An Update on Clostridioides difficile Binary Toxin

 ,
, 
Abstract
:1. Introduction
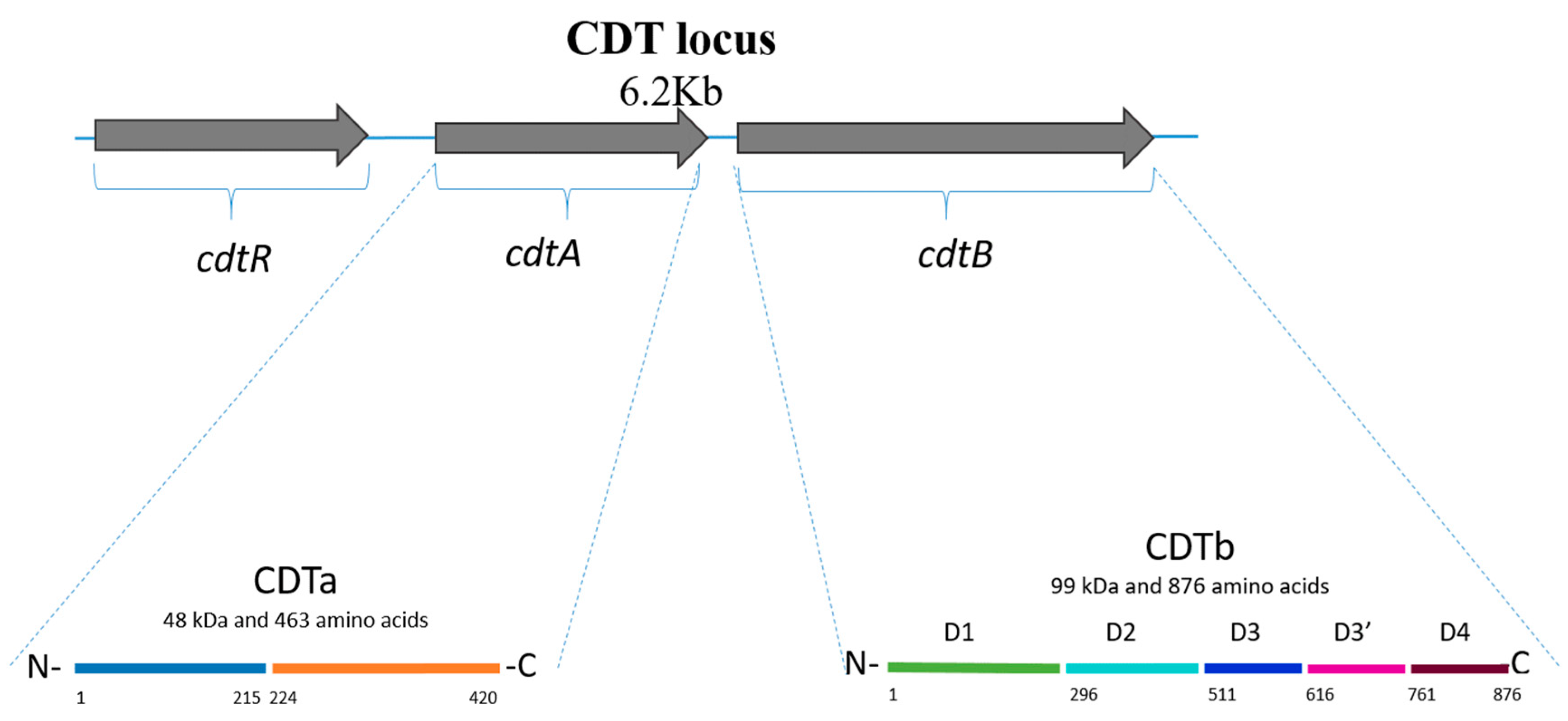
2. Clostridioides difficile Binary Toxin (CDT)
3. In Vitro Effects of CDT
4. CDT-Producing Ribotypes
5. Clinical Relevance
6. Implications of CDT in Laboratory Diagnosis
7. CDT as a Therapeutic Target
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Czepiel, J.; Drozdz, M.; Pituch, H.; Kuijper, E.J.; Perucki, W.; Mielimonka, A.; Goldman, S.; Wultanska, D.; Garlicki, A.; Biesiada, G. Clostridium difficile infection: Review. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Shoaei, P.; Shojaei, H.; Shirani, K. Phenotypic and Genotypic Characteristics of Clostridium difficile Isolates in Patients with Type 2 Diabetes in Iran. Infect. Drug Resist. 2020, 13, 683–690. [Google Scholar] [CrossRef] [Green Version]
- Abt, M.C.; McKenney, P.T.; Pamer, E.G. Clostridium difficile colitis: Pathogenesis and host defence. Nat. Rev. Microbiol. 2016, 14, 609–620. [Google Scholar] [CrossRef]
- Crobach, M.J.T.; Vernon, J.J.; Loo, V.G.; Kong, L.Y.; Péchiné, S.; Wilcox, M.H.; Kuijper, E.J. Understanding Clostridium difficile Colonization. Clin. Microbiol. Rev. 2018, 31, e00021-17. [Google Scholar] [CrossRef] [Green Version]
- Hung, Y.P.; Huang, I.H.; Lin, H.J.; Tsai, B.Y.; Liu, H.C.; Liu, H.C.; Lee, J.C.; Wu, Y.H.; Tsai, P.J.; Ko, W.C. Predominance of Clostridium difficile Ribotypes 017 and 078 among Toxigenic Clinical Isolates in Southern Taiwan. PLoS ONE 2016, 11, e0166159. [Google Scholar] [CrossRef] [Green Version]
- McDonald, L.C.; Killgore, G.E.; Thompson, A.; Owens, R.C., Jr.; Kazakova, S.V.; Sambol, S.P.; Johnson, S.; Gerding, D.N. An epidemic, toxin gene-variant strain of Clostridium difficile. N. Engl. J. Med. 2005, 353, 2433–2441. [Google Scholar] [CrossRef] [Green Version]
- Ghose, C. Clostridium difficile infection in the twenty-first century. Emerg. Microbes Infect. 2013, 2, e62. [Google Scholar] [CrossRef]
- Barbut, F.; Decre, D.; Lalande, V.; Burghoffer, B.; Noussair, L.; Gigandon, A.; Espinasse, F.; Raskine, L.; Robert, J.; Mangeol, A.; et al. Clinical features of Clostridium difficile-associated diarrhoea due to binary toxin (actin-specific ADP-ribosyltransferase)-producing strains. J. Med. Microbiol. 2005, 54, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Barth, H.; Aktories, K.; Popoff, M.R.; Stiles, B.G. Binary bacterial toxins: Biochemistry, biology, and applications of common Clostridium and Bacillus proteins. Microbiol. Mol. Biol. Rev. 2004, 68, 373–402. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, R.; Lacy, D.B. The role of toxins in Clostridium difficile infection. FEMS Microbiol. Rev. 2017, 41, 723–750. [Google Scholar] [CrossRef] [Green Version]
- Kordus, S.L.; Thomas, A.K.; Lacy, D.B. Clostridioides difficile toxins: Mechanisms of action and antitoxin therapeutics. Nat. Rev. Microbiol. 2021, 20, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Aktories, K.; Schwan, C.; Jank, T. Clostridium difficile Toxin Biology. Annu. Rev. Microbiol. 2017, 71, 281–307. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Lyras, D.; Allen, D.L.; Mackin, K.E.; Howarth, P.M.; O’Connor, J.R.; Rood, J.I. Binary toxin production in Clostridium difficile is regulated by CdtR, a LytTR family response regulator. J. Bacteriol. 2007, 189, 7290–7301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyon, S.A.; Hutton, M.L.; Rood, J.I.; Cheung, J.K.; Lyras, D. CdtR Regulates TcdA and TcdB Production in Clostridium difficile. PLoS Pathog. 2016, 12, e1005758. [Google Scholar] [CrossRef] [Green Version]
- Bilverstone, T.W.; Kinsmore, N.L.; Minton, N.P.; Kuehne, S.A. Development of Clostridium difficile R20291DeltaPaLoc model strains and in vitro methodologies reveals CdtR is required for the production of CDT to cytotoxic levels. Anaerobe 2017, 44, 51–54. [Google Scholar] [CrossRef]
- Bilverstone, T.W.; Minton, N.P.; Kuehne, S.A. Phosphorylation and functionality of CdtR in Clostridium difficile. Anaerobe 2019, 58, 103–109. [Google Scholar] [CrossRef]
- Gerding, D.N.; Johnson, S.; Rupnik, M.; Aktories, K. Clostridium difficile binary toxin CDT: Mechanism, epidemiology, and potential clinical importance. Gut Microbes 2014, 5, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.M.; Sheedlo, M.J.; Jensen, J.L.; Lacy, D.B. Structural insights into the transition of Clostridioides difficile binary toxin from prepore to pore. Nat. Microbiol. 2020, 5, 102–107. [Google Scholar] [CrossRef]
- Papatheodorou, P.; Carette, J.E.; Bell, G.W.; Schwan, C.; Guttenberg, G.; Brummelkamp, T.R.; Aktories, K. Lipolysis-stimulated lipoprotein receptor (LSR) is the host receptor for the binary toxin Clostridium difficile transferase (CDT). Proc. Natl. Acad. Sci. USA 2011, 108, 16422–16427. [Google Scholar] [CrossRef] [Green Version]
- Sheedlo, M.J.; Anderson, D.M.; Thomas, A.K.; Lacy, D.B. Structural elucidation of the Clostridioides difficile transferase toxin reveals a single-site binding mode for the enzyme. Proc. Natl. Acad. Sci. USA 2020, 117, 6139–6144. [Google Scholar] [CrossRef]
- Stieglitz, F.; Gerhard, R.; Pich, A. The Binary Toxin of Clostridioides difficile Alters the Proteome and Phosphoproteome of HEp-2 Cells. Front. Microbiol. 2021, 12, 725612. [Google Scholar] [CrossRef] [PubMed]
- Landenberger, M.; Nieland, J.; Roeder, M.; Norgaard, K.; Papatheodorou, P.; Ernst, K.; Barth, H. The cytotoxic effect of Clostridioides difficile pore-forming toxin CDTb. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183603. [Google Scholar] [CrossRef] [PubMed]
- Papatheodorou, P.; Hornuss, D.; Nolke, T.; Hemmasi, S.; Castonguay, J.; Picchianti, M.; Aktories, K. Clostridium difficile binary toxin CDT induces clustering of the lipolysis-stimulated lipoprotein receptor into lipid rafts. mBio 2013, 4, e00244-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwan, C.; Kruppke, A.S.; Nolke, T.; Schumacher, L.; Koch-Nolte, F.; Kudryashev, M.; Stahlberg, H.; Aktories, K. Clostridium difficile toxin CDT hijacks microtubule organization and reroutes vesicle traffic to increase pathogen adherence. Proc. Natl. Acad. Sci. USA 2014, 111, 2313–2318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwan, C.; Stecher, B.; Tzivelekidis, T.; van Ham, M.; Rohde, M.; Hardt, W.D.; Wehland, J.; Aktories, K. Clostridium difficile toxin CDT induces formation of microtubule-based protrusions and increases adherence of bacteria. PLoS Pathog. 2009, 5, e1000626. [Google Scholar] [CrossRef] [Green Version]
- Kuehne, S.A.; Collery, M.M.; Kelly, M.L.; Cartman, S.T.; Cockayne, A.; Minton, N.P. Importance of toxin A, toxin B, and CDT in virulence of an epidemic Clostridium difficile strain. J. Infect. Dis. 2014, 209, 83–86. [Google Scholar] [CrossRef]
- Carter, G.P.; Chakravorty, A.; Pham Nguyen, T.A.; Mileto, S.; Schreiber, F.; Li, L.; Howarth, P.; Clare, S.; Cunningham, B.; Sambol, S.P.; et al. Defining the Roles of TcdA and TcdB in Localized Gastrointestinal Disease, Systemic Organ Damage, and the Host Response during Clostridium difficile Infections. mBio 2015, 6, e00551. [Google Scholar] [CrossRef] [Green Version]
- Nibbering, B.; Gerding, D.N.; Kuijper, E.J.; Zwittink, R.D.; Smits, W.K. Host Immune Responses to Clostridioides difficile: Toxins and Beyond. Front. Microbiol. 2021, 12, 804949. [Google Scholar] [CrossRef]
- Cowardin, C.A.; Buonomo, E.L.; Saleh, M.M.; Wilson, M.G.; Burgess, S.L.; Kuehne, S.A.; Schwan, C.; Eichhoff, A.M.; Koch-Nolte, F.; Lyras, D.; et al. The binary toxin CDT enhances Clostridium difficile virulence by suppressing protective colonic eosinophilia. Nat. Microbiol. 2016, 1, 16108. [Google Scholar] [CrossRef] [Green Version]
- Marquardt, I.; Jakob, J.; Scheibel, J.; Hofmann, J.D.; Klawonn, F.; Neumann-Schaal, M.; Gerhard, R.; Bruder, D.; Jansch, L. Clostridioides difficile Toxin CDT Induces Cytotoxic Responses in Human Mucosal-Associated Invariant T (MAIT) Cells. Front. Microbiol. 2021, 12, 752549. [Google Scholar] [CrossRef]
- Knapp, O.; Benz, R.; Popoff, M.R. Pore-forming activity of clostridial binary toxins. Biochim. Biophys. Acta 2016, 1858, 512–525. [Google Scholar] [CrossRef]
- Labbe, A.C.; Poirier, L.; Maccannell, D.; Louie, T.; Savoie, M.; Beliveau, C.; Laverdiere, M.; Pepin, J. Clostridium difficile infections in a Canadian tertiary care hospital before and during a regional epidemic associated with the BI/NAP1/027 strain. Antimicrob. Agents Chemother. 2008, 52, 3180–3187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goorhuis, A.; Van der Kooi, T.; Vaessen, N.; Dekker, F.W.; Van den Berg, R.; Harmanus, C.; van den Hof, S.; Notermans, D.W.; Kuijper, E.J. Spread and epidemiology of Clostridium difficile polymerase chain reaction ribotype 027/toxinotype III in The Netherlands. Clin. Infect. Dis. 2007, 45, 695–703. [Google Scholar] [CrossRef] [Green Version]
- Warny, M.; Pepin, J.; Fang, A.; Killgore, G.; Thompson, A.; Brazier, J.; Frost, E.; McDonald, L.C. Toxin production by an emerging strain of Clostridium difficile associated with outbreaks of severe disease in North America and Europe. Lancet 2005, 366, 1079–1084. [Google Scholar] [CrossRef]
- Rao, K.; Micic, D.; Natarajan, M.; Winters, S.; Kiel, M.J.; Walk, S.T.; Santhosh, K.; Mogle, J.A.; Galecki, A.T.; LeBar, W.; et al. Clostridium difficile ribotype 027: Relationship to age, detectability of toxins A or B in stool with rapid testing, severe infection, and mortality. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2015, 61, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, K.; Higgins, P.D.R.; Young, V.B. An Observational Cohort Study of Clostridium difficile Ribotype 027 and Recurrent Infection. mSphere 2018, 3, e00033-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goorhuis, A.; Bakker, D.; Corver, J.; Debast, S.B.; Harmanus, C.; Notermans, D.W.; Bergwerff, A.A.; Dekker, F.W.; Kuijper, E.J. Emergence of Clostridium difficile infection due to a new hypervirulent strain, polymerase chain reaction ribotype 078. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2008, 47, 1162–1170. [Google Scholar] [CrossRef] [Green Version]
- Patterson, L.; Wilcox, M.H.; Fawley, W.N.; Verlander, N.Q.; Geoghegan, L.; Patel, B.C.; Wyatt, T.; Smyth, B. Morbidity and mortality associated with Clostridium difficile ribotype 078: A case-case study. J. Hosp. Infect. 2012, 82, 125–128. [Google Scholar] [CrossRef]
- Goorhuis, A.; Debast, S.B.; van Leengoed, L.A.; Harmanus, C.; Notermans, D.W.; Bergwerff, A.A.; Kuijper, E.J. Clostridium difficile PCR ribotype 078: An emerging strain in humans and in pigs? J. Clin. Microbiol. 2008, 46, 1157, author reply 1158. [Google Scholar] [CrossRef] [Green Version]
- Krutova, M.; Zouharova, M.; Matejkova, J.; Tkadlec, J.; Krejci, J.; Faldyna, M.; Nyc, O.; Bernardy, J. The emergence of Clostridium difficile PCR ribotype 078 in piglets in the Czech Republic clusters with Clostridium difficile PCR ribotype 078 isolates from Germany, Japan and Taiwan. Int. J. Med. Microbiol. 2018, 308, 770–775. [Google Scholar] [CrossRef]
- Kim, H.Y.; Cho, A.; Kim, J.W.; Kim, H.; Kim, B. High prevalence of Clostridium difficile PCR ribotype 078 in pigs in Korea. Anaerobe 2018, 51, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Shaw, H.A.; Preston, M.D.; Vendrik, K.E.W.; Cairns, M.D.; Browne, H.P.; Stabler, R.A.; Crobach, M.J.T.; Corver, J.; Pituch, H.; Ingebretsen, A.; et al. The recent emergence of a highly related virulent Clostridium difficile clade with unique characteristics. Clin. Microbiol. Infect. 2019, 26, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Eyre, D.W.; Tracey, L.; Elliott, B.; Slimings, C.; Huntington, P.G.; Stuart, R.L.; Korman, T.M.; Kotsiou, G.; McCann, R.; Griffiths, D.; et al. Emergence and spread of predominantly community-onset Clostridium difficile PCR ribotype 244 infection in Australia, 2010 to 2012. Eurosurveillance 2015, 20, 21059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida, M.N.; Heffernan, H.; Dervan, A.; Bakker, S.; Freeman, J.T.; Bhally, H.; Taylor, S.L.; Riley, T.V.; Roberts, S.A. Severe Clostridium difficile infection in New Zealand associated with an emerging strain, PCR-ribotype 244. N. Z. Med. J. 2013, 126, 9–14. [Google Scholar]
- Wiuff, C.; Brown, D.J.; Mather, H.; Banks, A.L.; Eastaway, A.; Coia, J.E. The epidemiology of Clostridium difficile in Scotland. J. Infect. 2011, 62, 271–279. [Google Scholar] [CrossRef]
- Tickler, I.A.; Goering, R.V.; Whitmore, J.D.; Lynn, A.N.; Persing, D.H.; Tenover, F.C.; Healthcare Associated Infection, C. Strain types and antimicrobial resistance patterns of Clostridium difficile isolates from the United States, 2011 to 2013. Antimicrob. Agents Chemother. 2014, 58, 4214–4218. [Google Scholar] [CrossRef] [Green Version]
- Davies, K.A.; Ashwin, H.; Longshaw, C.M.; Burns, D.A.; Davis, G.L.; Wilcox, M.H.; Group, E.S. Diversity of Clostridium difficile PCR ribotypes in Europe: Results from the European, multicentre, prospective, biannual, point-prevalence study of Clostridium difficile infection in hospitalised patients with diarrhoea (EUCLID), 2012 and 2013. Eurosurveillance 2016, 21, 30294. [Google Scholar] [CrossRef]
- Krutova, M.; Nyc, O.; Matejkova, J.; Allerberger, F.; Wilcox, M.H.; Kuijper, E.J. Molecular characterisation of Czech Clostridium difficile isolates collected in 2013–2015. Int. J. Med. Microbiol. 2016, 306, 479–485. [Google Scholar] [CrossRef]
- Dingle, K.E.; Griffiths, D.; Didelot, X.; Evans, J.; Vaughan, A.; Kachrimanidou, M.; Stoesser, N.; Jolley, K.A.; Golubchik, T.; Harding, R.M.; et al. Clinical Clostridium difficile: Clonality and pathogenicity locus diversity. PLoS ONE 2011, 6, e19993. [Google Scholar] [CrossRef]
- Janezic, S.; Rupnik, M. Genomic diversity of Clostridium difficile strains. Res. Microbiol. 2015, 166, 353–360. [Google Scholar] [CrossRef]
- Ziegler, M.; Landsburg, D.; Pegues, D.; Alby, K.; Gilmar, C.; Bink, K.; Gorman, T.; Moore, A.; Bonhomme, B.; Omorogbe, J.; et al. Clinical Characteristics and Outcomes of Hematologic Malignancy Patients with Positive Clostridium difficile Toxin Immunoassay Versus Polymerase Chain Reaction Test Results. Infect. Control. Hosp. Epidemiol. 2018, 39, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Ortiz, A.; Lopez-Barrera, D.; Hernandez-Garcia, R.; Galvan-De Los Santos, A.M.; Flores-Trevino, S.M.; Llaca-Diaz, J.M.; Maldonado-Garza, H.J.; Bosques-Padilla, F.J.; Garza-Gonzalez, E. First report of Clostridium difficile NAP1/027 in a Mexican hospital. PLoS ONE 2015, 10, e0122627. [Google Scholar] [CrossRef]
- Jones, A.M.; Kuijper, E.J.; Wilcox, M.H. Clostridium difficile: A European perspective. J. Infect. 2013, 66, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Urena, D.; Quesada-Gomez, C.; Miranda, E.; Fonseca, M.; Rodriguez-Cavallini, E. Spread of epidemic Clostridium difficile NAP1/027 in Latin America: Case reports in Panama. J. Med. Microbiol. 2014, 63, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Gomez, C.; Rodriguez, C.; Gamboa-Coronado Mdel, M.; Rodriguez-Cavallini, E.; Du, T.; Mulvey, M.R.; Villalobos-Zuniga, M.; Boza-Cordero, R. Emergence of Clostridium difficile NAP1 in Latin America. J. Clin. Microbiol. 2010, 48, 669–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Androga, G.O.; Hart, J.; Foster, N.F.; Charles, A.; Forbes, D.; Riley, T.V. Infection with Toxin A-Negative, Toxin B-Negative, Binary Toxin-Positive Clostridium difficile in a Young Patient with Ulcerative Colitis. J. Clin. Microbiol. 2015, 53, 3702–3704. [Google Scholar] [CrossRef] [Green Version]
- Knight, D.R.; Riley, T.V. Clostridium difficile clade 5 in Australia: Antimicrobial susceptibility profiling of PCR ribotypes of human and animal origin. J. Antimicrob. Chemother. 2016, 71, 2213–2217. [Google Scholar] [CrossRef] [Green Version]
- Hung, Y.P.; Tsai, P.J.; Lee, Y.T.; Tang, H.J.; Lin, H.J.; Liu, H.C.; Lee, J.C.; Tsai, B.Y.; Hsueh, P.R.; Ko, W.C. Nationwide surveillance of ribotypes and antimicrobial susceptibilities of toxigenic Clostridium difficile isolates with an emphasis on reduced doxycycline and tigecycline susceptibilities among ribotype 078 lineage isolates in Taiwan. Infect. Drug Resist. 2018, 11, 1197–1203. [Google Scholar] [CrossRef] [Green Version]
- Zaiss, N.H.; Witte, W.; Nubel, U. Fluoroquinolone resistance and Clostridium difficile, Germany. Emerg. Infect. Dis. 2010, 16, 675–677. [Google Scholar] [CrossRef]
- Cassir, N.; Fahsi, N.; Durand, G.; Lagier, J.C.; Raoult, D.; Fournier, P.E. Emergence of Clostridium difficile tcdC variant 078 in Marseille, France. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1971–1974. [Google Scholar] [CrossRef] [Green Version]
- Usui, M.; Nanbu, Y.; Oka, K.; Takahashi, M.; Inamatsu, T.; Asai, T.; Kamiya, S.; Tamura, Y. Genetic relatedness between Japanese and European isolates of Clostridium difficile originating from piglets and their risk associated with human health. Front. Microbiol. 2014, 5, 513. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Liu, S.; Meng, X.; Liang, W.; Xu, Z.; Tang, B.; Wang, Y.; Duan, J.; Fu, C.; Wu, B.; et al. Genome characterization of a novel binary toxin-positive strain of Clostridium difficile and comparison with the epidemic 027 and 078 strains. Gut Pathog. 2017, 9, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehrhahn, M.C.; Keighley, C.; Kurtovic, J.; Knight, D.R.; Hong, S.; Hutton, M.L.; Lyras, D.; Wang, Q.; Leong, R.; Borody, T.; et al. A series of three cases of severe Clostridium difficile infection in Australia associated with a binary toxin producing clade 2 ribotype 251 strain. Anaerobe 2019, 55, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Crobach, M.J.T.; Voor In ’t Holt, A.F.; Knetsch, C.W.; van Dorp, S.M.; Bras, W.; Harmanus, C.; Kuijper, E.J.; Vos, M.C. An outbreak of Clostridium difficile infections due to new PCR ribotype 826: Epidemiologic and microbiologic analyses. Clin. Microbiol. Infect. 2018, 24, 309.e1–309.e4. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Liu, S.; Zhou, P.; Duan, J.; Dou, Q.; Zhang, R.; Chen, H.; Cheng, Y.; Wu, A. Emergence of a Novel Binary Toxin-Positive Strain of Clostridium difficile Associated with Severe Diarrhea That Was Not Ribotype 027 and 078 in China. Infect. Control Hosp. Epidemiol. 2015, 36, 1112–1114. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Harmanus, C.; Zhu, D.; Meng, X.; Wang, S.; Duan, J.; Liu, S.; Fu, C.; Zhou, P.; Liu, R.; et al. Characterization of the virulence of a non-RT027, non-RT078 and binary toxin-positive Clostridium difficile strain associated with severe diarrhea. Emerg. Microbes Infect. 2018, 7, 211. [Google Scholar] [CrossRef] [Green Version]
- Riedel, T.; Neumann-Schaal, M.; Wittmann, J.; Schober, I.; Hofmann, J.D.; Lu, C.W.; Dannheim, A.; Zimmermann, O.; Lochner, M.; Gross, U.; et al. Characterization of Clostridioides difficile DSM 101085 with A-B-CDT+ Phenotype from a Late Recurrent Colonization. Genome Biol. Evol. 2020, 12, 566–577. [Google Scholar] [CrossRef] [Green Version]
- Androga, G.O.; Knight, D.R.; Lim, S.C.; Foster, N.F.; Riley, T.V. Antimicrobial resistance in large clostridial toxin-negative, binary toxin-positive Clostridium difficile ribotypes. Anaerobe 2018, 54, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Androga, G.O.; Knight, D.R.; Hutton, M.L.; Mileto, S.J.; James, M.L.; Evans, C.; Lyras, D.; Chang, B.J.; Foster, N.F.; Riley, T.V. In silico, in vitro and in vivo analysis of putative virulence factors identified in large clostridial toxin-negative, binary toxin-producing C. difficile strains. Anaerobe 2019, 60, 102083. [Google Scholar] [CrossRef]
- Carman, R.J.; Stevens, A.L.; Lyerly, M.W.; Hiltonsmith, M.F.; Stiles, B.G.; Wilkins, T.D. Clostridium difficile binary toxin (CDT) and diarrhea. Anaerobe 2011, 17, 161–165. [Google Scholar] [CrossRef]
- Stewart, D.B.; Berg, A.; Hegarty, J. Predicting recurrence of C. difficile colitis using bacterial virulence factors: Binary toxin is the key. J. Gastrointest. Surg. 2013, 17, 118–124, discussion p 124-5. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.B.; Berg, A.S.; Hegarty, J.P. Single nucleotide polymorphisms of the tcdC gene and presence of the binary toxin gene predict recurrent episodes of Clostridium difficile infection. Ann. Surg. 2014, 260, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Bacci, S.; Molbak, K.; Kjeldsen, M.K.; Olsen, K.E. Binary toxin and death after Clostridium difficile infection. Emerg. Infect. Dis. 2011, 17, 976–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Cárdenas, S.; Torres-Martos, E.; Mora-Delgado, J.; Sánchez-Calvo, J.M.; Santos-Peña, M.; Zapata López, Á.; Dolores López-Prieto, M.; Pérez-Cortés, S.; Carlos Alados, J. The prognostic value of toxin B and binary toxin in Clostridioides difficile infection. Gut Microbes 2021, 13, 1884516. [Google Scholar] [CrossRef]
- Carlson, T.J.; Endres, B.T.; Le Pham, J.; Gonzales-Luna, A.J.; Alnezary, F.S.; Nebo, K.; Miranda, J.; Lancaster, C.; Basseres, E.; Begum, K.; et al. Eosinopenia and Binary Toxin Increase Mortality in Hospitalized Patients with Clostridioides difficile Infection. Open Forum Infect. Dis. 2020, 7, ofz552. [Google Scholar] [CrossRef]
- Goldenberg, S.D.; French, G.L. Lack of association of tcdC type and binary toxin status with disease severity and outcome in toxigenic Clostridium difficile. J. Infect. 2011, 62, 355–362. [Google Scholar] [CrossRef]
- Hensgens, M.P.; Kuijper, E.J. Clostridium difficile infection caused by binary toxin-positive strains. Emerg. Infect. Dis. 2013, 19, 1539–1540. [Google Scholar] [CrossRef]
- Walk, S.T.; Micic, D.; Jain, R.; Lo, E.S.; Trivedi, I.; Liu, E.W.; Almassalha, L.M.; Ewing, S.A.; Ring, C.; Galecki, A.T.; et al. Clostridium difficile ribotype does not predict severe infection. Clin. Infect. Dis. 2012, 55, 1661–1668. [Google Scholar] [CrossRef]
- Pilate, T.; Verhaegen, J.; Van Ranst, M.; Saegeman, V. Binary toxin and its clinical importance in Clostridium difficile infection, Belgium. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1741–1747. [Google Scholar] [CrossRef]
- Berry, C.E.; Davies, K.A.; Owens, D.W.; Wilcox, M.H. Is there a relationship between the presence of the binary toxin genes in Clostridium difficile strains and the severity of C. difficile infection (CDI)? Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2405–2415. [Google Scholar] [CrossRef]
- Reigadas, E.; Alcala, L.; Marin, M.; Martin, A.; Iglesias, C.; Bouza, E. Role of binary toxin in the outcome of Clostridium difficile infection in a non-027 ribotype setting. Epidemiol. Infect. 2016, 144, 268–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crobach, M.J.; Planche, T.; Eckert, C.; Barbut, F.; Terveer, E.M.; Dekkers, O.M.; Wilcox, M.H.; Kuijper, E.J. European Society of Clinical Microbiology and Infectious Diseases: Update of the diagnostic guidance document for Clostridium difficile infection. Clin. Microbiol. Infect. 2016, 22, S63–S81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; Coffin, S.E.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical Practice Guidelines for Clostridium difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, e1–e48. [Google Scholar] [CrossRef] [PubMed]
- Simor, A.E. Diagnosis, management, and prevention of Clostridium difficile infection in long-term care facilities: A review. J. Am. Geriatr. Soc. 2010, 58, 1556–1564. [Google Scholar] [CrossRef]
- Bartlett, J.G. Detection of Clostridium difficile infection. Infect. Control Hosp. Epidemiol. 2010, 31, S35–S37. [Google Scholar] [CrossRef]
- Camargo, T.S.; Junior, M.S.; Camargo, L.F.A.; Biotto, V.P.; Doi, A.M.; Koga, P.C.M.; Franca, C.N.; Martino, M.D.V. Clostridioides difficile laboratory diagnostic techniques: A comparative approach of rapid and molecular methods. Arch. Microbiol. 2021, 203, 1683–1690. [Google Scholar] [CrossRef]
- Garimella, P.S.; Agarwal, R.; Katz, A. The utility of repeat enzyme immunoassay testing for the diagnosis of Clostridium difficile infection: A systematic review of the literature. J. Postgrad. Med. 2012, 58, 194–198. [Google Scholar] [CrossRef]
- Brecher, S.M.; Novak-Weekley, S.M.; Nagy, E. Laboratory diagnosis of Clostridium difficile infections: There is light at the end of the colon. Clin. Infect. Dis. 2013, 57, 1175–1181. [Google Scholar] [CrossRef]
- Norén, T.; Alriksson, I.; Andersson, J.; Akerlund, T.; Unemo, M. Rapid and sensitive loop-mediated isothermal amplification test for Clostridium difficile detection challenges cytotoxin B cell test and culture as gold standard. J. Clin. Microbiol. 2011, 49, 710–711. [Google Scholar] [CrossRef] [Green Version]
- King, A.M.; Mackin, K.E.; Lyras, D. Emergence of toxin A-negative, toxin B-positive Clostridium difficile strains: Epidemiological and clinical considerations. Future Microbiol. 2015, 10, 1–4. [Google Scholar] [CrossRef]
- Abeyawardhane, D.L.; Godoy-Ruiz, R.; Adipietro, K.A.; Varney, K.M.; Rustandi, R.R.; Pozharski, E.; Weber, D.J. The Importance of Therapeutically Targeting the Binary Toxin from Clostridioides difficile. Int. J. Mol. Sci. 2021, 22, 2926. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Li, H.; Zhao, X.; Wang, X.; Wei, X.; Lin, W.; Li, P.; Cui, L.; Yuan, J. Rapid visual detection of binary toxin producing Clostridium difficile by loop-mediated isothermal amplification. Exp. Ther. Med. 2017, 14, 4781–4788. [Google Scholar] [CrossRef] [PubMed]
- Bagdasarian, N.; Rao, K.; Malani, P.N. Diagnosis and treatment of Clostridium difficile in adults: A systematic review. JAMA 2015, 313, 398–408. [Google Scholar] [CrossRef] [PubMed]
- McGovern, A.M.; Androga, G.O.; Moono, P.; Collins, D.A.; Foster, N.F.; Chang, B.J.; Riley, T.V. Evaluation of the Cepheid((R)) Xpert((R)) C. difficile binary toxin (BT) diagnostic assay. Anaerobe 2018, 51, 12–16. [Google Scholar] [CrossRef]
- Androga, G.O.; McGovern, A.M.; Elliott, B.; Chang, B.J.; Perkins, T.T.; Foster, N.F.; Riley, T.V. Evaluation of the Cepheid Xpert C. difficile/Epi and meridian bioscience illumigene C. difficile assays for detecting Clostridium difficile ribotype 033 strains. J. Clin. Microbiol. 2015, 53, 973–975. [Google Scholar] [CrossRef] [Green Version]
- Tojo, M.; Nagamatsu, M.; Hayakawa, K.; Mezaki, K.; Kirikae, T.; Ohmagari, N. Evaluation of an automated rapid diagnostic test for detection of Clostridium difficile. PLoS ONE 2014, 9, e106102. [Google Scholar] [CrossRef] [Green Version]
- Gateau, C.; Couturier, J.; Coia, J.; Barbut, F. How to: Diagnose infection caused by Clostridium difficile. Clin. Microbiol. Infect. 2018, 24, 463–468. [Google Scholar] [CrossRef] [Green Version]
- Ernst, K.; Landenberger, M.; Nieland, J.; Norgaard, K.; Frick, M.; Fois, G.; Benz, R.; Barth, H. Characterization and Pharmacological Inhibition of the Pore-Forming Clostridioides difficile CDTb Toxin. Toxins 2021, 13, 390. [Google Scholar] [CrossRef]
- Korbmacher, M.; Fischer, S.; Landenberger, M.; Papatheodorou, P.; Aktories, K.; Barth, H. Human alpha-Defensin-5 Efficiently Neutralizes Clostridioides difficile Toxins TcdA, TcdB, and CDT. Front. Pharmacol. 2020, 11, 1204. [Google Scholar] [CrossRef]
- Fischer, S.; Uckert, A.K.; Landenberger, M.; Papatheodorou, P.; Hoffmann-Richter, C.; Mittler, A.K.; Ziener, U.; Hagele, M.; Schwan, C.; Muller, M.; et al. Human peptide alpha-defensin-1 interferes with Clostridioides difficile toxins TcdA, TcdB, and CDT. FASEB J. 2020, 34, 6244–6261. [Google Scholar] [CrossRef] [Green Version]
- Ernst, K.; Schmid, J.; Beck, M.; Hagele, M.; Hohwieler, M.; Hauff, P.; Uckert, A.K.; Anastasia, A.; Fauler, M.; Jank, T.; et al. Hsp70 facilitates trans-membrane transport of bacterial ADP-ribosylating toxins into the cytosol of mammalian cells. Sci. Rep. 2017, 7, 2724. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, E.; Kroll, C.; Ernst, K.; Schwan, C.; Popoff, M.; Fischer, G.; Buchner, J.; Aktories, K.; Barth, H. Membrane translocation of binary actin-ADP-ribosylating toxins from Clostridium difficile and Clostridium perfringens is facilitated by cyclophilin A and Hsp90. Infect. Immun. 2011, 79, 3913–3921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, E.; Bohm, N.; Ernst, K.; Langer, S.; Schwan, C.; Aktories, K.; Popoff, M.; Fischer, G.; Barth, H. FK506-binding protein 51 interacts with Clostridium botulinum C2 toxin and FK506 inhibits membrane translocation of the toxin in mammalian cells. Cell Microbiol. 2012, 14, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Ernst, K.; Sailer, J.; Braune, M.; Barth, H. Intoxication of mammalian cells with binary clostridial enterotoxins is inhibited by the combination of pharmacological chaperone inhibitors. Naunyn Schmiedeberg’s Arch. Pharmacol. 2021, 394, 941–954. [Google Scholar] [CrossRef]
- Secore, S.; Wang, S.; Doughtry, J.; Xie, J.; Miezeiewski, M.; Rustandi, R.R.; Horton, M.; Xoconostle, R.; Wang, B.; Lancaster, C.; et al. Development of a Novel Vaccine Containing Binary Toxin for the Prevention of Clostridium difficile Disease with Enhanced Efficacy against NAP1 Strains. PLoS ONE 2017, 12, e0170640. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| RT | Toxin Genotype | ST | Clade | Characteristics | References |
|---|---|---|---|---|---|
| 023 | tcdA+, tcdB+, cdtA+, cdtB+ | 5, 22, 25 | 3 | Resistance to erythromycin, levofloxacin, and moxifloxacin. Reports from USA, Northern and Eastern Europe. | [45,46,47] |
| 027/176 | tcdA+, tcdB+, cdtA+, cdtB+ | 1 | 2 | Strain associated with increased morbidity and mortality. Reports from Korea, Singapore, Austria, Belgium, Denmark, Finland, France, Germany, Hungary, Ireland, Luxembourg, The Netherlands, Norway, Spain, Sweden, UK, Chile, Panama, Costa Rica, Mexico, Japan, China. | [33,48,49,50,51,52,53,54,55] |
| 033 | tcdA−, tcdB−, cdtA+, cdtB+ | ND | 5 | Isolated from a young patient with ulcerative colitis and severe diarrhea in Australia. | [56,57] |
| 078/126 | tcdA+, tcdB+, cdtA+, cdtB+ | 11 | 5 | Community-associated and zoonotic strain with increased morbidity and mortality. Reports from France, Italy, Germany, Taiwan, Czech Republic, Korea, Japan, Australia. | [5,40,41,48,50,57,58,59,60,61] |
| 244 | tcdA+, tcdB+, cdtA+, cdtB+ | 41 | 2 | Community-associated; cause of outbreaks. Reports in Australia, New Zealand. | [44,45,62] |
| 251 | tcdA+, tcdB+, cdtA+, cdtB+ | 231 | 2 | Isolated from three patients in Australia with severe diarrhea, recurrent disease, and one death. | [63] |
| 826 | tcdA+, tcdB+, cdtA+, cdtB+ | ND | 5 | Identified in an outbreak in The Netherlands, associated with recurrent and severe disease in two of five patients | [64] |
| ND | tcdA+, tcdB+, cdtA+, cdtB+ | 201 | 3 | Isolated from a patient in China, with a severe clinical phenotype; it exhibits a faster germination rate, higher motility, and a higher biofilm formation than RT027 and RT078. | [62,65,66] |
| ND | tcdA−, tcdB−, cdtA+, cdtB+ | 11 | 5 | Isolated from a patient in Germany, with eight episodes of CDI ranging from mild to severe symptoms. | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Meléndez, A.; Cruz-López, F.; Morfin-Otero, R.; Maldonado-Garza, H.J.; Garza-González, E. An Update on Clostridioides difficile Binary Toxin. Toxins 2022, 14, 305. https://doi.org/10.3390/toxins14050305
Martínez-Meléndez A, Cruz-López F, Morfin-Otero R, Maldonado-Garza HJ, Garza-González E. An Update on Clostridioides difficile Binary Toxin. Toxins. 2022; 14(5):305. https://doi.org/10.3390/toxins14050305
Chicago/Turabian StyleMartínez-Meléndez, Adrián, Flora Cruz-López, Rayo Morfin-Otero, Héctor J. Maldonado-Garza, and Elvira Garza-González. 2022. "An Update on Clostridioides difficile Binary Toxin" Toxins 14, no. 5: 305. https://doi.org/10.3390/toxins14050305
APA StyleMartínez-Meléndez, A., Cruz-López, F., Morfin-Otero, R., Maldonado-Garza, H. J., & Garza-González, E. (2022). An Update on Clostridioides difficile Binary Toxin. Toxins, 14(5), 305. https://doi.org/10.3390/toxins14050305