (ΣMC), and were still increasing in September (~200 ng L−1), while the levels of ΣMC had already receded. In Figure 3b, vectors of AEG, BAMA, and DAB are on the same PCA correlation circle quadrant (i.e., highly correlated together) but near-orthogonal (i.e., unrelated) to those of ΣMC and chlorophyll-a. Pip et al. also observed nonsignificant or weakly significant correlations of BMAA and microcystins in water samples of Lake Winnipeg, Canada [17]. BMAA and isomers may be produced by later-blooming communities compared with those responsible for the observed peak of ΣMC. The related BPL taxonomic identification suggested a shift in community composition from Dolichospermum flos aquae to Planktothrix agardhii dominated blooms during the 2019 season, which might be associated with different toxigenic cyanopeptide profiles, as recently discussed in Painter et al. [38].
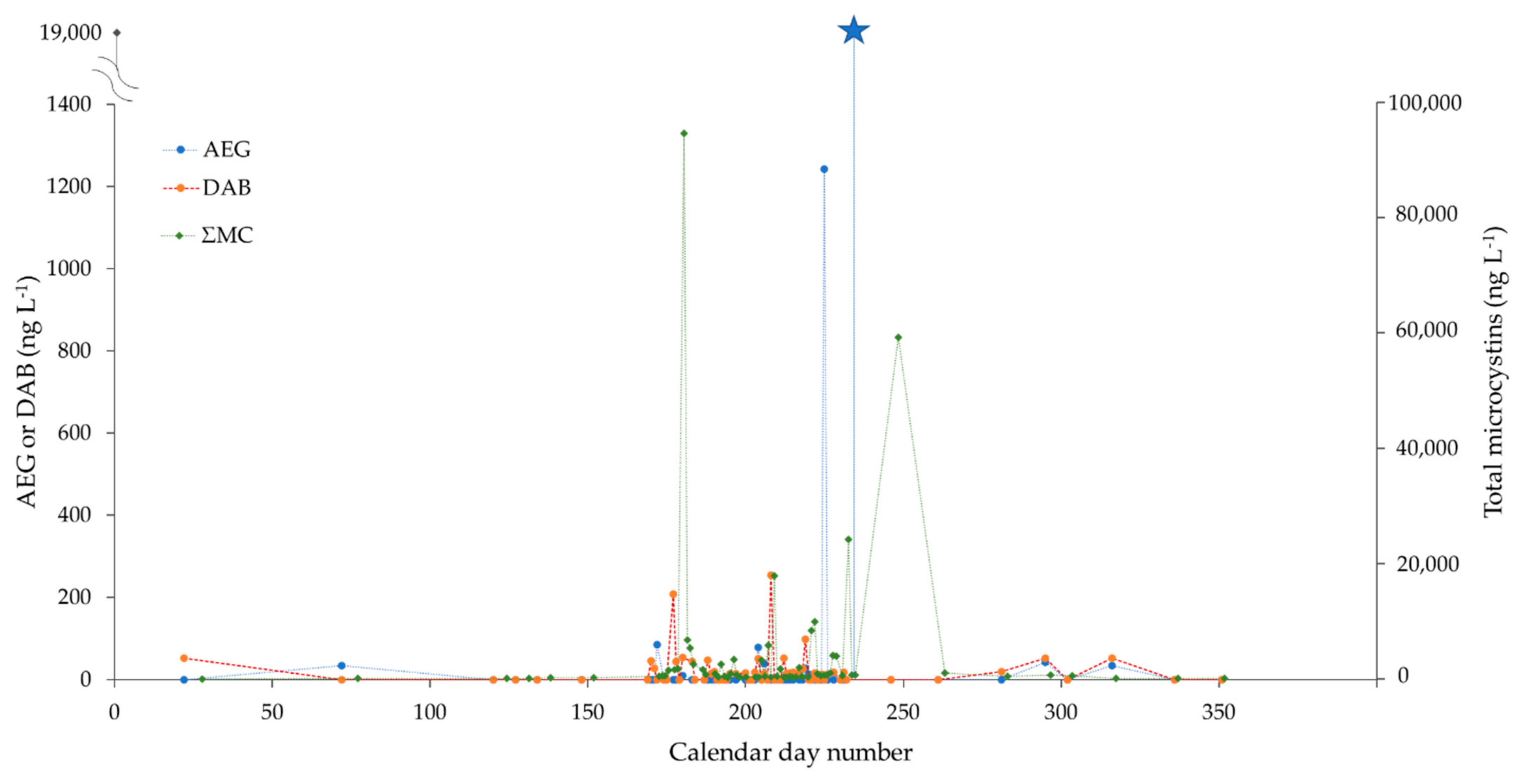
PLSF is a shallow hypereutrophic lake (1.7 km × 0.7 km, max. depth of 2 m) located in the municipality of Saint-François-Xavier-de-Brompton (Estrie, QC, Canada). Nutrient physicochemistry indicates a relatively poor water quality status and advanced eutrophication [39,40]. A subset of 66 PLSF water samples collected between 22 January and 17 December 2019 were included for analysis (Figure 4). A high sampling intensity was achieved during the two-month summer bloom period, with near-daily sample collections. Concentrations of AEG and DAB were low in the winter and spring seasons (<5–50 ng L−1), at or near the background levels of Québec rivers without known CyanoHAB impacts ([29]; see also Supporting Excel data). The AEG concentrations drastically increased at ca. 1200 ng L−1 in mid-August and then peaked to even higher levels (19,000 ng L−1) one week later, seemingly unrelated to ΣMC (Figure 4). Even after removing these two outlier points, the vector of AEG remained orthogonal to that of ΣMC (and DAB) on the PCA correlation circle (Supplementary Material Figure S2). The early summer bloom was dominated by Aphanizomenon flos-aquae, Dolichospermum spp., and Microcystis aeruginosa (June–July 2019); similar to BPL, the contribution of Planktothrix agardhii increased during the later part of the summer for PLSF. However, biomass is not per se indicative of toxin production (microcystis biomass and microcystins do, in general, show a correlation, but this does not always occur with other toxins or other cyanobacteria species). Although clearly beyond the scope of the present study, multi-year seasonal monitoring and acquisition of metagenome sequencing data may help clarify these trends.
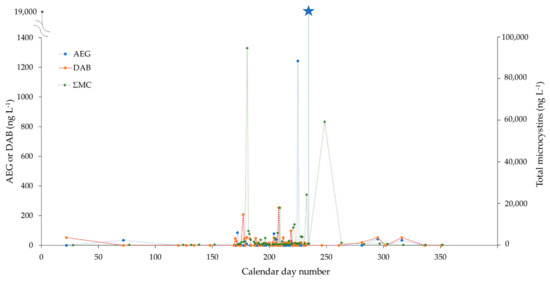
Figure 4.
Concentrations of AEG (ng L−1), DAB (ng L−1) and total microcystins (ng L−1) (MMPB method) in lake water samples of Petit lac Saint-François, collected between January and December 2019 (x-axis: calendar day number). High-frequency intensity sampling was conducted during the two-month summer bloom period (June–August), with near-daily sample collections. AEG and DAB concentrations are shown on the primary (left) y-axis, while total microcystins are plotted against the secondary (right) y-axis. (The different y-axis scales are for visualization purposes only, and they do not refer to scaled toxicities). The primary axis break was applied to plot an extremely high value of AEG observed on 20 August 2019 (19,000 ng L−1, blue star marker). For each time point and measured parameter, only a single datum point was provided; hence, error bars cannot be displayed.
3. Conclusions
In this study, we evaluated the trends of BMAA isomers in lake water with two protocol variations. We observed that previously published procedures that did not involve TCA addition may have led to lower levels of extracellular concentrations of BMAA isomers, compared with TCA-amended samples. The magnitude of the difference was variable depending on the water source but could reach values as high as 15-times for the TCA method (and a maximum observed magnitude difference of +15,000 ng/L for AEG). The standard TCA concentration used in our study was derived from the literature on BMAA analysis of solids, such as cycad seeds, shellfish, and powdered cyanobacteria [26,27]. The herein described procedure may still require additional refinement, including optimization of the TCA concentration [27] or investigation of hydrolysis under stronger conditions, such as concentrated HCl/heat [26]. Since our current method involved derivatization and a large volume injection (1 mL) by on-line SPE-UHPLC-HRMS, it was not deemed feasible to include a 6 M HCl hydrolysis step of the bulk lake water. To avoid substantial sample dilution, testing of a harsher hydrolysis step would likely necessitate pre-emptive freeze-drying of the lake water aliquots prior to reconstitution in a small volume of concentrated HCl.
The TCA method was applied to a large set of field-collected lake samples from America and Europe (overall n = 390). BMAA was not found (LOD of 10 ng L−1) in any of the 45 lakes monitored during the bloom season, while two of its isomers, AEG and DAB, could reach peak concentrations in the μg L−1 range. The results from the present study and a few others [15,16,29,34,41] indicate that BMAA may be less common in freshwater cyanobacterial algal blooms than previously thought. The most frequent occurrences of AEG and DAB were reported here for lakes with known cyanobacterial bloom impacts and sampled during the summer–fall bloom season. Much lower detection rates were observed here in the winter–spring season for the two lakes submitted to temporal monitoring. BMAA isomers are also expected to fall in the low or nondetectable ranges for other types of water bodies, including rivers [29].
While DAB may show similar neurotoxic effects as BMAA [42], in vitro bioassays suggest that AEG may be less toxic than BMAA and DAB [43,44]. Although no guidelines are currently available for BMAA in water, we note that peak levels of total microcystins far exceeded the interim drinking water advisory levels in one of the lakes targeted for seasonal monitoring. With some modifications of the sample preparation or instrumental procedures, the sensitive method could be expanded to the study of BMAA isomers in fish, shellfish, and spirulina food supplements in future work. Future surveillance and cyanotoxin risk assessment studies in bloom-impacted freshwater and marine ecosystems should preferably include analysis of BMAA isomers along with other cyanotoxins and taxonomic analysis of the phytoplankton.
4. Materials and Methods
4.1. Chemicals and Standards
N-(2-aminoethyl) glycine (AEG) was obtained from Toronto Research Chemicals Inc. (North York, ON, Canada). β-N-methylamino-L-alanine hydrochloride (L-BMAA) (purity ≥ 97.0%) and L-2-2-diaminobutyric acid dihydrochloride (DAB) (purity ≥ 95.0%) were purchased from Sigma Aldrich (Oakville, ON, Canada). β-amino-N-methyl-alanine (BAMA) was acquired from the National Research Council of Canada (Halifax, NS, Canada). The isotope-labelled internal standard L-BMAA hydrochloride-d3 (BMAA-d3) was purchased from Abraxis, Inc. (Warminster, PA, USA).
Acetonitrile (ACN), methanol (MeOH), and water of HPLC quality were purchased from Fisher Scientific (Whitby, ON, Canada). Ammonium acetate (purity ≥ 98%), sodium citrate dibasic sesquihydrate (citrate; purity 99.0%), sodium tetraborate decahydrate (borate; purity 99.5%), potassium hydroxide (KOH; purity 90%), and trichloroacetic acid (TCA; purity ≥ 99.0%) were purchased from Sigma Aldrich (Oakville, ON, Canada). The derivatizing agent 9-fluorenylmethyl chloroformate (FMOC-Cl; 98.0% purity) was obtained from Alfa AeSar (Fisher Scientific, Whitby, ON, Canada).
4.2. Sample Collections
Surface water samples (n = 390) were collected in 2016–2021 from 45 locations in Brazil, Canada (from the provinces of Ontario, Québec, and Saskatchewan), France, Mexico, and the United Kingdom. Their geographical distribution is illustrated in Figure 1. Several of these sites corresponded to freshwater lakes with a documented history of harmful cyanobacterial blooms [38,39,40,45,46,47]. Sampling was conducted as part of the ATRAPP project by trained university staff and partners. Additional water samples from bloom-impacted lakes were obtained through a citizen-science project (Adopt a Lake) [48]. Surface water samples for cyanotoxin analysis were collected in 125-mL amber polyethylene terephthalate glycol-modified (PETG) bottles, previously washed in the laboratory and rinsed in situ three times with the site surface water. Samples were kept at 4 °C and shipped within 1–3 days of collection to the analytical facilities, where they were lysed (three freeze–thaw cycles) prior to storage at −20 °C [49] until preparation and analysis.
In addition, 500-mL wide-mouth high-density polyethylene (HDPE) bottles (opaque, acid-washed) were co-collected for the analysis of the ancillary surface water parameters, such as nutrients and chlorophyll-a; filtered; and stored at 4 °C until analysis. Nutrient chemistry analyses were performed at the University of Saskatchewan Global Institute for Water Security (Saskatoon, SK, Canada), Environmental Geochemistry Laboratory University of Waterloo (Waterloo, ON, Canada) and Université de Montréal Department of Biological Sciences (Montreal, QC, Canada) using standard methods. Taxonomic analyses were conducted at Water’s Edge Scientific LLC (Baraboo, WI, USA). Preserved samples were analyzed according to APHA Method 10200F (APHA, 2012), and cell counting was performed in a Sedgewick-Rafter counting chamber.
4.3. Sample Preparation
The analysis of surface water samples by method B (with TCA) was performed as follows. An aliquot of the lysed surface water was filtered through a nitrocellulose filter (0.2 μm, 25 mm) fitted onto a syringe filter holder. A five-mL aliquot of the filtrate was then spiked with the isotopically-labelled internal standard (ILIS: BMAA-d3) to achieve an initial concentration of 500 ng L−1. TCA was added to achieve a concentration of 0.1 M [26,30], followed by a 10-min wait time. The solution was brought to a circumneutral pH with KOH prior to the addition of buffer solutions of borate (0.3 mL of a 100 mM solution) and citrate (0.3 mL of a 150 mM solution) to achieve a pH of ~9. The mixture was vortexed for 10 s (3200 rpm), followed by a five-min wait time. FMOC-Cl was added to the samples (300 μL of a 3 mg mL−1 solution prepared in ACN), and the derivatization reaction proceeded for 1 h while stirring (200 rpm, lab oven shaker; 65 °C; without light). The samples were left to cool to room temperature, and an organic cosolvent (300 µL of MeOH) was subsequently added to quench the reaction and minimize sorption losses of FMOC-derived amino acids [29]. Finally, reacted samples were vortexed (10 s; 3200 rpm) and centrifuged (10 min; 6000 rpm) prior to aliquoting of 1.5 mL of the supernatant in a two-mL LC-MS vial.
A subset of the surface water samples was also analyzed by method A (without TCA added) [29]. Lysed water samples were filtered (nitrocellulose, 0.2 μm, 25 mm), spiked with BMAA-D3 (500 ng L−1), amended with borate and citrate buffers, and derivatized with FMOC-Cl as previously described.
4.4. Instrumental Analysis
Derivatized samples were analyzed by on-line solid-phase extraction (on-line SPE) coupled with ultra-high-performance liquid chromatography high-resolution mass spectrometry (UHPLC-HRMS), adapted from Vo Duy et al. [29] with some modifications.
An injection was performed using a PAL RTC autosampler (Zwingen, Switzerland) and a one-mL stainless-steel loop (SST). The injection volume was set at 1 mL. On-line enrichment was performed with a Thermo Dionex UltiMate™ 3400 SD pump and a Thermo HyperSep Retain PEP column (hydrophilic lipophilic balance, 20 mm × 2.1 mm, particle size 40–60 μm). The loading flow rate was set at 1500 μL min−1. After sample loading, the on-line aqueous mobile phase (HPLC water) was allowed to flow for an additional 2 mL to remove salts.
The target analytes were then eluted at 450 μL min −1 in back-flush mode with the analytical mobile phase (A: 2.5 mM CH3COONH4 in HPLC water; B: ACN), using a Thermo Dionex UltiMate™ 3400 RS pump. For UHPLC separation, a Thermo Hypersil Gold C18 column (100 mm × 2.1 mm, particle size 1.9 µm, pore size 175 Å) thermostated at 35 °C was used. The UHPLC column was fitted with a 0.2-µm column prefilter. Details on the chromatographic elution gradient are provided in Supplementary Material (Table S1), as are representative UHPLC-HRMS chromatograms showing the separation of the four isomers in spiked lake water (Supplementary Material Figure S1).
Analyte detection was performed using negative electrospray ionization (ESI) and a Thermo Q-Exactive Orbitrap mass spectrometer (Thermo Scientific, San Jose, CA, USA), with a full scan MS range of m/z 200–600 and a resolution setting of 70,000 full width at half maximum (FWHM; value at m/z 200). Further details on the ESI source and MS acquisition parameters are provided in Supplementary Material (Table S1).
Total microcystins (ΣMC) were examined in conjunction with BMAA/isomers for the high-intensity sampling sites (Figure 1, sites #4 and #34). The ΣMC were analyzed in the nitrocellulose-filtered fraction of the lysed water sample. The 2-methyl-3-methoxy-4-phenylbutyric acid (MMPB) moiety generated via Lemieux-von Rudloff oxidation was analyzed using a previously validated method [50]. An aliquot of the reacted sample was spiked with ILIS (MMPB-d3) and analyzed by on-line SPE coupled to UHPLC tandem mass spectrometry (TSQ Quantiva LC-MS/MS, Thermo Scientific, San Jose, CA, USA). Further details on the analytical method for ΣMC are summarized in Supplementary Material (Text S1 and Table S2).
4.5. Quality Assurance/Quality Control (QA/QC)
The identification of BMAA and its isomers in surface water samples was based on matching retention times (±0.1 min) with calibration curve standards (e.g., Supplementary Material Figure S1) and mass accuracy of observed vs. theoretical exact m/z (tolerance of ±5 ppm).
The analytical method without TCA (method A) was previously subject to matrix-matched validation in lake water [29], including the assessment of linearity (R2 = 0.9963–0.9982), whole-method accuracy (spike level of 75 ng L−1; accuracy = 76–101%), and intermediate precision (75 ng L−1; intraday RSD of 2.1–6.7% and interday RSD of 8.1–13%). Matrix-matched calibration was adopted, and relative matrix effects (standard additions to select lake water samples) were within −22% to +17% [29].
Following the initial demonstration of method capability [29], continued QA/QC measures were implemented for the present survey, for both methods A and B. Method blanks were performed for each batch of samples using surface water aliquots from Lac Pohénégamook (QC, Canada) and submitted to the entire preparation procedure; no contamination was noted. Method detection limits in the present study were 5 ng L−1 for AEG and 10 ng L−1 for BAMA, BMAA, and DAB. An eight-point based matrix-matched calibration curve (15–1000 ng L−1, additions to a blank lake matrix from Lac Pohénégamook, QC, Canada, subsequently submitted to either method A or B) was performed at the beginning of each LC-MS sequence. Determination coefficients (R2) were within the typical range of 0.995–0.999. After the initial calibration, continued calibration verification (CCV) standards were run as matrix spikes (fortification level: 75 ng L−1). The accuracy of CCV standards was required to fall within 70–120% [51].
4.6. Data Curation and Statistical Analyses
Processing of LC-MS data was performed using the Xcalibur 4.3 software (Thermo Scientific). Statistical analyses were conducted with the R statistical software version 4.1.1 (R Core Team [52]). Statistical significance was set at p <0.05. A Principal Component Analysis (PCA) of center-reduced data was performed using the FactoMineR R-package (graphs plotted with factoextra and ggplot2). Wilcoxon’s signed rank tests for paired data were used to evaluate the statistical differences between TCA and non-TCA treatments; statistical differences could be investigated for AEG and DAB only, i.e., the two compounds with sufficient detections with both treatments. The map of sample locations was designed using Quantum GIS (QGIS 3.6 Noosa) as a geographic information system, and the base maps were obtained from Natural Earth (free vector and raster map data available at naturalearthdata.com, accessed on 6 January 2022).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/toxins14040251/s1, Text S1: Analysis of total microcystins via Lemieux-von Rudloff oxidation, Figure S1: UHPLC-HRMS chromatograms of FMOC-derivatized BMAA isomers (AEG, BAMA, BMAA, and DAB) spiked at 50 ng L−1 in blank lake water. The lower pane shows the corresponding isotope-labeled internal standard (ILIS: BMAA-D3) spiked at 500 ng L−1, Figure S2: Principal component analysis (PCA) correlation circle of active variables (total MCs, AEG, DAB) and additional environmental variables superimposed on the plot (red font dotted arrows) for the PLSF site. The PCA is applied to a subset of n = 64 samples (two outliers removed). Chlorophyll-a data were not available for all time points and therefore could not be included in the statistical analysis, Table S1: Details on the UHPLC-HRMS instrumental method for the analysis of FMOC-derivatized BMAA and its isomers, Table S2: Details on the UHPLC-MS/MS instrumental method for the analysis of total microcystins via oxidative cleavage (MMPB method), Table S3: Measured concentrations (ng L−1) of AEG, BAMA, BMAA, and DAB in samples of freshwater lakes and reservoirs, with or without amendment of 0.1M TCA. Sample-specific concentration data of the present study and aggregated literature are also provided in the Supporting Excel data.
Author Contributions
Conceptualization, S.V.D., G.M., D.F.S. and S.S.; Data curation, S.A.; Formal analysis, S.A. and G.M.; Funding acquisition, S.S.; Investigation, S.A., S.V.D. and Q.T.D.; Methodology, S.A., S.V.D. and Q.T.D.; Project administration, D.F.S. and S.S.; Resources, D.F.S., B.H., H.M.B., B.V.-L., N.F., C.W.G., M.L.L., J.J.V., F.F.M.J., A.G., C.D.L., N.T. and S.S.; Software, G.M.; Supervision, S.S.; Validation, S.V.D. and Q.T.D.; Visualization, S.A. and G.M.; Writing—original draft, S.A.; Writing—review & editing, S.A., S.V.D., G.M., Q.T.D., D.F.S., B.H., H.M.B., B.V.-L., N.F., C.W.G., M.L.L., J.J.V., F.F.M.J., A.G., C.D.L., N.T. and S.S. All authors have read and agreed to the published version of the manuscript.
Funding
Genome Canada and Génome Québec (LSARP 2016); Natural Sciences and Engineering Research Council of Canada (NSERC-CRSNG; STPGP 478774); Quebec Research Fund (FRQ; PR-183278); Canada Foundation for Innovation (CFI; 30044); Global Water Futures project FORMBLOOM (Canada First Research Excellence Fund); Furnas Centais Eletricas SA (FURNAS), Companhia Eletrica de Minas Gerais (CEMIG); Fundação de Amparo a Pesquisa de Minas Gerais (FAPEMIG); National Council of Science and Technology (CONACYT) México; ANSWER project (France, ANR-16-CE32-0009-02).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available as a Supporting Information Excel file.
Acknowledgments
This study was conducted within the framework of the ATRAPP project (Algal Blooms, Treatment, Risk Assessment, Prediction and Prevention through Genomics), with financial support from Genome Canada and Génome Québec (LSARP 2016). We also acknowledge funding from the Natural Sciences and Engineering Research Council of Canada (NSERC-CRSNG; STPGP 478774), the Quebec Research Fund (FRQ; PR-183278), the Canada Foundation for Innovation (CFI; 30044), the Global Water Futures project FORMBLOOM (Canada First Research Excellence Fund), Furnas Centais Eletricas SA (FURNAS), Companhia Eletrica de Minas Gerais (CEMIG), Fundação de Amparo a Pesquisa de Minas Gerais (FAPEMIG) and the National Council of Science and Technology (CONACYT) México. The samples from the French lake were collected within the framework of the ANSWER project (ANR-16-CE32-0009-02). We thank Katy Nugent, Cameron Hoggarth and others (University of Saskatchewan), Richard Elgood (University of Waterloo), Roxane Poirier (Groupe Hémisphères), as well as the participants of the Adopt a Lake campaign for their help in obtaining lake samples from Canada. We also thank the funders, staff, and students who have helped maintain the long-term eutrophication experiment in Lake 227 at the IISD Experimental Lakes Area. We acknowledge technical support from Thermo Fisher Scientific and Phytronix Technologies.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Costa, D.; Magalhães, J.D.; Cardoso, S.M.; Empadinhas, N. Microbial BMAA and the pathway for Parkinson’s disease neurodegeneration. Front. Aging Neurosci. 2020, 12, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, S.D.; Banack, S.A.; Cox, P.A.; Weiss, J.H. BMAA selectively injures motor neurons via AMPA/kainate receptor activation. Exp. Neurol. 2006, 201, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Lobner, D.; Piana, P.M.T.; Salous, A.K.; Peoples, R.W. β-N-methylamino-l-alanine enhances neurotoxicity through multiple mechanisms. Neurobiol. Dis. 2007, 25, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esterhuizen, M.; Downing, T.G. β-N-methylamino-l-alanine (BMAA) in novel South African cyanobacterial isolates. Ecotoxicol. Environ. Saf. 2008, 71, 309–313. [Google Scholar] [CrossRef]
- Banack, S.; Johnson, H.; Cheng, R.; Cox, P. Production of the neurotoxin BMAA by a marine cyanobacterium. Mar. Drugs 2007, 5, 180–196. [Google Scholar] [CrossRef]
- Snyder, L.R.; Cruz-Aguado, R.; Sadilek, M.; Galasko, D.; Shaw, C.A.; Montine, T.J. Parkinson–dementia complex and development of a new stable isotope dilution assay for BMAA detection in tissue. Toxicol. Appl. Pharmacol. 2009, 240, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Berntzon, L.; Ronnevi, L.O.; Bergman, B.; Eriksson, J. Detection of BMAA in the human central nervous system. Neuroscience 2015, 292, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Chang, A.C.G.; Chen, T.; Li, N.; Duan, J. Perspectives on endosymbiosis in coralloid roots: Association of cycads and cyanobacteria. Front. Microbiol. 2019, 10, 1888. [Google Scholar] [CrossRef] [Green Version]
- Lance, E.; Arnich, N.; Maignien, T.; Biré, R. Occurrence of β-N-methylamino-l-alanine (BMAA) and isomers in aquatic environments and aquatic food sources for humans. Toxins 2018, 10, 83. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Chen, Q.; Chen, X.; Wang, X.; Liao, X.; Jiang, L.; Wu, J.; Yang, L. Occurrence and transfer of a cyanobacterial neurotoxin β-methylamino-l-alanine within the aquatic food webs of Gonghu Bay (Lake Taihu, China) to evaluate the potential human health risk. Sci. Total Environ. 2014, 468–469, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Manolidi, K.; Triantis, T.M.; Kaloudis, T.; Hiskia, A. Neurotoxin BMAA and its isomeric amino acids in cyanobacteria and cyanobacteria-based food supplements. J. Hazard. Mater. 2019, 365, 346–365. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E. Presence of the neurotoxin BMAA in aquatic ecosystems: What do we really know? Toxins 2014, 6, 1109–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metcalf, J.S.; Banack, S.A.; Powell, J.T.; Tymm, F.J.M.; Murch, S.J.; Brand, L.E.; Cox, P.A. Public health responses to toxic cyanobacterial blooms: Perspectives from the 2016 Florida event. Water Policy 2018, 20, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Chatziefthimiou, A.D.; Deitch, E.J.; Glover, W.B.; Powell, J.T.; Banack, S.A.; Richer, R.A.; Cox, P.A.; Metcalf, J.S. Analysis of neurotoxic amino acids from marine waters, microbial mats, and seafood destined for human consumption in the Arabian Gulf. Neurotox. Res. 2018, 33, 143–152. [Google Scholar] [CrossRef]
- Bishop, S.L.; Kerkovius, J.K.; Menard, F.; Murch, S.J. N-β-Methylamino-L-Alanine and its naturally occurring isomers in cyanobacterial blooms in Lake Winnipeg. Neurotox. Res. 2018, 33, 133–142. [Google Scholar] [CrossRef]
- Pip, E.; Munford, K.; Bowman, L. Seasonal nearshore occurrence of the neurotoxin β N methylamino L alanine (BMAA) in Lake Winnipeg, Canada. Environ. Pollut. 2016, 5, 110. [Google Scholar] [CrossRef]
- Al-Sammak, M.; Hoagland, K.; Cassada, D.; Snow, D. Co-occurrence of the cyanotoxins BMAA, DABA and anatoxin-a in Nebraska reservoirs, fish, and aquatic plants. Toxins 2014, 6, 488–508. [Google Scholar] [CrossRef] [Green Version]
- Roy-Lachapelle, A.; Solliec, M.; Sauvé, S. Determination of BMAA and three alkaloid cyanotoxins in lake water using dansyl chloride derivatization and high-resolution mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 5487–5501. [Google Scholar] [CrossRef]
- Bishop, S.L.; Murch, S.J. A systematic review of analytical methods for the detection and quantification of β-N-methylamino-l-alanine (BMAA). Analyst 2020, 145, 13–28. [Google Scholar] [CrossRef]
- Cohen, S.A. Analytical techniques for the detection of α-amino-β-methylaminopropionic acid. Analyst 2012, 137, 1991. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.J.; Beekman, W.; Lürling, M. Evaluation of a commercial enzyme linked immunosorbent assay (ELISA) for the determination of the neurotoxin BMAA in surface waters. PLoS ONE 2013, 8, e65260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tymm, F.J.M.; Bishop, S.L.; Murch, S.J. A single laboratory validation for the analysis of underivatized β-N-Methylamino-L-Alanine (BMAA). Neurotox. Res. 2021, 39, 49–71. [Google Scholar] [CrossRef]
- Zhang, Y.; Husk, B.R.; Duy, S.V.; Dinh, Q.T.; Sanchez, J.S.; Sauvé, S.; Whalen, J.K. Quantitative screening for cyanotoxins in soil and groundwater of agricultural watersheds in Quebec, Canada. Chemosphere 2021, 274, 129781. [Google Scholar] [CrossRef]
- Bláhová, L.; Kohoutek, J.; Kadlecová, E.; Kozáková, L.; Bláha, L. Assessment of non-derivatized β-N-methylamino-l-alanine (BMAA) neurotoxin in free form in urine of patients with nonspecific neurological symptoms. Toxicon 2017, 133, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.; Antoniou, M.; Beekman-Lukassen, W.; Blahova, L.; Chernova, E.; Christophoridis, C.; Combes, A.; Edwards, C.; Fastner, J.; Harmsen, J.; et al. A collaborative evaluation of LC-MS/MS based methods for BMAA analysis: Soluble bound BMAA found to be an important fraction. Mar. Drugs 2016, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- Lage, S.; Burian, A.; Rasmussen, U.; Costa, P.R.; Annadotter, H.; Godhe, A.; Rydberg, S. BMAA extraction of cyanobacteria samples: Which method to choose? Environ. Sci. Pollut. Res. 2016, 23, 338–350. [Google Scholar] [CrossRef]
- Jiang, L.; Johnston, E.; Åberg, K.M.; Nilsson, U.; Ilag, L.L. Strategy for quantifying trace levels of BMAA in cyanobacteria by LC/MS/MS. Anal. Bioanal. Chem. 2013, 405, 1283–1292. [Google Scholar] [CrossRef]
- Vo Duy, S.; Munoz, G.; Dinh, Q.T.; Do, D.T.; Simon, D.F.; Sauvé, S. Analysis of the neurotoxin β-N-methylamino-L-alanine (BMAA) and isomers in surface water by FMOC derivatization liquid chromatography high resolution mass spectrometry. PLoS ONE 2019, 14, e0220698. [Google Scholar] [CrossRef] [Green Version]
- Foss, A.J.; Chernoff, N.; Aubel, M.T. The analysis of underivatized β-Methylamino-L-alanine (BMAA), BAMA, AEG & 2, 4-DAB in Pteropus mariannus mariannus specimens using HILIC-LC-MS/MS. Toxicon 2018, 152, 150–159. [Google Scholar]
- Baptista, M.S.; Cianca, R.C.C.; Lopes, V.R.; Almeida, C.M.R.; Vasconcelos, V.M. Determination of the non protein amino acid β-N-methylamino-l-alanine in estuarine cyanobacteria by capillary electrophoresis. Toxicon 2011, 58, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, M.; Pozo, Ó.J.; Sancho, J.V.; López, F.J.; Hernández, F. Re-evaluation of glyphosate determination in water by liquid chromatography coupled to electrospray tandem mass spectrometry. J. Chromatogr. A 2006, 1134, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Yvon, M.; Chabanet, C.; Pélissier, J.-P. Solubility of peptides in trichloroacetic acid (TCA) solutions Hypothesis on the precipitation mechanism. Int. J. Pept. Protein Res. 2009, 34, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Main, B.J.; Bowling, L.C.; Padula, M.P.; Bishop, D.P.; Mitrovic, S.M.; Guillemin, G.J.; Rodgers, K.J. Detection of the suspected neurotoxin β-methylamino-l-alanine (BMAA) in cyanobacterial blooms from multiple water bodies in Eastern Australia. Harmful Algae 2018, 74, 10–18. [Google Scholar] [CrossRef]
- Kehoe, M.J.; Chun, K.P.; Baulch, H.M. Who smells? Forecasting taste and odor in a drinking water reservoir. Environ. Sci. Technol. 2015, 49, 10984–10992. [Google Scholar] [CrossRef]
- Politi, E.; Prairie, Y.T. The potential of Earth Observation in modelling nutrient loading and water quality in lakes of southern Québec, Canada. Aquat. Sci. 2018, 80, 8. [Google Scholar] [CrossRef]
- Dillenberg, H.; Dehnel, M. Toxic waterbloom in Saskatchewan, 1959. Can. Med. Assoc. J. 1960, 83, 1151. [Google Scholar]
- Painter, K.J.; Venkiterwaran, J.J.; Simon, D.F.; Vo Duy, S.; Sauvé, S.; Baulch, H.M. Early and Late Bloomers in a Shallow, Eutrophic Lake. 2022. Available online: https://www.essoar.org/doi/abs/10.1002/essoar.10510183.1 (accessed on 6 January 2022).
- Ministère de l’Environnement et de la Lutte Contre les Changements Climatiques (MELCC). Réseau de Surveillance Volontaire des lacs. Suivi de la Qualité de l’eau 2016; Technical Report; Ministère de l’Environnement et de la Lutte Contre les Changements Climatiques: Quebec City, QC, Canada, 2016.
- Available online: https://www.associationdulactomcod.org/cyanobact%C3%A9ries.php (accessed on 6 January 2022).
- Violi, J.P.; Mitrovic, S.M.; Colville, A.; Main, B.J.; Rodgers, K.J. Prevalence of β-methylamino-L-alanine (BMAA) and its isomers in freshwater cyanobacteria isolated from eastern Australia. Ecotoxicol. Environ. Saf. 2019, 172, 72–81. [Google Scholar] [CrossRef]
- Weiss, J.H.; Christine, C.W.; Choi, D.W. Bicarbonate dependence of glutamate receptor activation by β-N-methylamino-L-alanine: Channel recording and study with related compounds. Neuron 1989, 3, 321–326. [Google Scholar] [CrossRef]
- Main, B.J.; Rodgers, K.J. Assessing the combined toxicity of BMAA and its isomers 2,4-DAB and AEG in vitro using human neuroblastoma cells. Neurotox. Res. 2018, 33, 33–42. [Google Scholar] [CrossRef]
- Metcalf, J.; Banack, S.; Richer, R.; Cox, P. Neurotoxic amino acids and their isomers in desert environments. J. Arid Environ. 2015, 112, 140–144. [Google Scholar] [CrossRef]
- Larsen, M.L.; Baulch, H.M.; Schiff, S.L.; Simon, D.F.; Sauvé, S.; Venkiteswaran, J.J.; Swart, N.C. Extreme rainfall drives early onset cyanobacterial bloom. FACETS 2020, 5, 899–920. [Google Scholar] [CrossRef]
- Giani, A.; Taranu, Z.E.; von Rückert, G.; Gregory-Eaves, I. Comparing key drivers of cyanobacteria biomass in temperate and tropical systems. Harmful Algae 2020, 97, 101859. [Google Scholar] [CrossRef]
- Tran Khac, V.; Hong, Y.; Plec, D.; Lemaire, B.; Dubois, P.; Saad, M.; Vinçon-Leite, B. An automatic monitoring system for high-frequency measuring and real-time management of cyanobacterial blooms in urban water bodies. Processes 2018, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Roy-Lachapelle, A.; Duy, S.V.; Munoz, G.; Dinh, Q.T.; Bahl, E.; Simon, D.F.; Sauvé, S. Analysis of multiclass cyanotoxins (microcystins, anabaenopeptins, cylindrospermopsin and anatoxins) in lake waters using on-line SPE liquid chromatography high-resolution Orbitrap mass spectrometry. Anal. Methods 2019, 11, 5289–5300. [Google Scholar] [CrossRef] [Green Version]
- Dinh, Q.T.; Munoz, G.; Simon, D.F.; Duy, S.V.; Husk, B.; Sauvé, S. Stability issues of microcystins, anabaenopeptins, anatoxins, and cylindrospermopsin during short-term and long-term storage of surface water and drinking water samples. Harmful Algae 2021, 101, 101955. [Google Scholar] [CrossRef] [PubMed]
- Munoz, G.; Duy, S.V.; Roy-Lachapelle, A.; Husk, B.; Sauvé, S. Analysis of individual and total microcystins in surface water by on-line preconcentration and desalting coupled to liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2017, 1516, 9–20. [Google Scholar] [CrossRef]
- Meriluoto, J.; Spoof, L.; Codd, G.A. Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Hoboken, NJ, USA, 2017. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 6 January 2022).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).