Mpp23Aa/Xpp37Aa Insecticidal Proteins from Bacillus thuringiensis (Bacillales: Bacillaceae) Are Highly Toxic to Anthonomus grandis (Coleoptera: Curculionidae) Larvae

 , , ,
, , ,  and
and 
Abstract
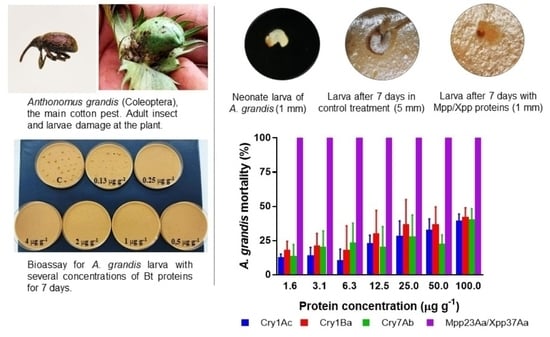
:
1. Introduction
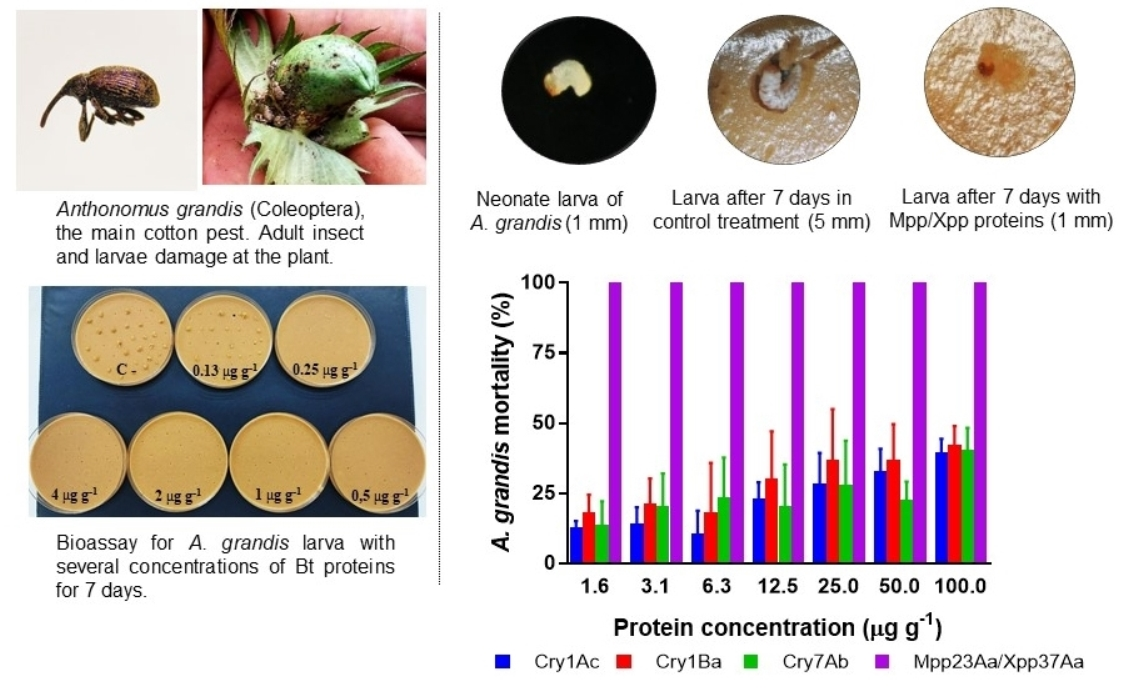
2. Results
3. Discussion
4. Materials and Methods
4.1. Bt Proteins Preparation
4.2. Rearing of Anthonomus grandis
4.3. Bioassays of A. grandis with Bt Proteins
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CONAB. Acompanhamento da Safra Brasileira de Grãos, Safra 2021/22. 2022. Available online: https://www.conab.gov.br/info-agro/safras/graos/boletim-da-safra-de-graos (accessed on 12 December 2022).
- USDA. Cotton: World Markets and Trade. 2022. Available online: https://www.fas.usda.gov/data/cotton-world-markets-and-trade (accessed on 12 December 2022).
- IMEA. Custo de Produção do Algodão, Safra 2020/2021. 2022. Available online: https://www.imea.com.br/imea-site/relatorios-mercado (accessed on 12 December 2022).
- Arruda, L.S.; Torres, J.B.; Rolim, G.G.; Silva-Torres, C.S.A. Dispersal of boll weevil toward and within the cotton plant and implications for insecticide exposure. Pest Manag. Sci. 2020, 77, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Grigolli, J.F.J.; Souza, L.A.; Fernandes, M.G.; Busoli, A.C. Spatial distribution of adult Anthonomus grandis Boheman (Coleoptera: Curculionidae) and damage to cotton flower buds due to feeding and oviposition. Neotrop. Entomol. 2017, 46, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Neves, R.C.S.; Showler, A.T.; Pinto, E.S.; Bastos, C.S.; Torres, J.B. Reducing boll weevil populations by clipping terminal buds and removing abscised fruiting bodies. Entomol. Exp. Appl. 2012, 146, 276–285. [Google Scholar] [CrossRef]
- Perkin, L.C.; Perez, J.L.; Suh, C.P.-C. The identification of boll weevil, Anthonomus grandis grandis (Coleoptera: Curculionidae), genes involved in pheromone production and pheromone biosynthesis. Insects 2021, 12, 893. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.A.S.; Araújo, T.A.; Showler, A.T.; Araújo, A.C.A.; Almeida, I.S.; Aguiar, R.S.A.; Miranda, J.E.; Fernandes, F.L.; Bastos, C.S. Spatio-temporal distribution of Anthonomus grandis Boh. in tropical cotton fields. Pest Manag. Sci. 2022, 78, 2492–2501. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.B.; Rolim, G.G.; Arruda, L.S.; Dos Santos, M.P.; Leite, S.A.; Neves, R.C.D.S. Insecticides in use and risk of control failure of boll weevil (Coleoptera: Curculionidae) in the Brazilian Cerrado. Neotrop. Entomol. 2022, 51, 613–627. [Google Scholar] [CrossRef]
- Bravo, A.; Gómez, I.; Porta, H.; García-Gómez, B.I.; Rodriguez-Almazan, C.; Pardo, L.; Soberón, M. Evolution of Bacillus thuringiensis Cry toxins insecticidal activity. Microb. Biotechnol. 2012, 6, 17–26. [Google Scholar] [CrossRef]
- Ribeiro, T.P.; Arraes, F.B.M.; Lourenço-Tessutti, I.T.; Silva, M.S.; Lisei-de-Sá, M.E.; Lucena, W.A.; Macedo, L.L.P.; Lima, J.N.; Santos Amorim, R.M.; Artico, S.; et al. Transgenic cotton expressing Cry10Aa toxin confers high resistance to the cotton boll weevil. Plant Biotechnol. J. 2017, 15, 997–1009. [Google Scholar] [CrossRef] [Green Version]
- ISAAA. GM Approval Database. 2022. Available online: https://www.isaaa.org/gmapprovaldatabase (accessed on 12 December 2022).
- Ribeiro, T.P.; Basso, M.F.; Carvalho, M.H.d.; Macedo, L.L.P.d.; Silva, D.M.L.d.; Lourenço-Tessutti, I.T.; Oliveira-Neto, O.B.d.; Campos-Pinto, E.R.d.; Lucena, W.A.; Silva, M.C.M.d.; et al. Stability and tissue-specific Cry10Aa overexpression improves cotton resistance to the cotton boll weevil. Biotechnol. Res. Innov. 2019, 3, 27–41. [Google Scholar] [CrossRef]
- Domínguez-Arrizabalaga, M.; Villanueva, M.; Escriche, B.; Ancín-Azpilicueta, C.; Caballero, P. Insecticidal activity of Bacillus thuringiensis proteins against Coleopteran Pests. Toxins 2020, 12, 430. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of on insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Martins, E.S.; Monnerat, R.G.; Queiroz, P.R.; Dumas, V.F.; Braz, S.V.; Aguiar, R.W.S.; Gomes, A.C.M.M.; Sánchez, J.; Bravo, A.; Ribeiro, B.M. Midgut GPI-anchored proteins with alkaline phosphatase activity from the cotton boll weevil (A. grandis) are putative receptors for the Cry1B protein of Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2010, 40, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Farias, D.F.; Peijnenburg, A.A.; Grossi-de-Sa, M.F.; Carvalho, A.F. Food safety knowledge on the Bt mutant protein Cry8Ka5 employed in the development of coleopteran-resistant transgenic cotton plants. Bioengineered 2015, 6, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Martins, E.S.; Aguiar, R.W.; Martins, N.F.; Melatti, V.M.; Falcao, R.; Gomes, A.C.; Ribeiro, B.M.; Monnerat, R.G. Recombinant Cry1Ia protein is highly toxic to cotton boll weevil (Anthonomus grandis Boheman) and fall armyworm (Spodoptera frugiperda). J. App. Microbiol. 2008, 104, 1363–1371. [Google Scholar] [CrossRef]
- Silva, C.R.; Monnerat, R.; Lima, L.M.; Martins, É.S.; Melo Filho, P.A.; Pinheiro, M.P.; Santos, R.C. Stable integration and expression of a cry1Ia gene conferring resistance to fall armyworm and boll weevil in cotton plants. Pest Manag. Sci. 2016, 72, 1549–1557. [Google Scholar] [CrossRef]
- de Oliveira, R.S.; Oliveira-Neto, O.B.; Moura, H.F.; de Macedo, L.L.; Arraes, F.B.; Lucena, W.A.; Lourenco-Tessutti, I.T.; de Deus Barbosa, A.A.; da Silva, M.C.; Grossi-de-Sa, M.F. Transgenic cotton plants expressing Cry1Ia12 toxin confer resistance to fall armyworm (Spodoptera frugiperda) and cotton boll weevil (Anthonomus grandis). Front. Plant Sci. 2016, 7, 165. [Google Scholar] [CrossRef] [Green Version]
- Grossi-de-Sa, M.F.; Quezado de Magalhaes, M.; Silva, M.S.; Silva, S.M.; Dias, S.C.; Nakasu, E.Y.; Brunetta, P.S.; Oliveira, G.R.; Neto, O.B.; Sampaio de Oliveira, R.; et al. Susceptibility of Anthonomus grandis (cotton boll weevil) and Spodoptera frugiperda (fall armyworm) to a cry1ia-type toxin from a Brazilian Bacillus thuringiensis strain. J. Biochem. Mol. Biol. 2007, 40, 773–782. [Google Scholar] [CrossRef]
- Aguiar, R.W.d.S.; Martins, E.S.; Ribeiro, B.M.; Monnerat, R.G. Cry10Aa protein is highly toxic to Anthonomus grandis Boheman (Coleoptera: Curculionidae), an important insect pest in Brazilian cotton crop fields. Bt. Res. 2012, 3, 20–28. [Google Scholar] [CrossRef]
- Jakka, S.; Ferré, J.; Jurat-Fuentes, J.L. Cry toxin binding site models and their use in strategies to delay resistance evolution. In Bt Resistance: Characterization and Strategies for GM Crops Producing Bacillus Thuringiensis Toxins; Soberón, M., Gao, Y., Bravo, A., Eds.; Oxfordshire, UK, 2015; pp. 138–149. [Google Scholar]
- Abdelgaffar, H.M.; Oppert, C.; Sun, X.; Monserrate, J.; Jurat-Fuentes, J.L. Differential heliothine susceptibility to Cry1Ac associated with gut proteolytic activity. Pestic. Biochem. Physiol. 2019, 153, 1–8. [Google Scholar] [CrossRef]
- Wang, P.; Ma, J.; Head, G.P.; Xia, D.; Li, J.; Wang, H.; Yang, M.; Xie, Z.; Zalucki, M.P.; Lu, Z. Susceptibility of Helicoverpa armigera to two Bt toxins, Cry1Ac and Cry2Ab, in northwestern China: Toward developing an IRM strategy. J. Pest. Sci. 2018, 92, 923–931. [Google Scholar] [CrossRef]
- Bel, Y.; Sheets, J.J.; Tan, S.Y.; Narva, K.E.; Escriche, B. Toxicity and binding studies of Bacillus thuringiensis Cry1Ac, Cry1F, Cry1C, and Cry2A proteins in the soybean pests Anticarsia gemmatalis and Chrysodeixis (Pseudoplusia) includens. Appl. Environ. Microbiol. 2017, 83, e00326-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, G.; Lu, J.; Zhang, P.; Li, J.; Zhu, F.; Chen, J.; Liu, Y.; Yu, Z.; Zhao, X. The cry1Ac gene of Bacillus thuringiensis ZQ-89 encodes a toxin against long-horned beetle adult. J. Appl. Microbiol. 2011, 110, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Paritosh, K.; Kant, U.P.; Burma, K.; Pental, D. High expression of Cry1Ac protein in cotton (Gossypium hirsutum) by combining independent transgenic events that target the protein to cytoplasm and plastids. PLoS ONE 2016, 11, e0158603. [Google Scholar] [CrossRef] [Green Version]
- Dhanaraj, A.L.; Willse, A.R.; Kamath, S.P. Stability of expression of Cry1Ac and Cry2Ab2 proteins in Bollgard-II hybrids at different stages of crop growth in different genotypes across cropping seasons and multiple geographies. Transgenic Res. 2018, 28, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-González, A.; Porteous-Álvarez, A.J.; Val, M.; Casquero, P.A.; Escriche, B. Toxicity of five Cry proteins against the insect pest Acanthoscelides obtectus (Coleoptera: Chrisomelidae: Bruchinae). J. Invertebr. Pathol. 2020, 169, 107295. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Wang, Q.; Nangong, Z.; Su, J.; Ge, D. Identification of Henosepilachna vigintioctomaculata (Coleoptera: Coccinellidae) midgut putative receptor for Bacillus thuringiensis insecticidal Cry7ab3 toxin. J. Invertebr. Pathol. 2012, 109, 318–322. [Google Scholar] [CrossRef]
- Rodríguez-González, A.; Porteous-Álvarez, A.J.; Guerra, M.; González-López, O.; Casquero, P.A.; Escriche, B. Effect of Cry toxins on Xylotrechus arvicola (Coleoptera: Cerambycidae) larvae. Insects 2021, 13, 27. [Google Scholar] [CrossRef]
- Guzov, V.M.; Malvar, T.M.; Roberts, J.K.; Sivasupramaniam, S. Insect inhibitory Bacillus thuringiensis proteins, fusions, and methods of use therefor. U.S. Patent 7,655,838, 2 February 2010. [Google Scholar]
- Contreras, E.; Rausell, C.; Real, M.D. Proteome response of Tribolium castaneum larvae to Bacillus thuringiensis toxin producing strains. PLoS ONE 2013, 8, e0055330. [Google Scholar] [CrossRef] [Green Version]
- Ekobu, M.; Solera, M.; Kyamanywa, S.; Mwanga, R.O.; Odongo, B.; Ghislain, M.; Moar, W.J. Toxicity of seven Bacillus thuringiensis Cry proteins against Cylas puncticollis and Cylas brunneus (Coleoptera: Brentidae) using a novel artificial diet. J. Econ. Entomol. 2010, 103, 1493–1502. [Google Scholar] [CrossRef]
- Hernández-Martínez, P.; Khorramnejad, A.; Prentice, K.; Andrés-Garrido, A.; Vera-Velasco, N.M.; Smagghe, G.; Escriche, B. The independent biological activity of Bacillus thuringiensis Cry23Aa protein against Cylas puncticollis. Front. Microbiol. 2020, 11, 1734. [Google Scholar] [CrossRef]
- Monnerat, R.G.; Dias, S.C.; Oliveira Neto, O.B.; Nobre, S.D.; Silva-Werneck, J.O.; Sá, M.F.G. Criação Massal do Bicudo do Algodoeiro Anthonomus grandis em Laboratório; Embrapa Recursos Genéticos e Biotecnologia: Brasília, Brazil, 2000; p. 4. [Google Scholar]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Bt Protein | n | Slope (±SE) | LC10 (FL95) | LC50 (FL95) | χ2 (d.f.) |
|---|---|---|---|---|---|
| Cry1Ac | 841 | 0.58 (0.16) | 2.01 (0.04–7.70) | 317 (188–3378) * | 2.08 (5) |
| Cry1Ba | 840 | 0.43 (0.11) | 0.33 (0.01–1.84) | 293 (92–4486) * | 2.05 (5) |
| Cry7Aa | 841 | 0.35 (0.12) | 0.38 (0.01–3.05) | 1598 (233–ND) * | 4.60 (5) |
| Mpp23Aa/Xpp37Aa | 735 | 3.80 (0.47) | 0.08 (0.04–0.12) * | 0.18 (0.13–0.22) | 4.43 (4) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, J.A.; Negri, B.F.; Hernández-Martínez, P.; Basso, M.F.; Escriche, B. Mpp23Aa/Xpp37Aa Insecticidal Proteins from Bacillus thuringiensis (Bacillales: Bacillaceae) Are Highly Toxic to Anthonomus grandis (Coleoptera: Curculionidae) Larvae. Toxins 2023, 15, 55. https://doi.org/10.3390/toxins15010055
de Oliveira JA, Negri BF, Hernández-Martínez P, Basso MF, Escriche B. Mpp23Aa/Xpp37Aa Insecticidal Proteins from Bacillus thuringiensis (Bacillales: Bacillaceae) Are Highly Toxic to Anthonomus grandis (Coleoptera: Curculionidae) Larvae. Toxins. 2023; 15(1):55. https://doi.org/10.3390/toxins15010055
Chicago/Turabian Stylede Oliveira, Jéssica A., Bárbara F. Negri, Patricia Hernández-Martínez, Marcos F. Basso, and Baltasar Escriche. 2023. "Mpp23Aa/Xpp37Aa Insecticidal Proteins from Bacillus thuringiensis (Bacillales: Bacillaceae) Are Highly Toxic to Anthonomus grandis (Coleoptera: Curculionidae) Larvae" Toxins 15, no. 1: 55. https://doi.org/10.3390/toxins15010055
APA Stylede Oliveira, J. A., Negri, B. F., Hernández-Martínez, P., Basso, M. F., & Escriche, B. (2023). Mpp23Aa/Xpp37Aa Insecticidal Proteins from Bacillus thuringiensis (Bacillales: Bacillaceae) Are Highly Toxic to Anthonomus grandis (Coleoptera: Curculionidae) Larvae. Toxins, 15(1), 55. https://doi.org/10.3390/toxins15010055