Discovery of an Insect Neuroactive Helix Ring Peptide from Ant Venom

 , , , , , , , , , , , ,
, , , , , , , , , , , ,  and
and
Abstract
:1. Introduction
2. Results
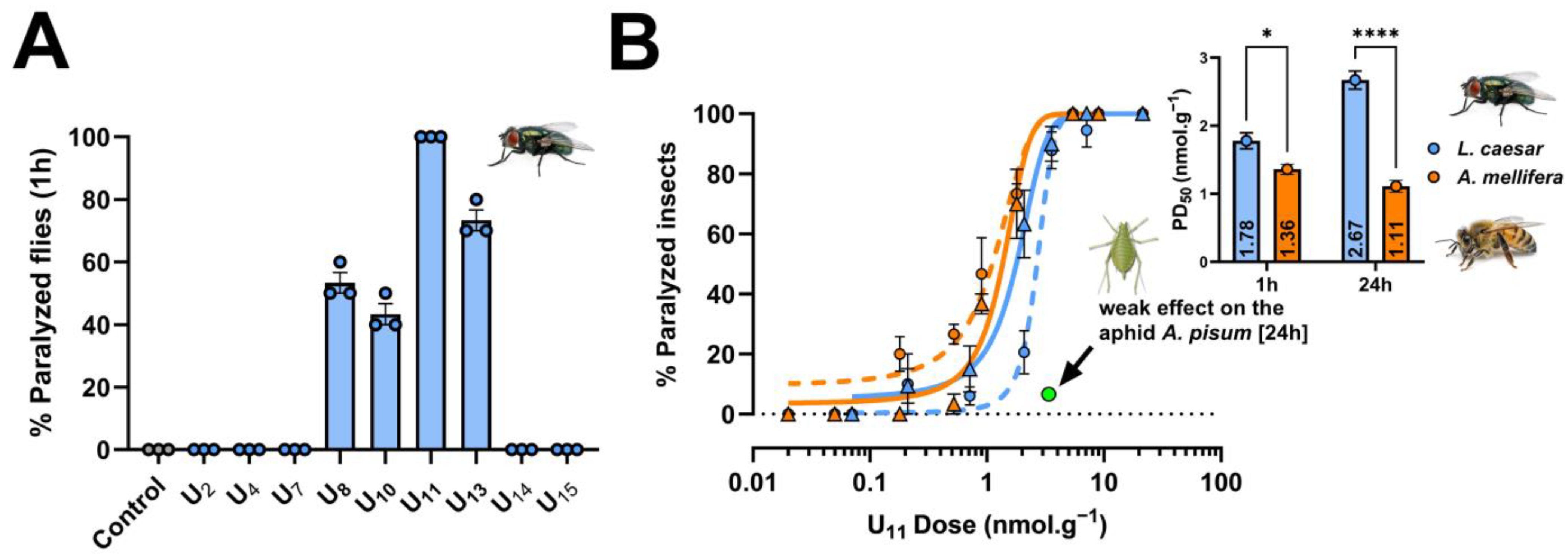
2.1. Neuroactivity Screen of Venom Peptides and Selection of U11
2.2. Mechanism of Action of U11
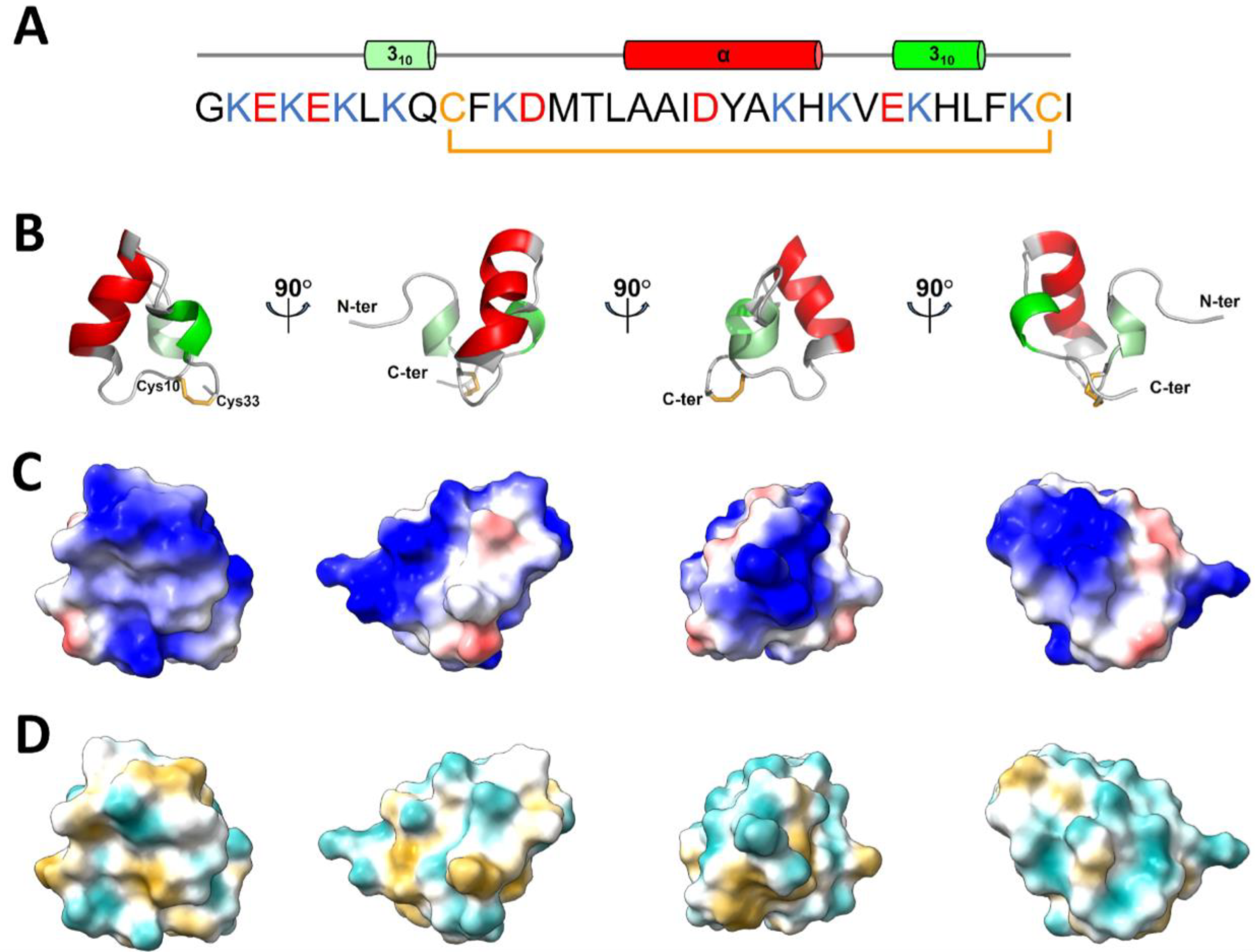
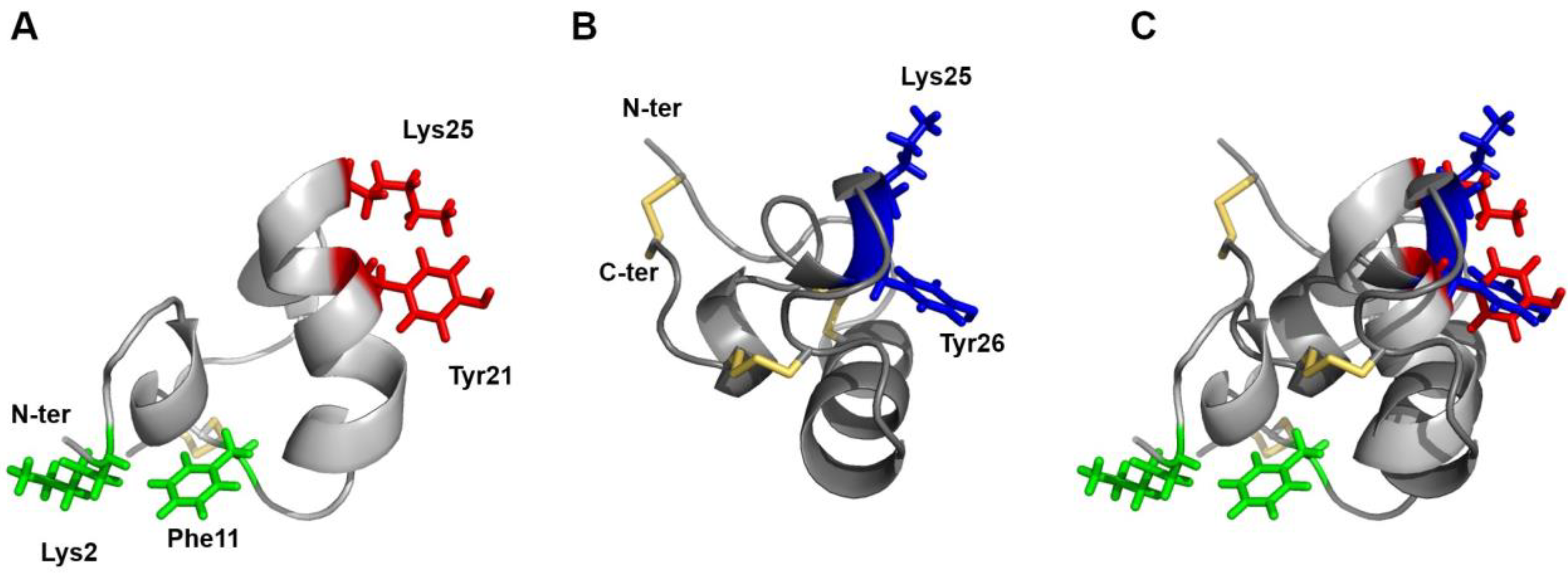
2.3. NMR Structure of U11
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Peptide Synthesis
5.2. Injection Assays
5.3. Ingestion Assays
5.4. S2 Cell Culture
5.5. Cytotoxicity Assays
5.6. Pharmacological Experiments
5.7. NMR Experiments
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef] [PubMed]
- Laxme, R.R.S.; Suranse, V.; Sunagar, K. Arthropod venoms: Biochemistry, ecology and evolution. Toxicon 2019, 158, 84–103. [Google Scholar] [CrossRef]
- Fanning, P.D.; VanWoerkom, A.; Wise, J.C.; Isaacs, R. Assessment of a commercial spider venom peptide against spotted-wing Drosophila and interaction with adjuvants. J. Pest Sci. 2018, 91, 1279–1290. [Google Scholar] [CrossRef]
- Nakasu, E.Y.T.; Williamson, S.M.; Edwards, M.G.; Fitches, E.C.; Gatehouse, J.A.; Wright, G.A.; Gatehouse, A.M.R. Novel biopesticide based on a spider venom peptide shows no adverse effects on honeybees. Proc. R. Soc. B Biol. Sci. 2014, 281. [Google Scholar] [CrossRef]
- Bolton, B. An Online Catalog of the Ants of the World. Available online: http://antcat.org (accessed on 8 September 2023).
- Wilson, E.O. Success and Dominance in Ecosystems: The Case of the Social Insects; Ecology Institute: Oldendorf/Luhe, Germany, 1990; Volume 2. [Google Scholar]
- Cerdá, X.; Dejean, A. Predation by ants on arthropods and other animals. In Predation in the Hymenoptera: An Evolutionary Perspective; Polidori, C., Ed.; TransWorld Research Network: India, Kerala, 2011; pp. 39–78. [Google Scholar]
- Maschwitz, U.; Hahn, M.; Schönegge, P. Paralysis of prey in ponerine ants. Naturwissenschaften 1979, 66, 213–214. [Google Scholar] [CrossRef]
- Aili, S.R.; Touchard, A.; Escoubas, P.; Padula, M.P.; Orivel, J.; Dejean, A.; Nicholson, G.M. Diversity of peptide toxins from stinging ant venoms. Toxicon 2014, 92, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Touchard, A.; Brust, A.; Cardoso, F.C.; Chin, Y.K.-Y.; Herzig, V.; Jin, A.-H.; Dejean, A.; Alewood, P.F.; King, G.F.; Orivel, J.; et al. Isolation and characterization of a structurally unique β-hairpin venom peptide from the predatory ant Anochetus emarginatus. Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 2553–2562. [Google Scholar] [CrossRef] [PubMed]
- Heep, J.; Klaus, A.; Kessel, T.; Seip, M.; Vilcinskas, A.; Skaljac, M. Proteomic analysis of the venom from the ruby ant Myrmica rubra and the isolation of a novel insecticidal decapeptide. Insects 2019, 10, 42. [Google Scholar] [CrossRef]
- Heep, J.; Skaljac, M.; Grotmann, J.; Kessel, T.; Seip, M.; Schmidtberg, H.; Vilcinskas, A. Identification and functional characterization of a novel insecticidal decapeptide from the myrmicine ant Manica rubida. Toxins 2019, 11, 562. [Google Scholar] [CrossRef]
- Hurka, S.; Lüddecke, T.; Paas, A.; Dersch, L.; Schulte, L.; Eichberg, J.; Hardes, K.; Brinkrolf, K.; Vilcinskas, A. Bioactivity profiling of in silico predicted linear toxins from the ants Myrmica rubra and Myrmica ruginodis. Toxins 2022, 14, 846. [Google Scholar] [CrossRef]
- Robinson, S.D.; Kambanis, L.; Clayton, D.; Hinneburg, H.; Corcilius, L.; Mueller, A.; Walker, A.A.; Keramidas, A.; Kulkarni, S.S.; Jones, A. A pain-causing and paralytic ant venom glycopeptide. iScience 2021, 24, 103175. [Google Scholar] [CrossRef] [PubMed]
- Touchard, A.; Aili, S.R.; Téné, N.; Barassé, V.; Klopp, C.; Dejean, A.; Kini, R.M.; Mrinalini; Coquet, L.; Jouenne, T.; et al. Venom peptide repertoire of the European myrmicine ant Manica rubida: Identification of insecticidal toxins. J. Proteome Res. 2020, 19, 1800–1811. [Google Scholar] [CrossRef] [PubMed]
- Von Reumont, B.M. Studying smaller and neglected organisms in modern evolutionary venomics implementing RNASeq (Transcriptomics)—A critical guide. Toxins 2018, 10, 292. [Google Scholar] [CrossRef]
- Robinson, S.D.; Mueller, A.; Clayton, D.; Starobova, H.; Hamilton, B.R.; Payne, R.J.; Vetter, I.; King, G.F.; Undheim, E.A.B. A Comprehensive portrait of the venom of the giant red bull ant, Myrmecia gulosa, reveals a hyperdiverse hymenopteran toxin gene family. Sci. Adv. 2018, 4, eaau4640. [Google Scholar] [CrossRef]
- Tani, N.; Kazuma, K.; Ohtsuka, Y.; Shigeri, Y.; Masuko, K.; Konno, K.; Inagaki, H. Mass spectrometry analysis and biological characterization of the predatory ant Odontomachus monticola venom and venom sac components. Toxins 2019, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Schendel, V.; Schroeder, C.I.; Moen, S.; Mueller, A.; Walker, A.A.; McKinnon, N.; Neely, G.G.; Vetter, I.; King, G.F.; et al. Intra-colony venom diversity contributes to maintaining eusociality in a cooperatively breeding ant. BMC Biol. 2023, 21, 5. [Google Scholar] [CrossRef] [PubMed]
- Touchard, A.; Téné, N.; Song, P.C.T.; Lefranc, B.; Leprince, J.; Treilhou, M.; Bonnafé, E. Deciphering the molecular diversity of an ant venom peptidome through a venomics approach. J. Proteome Res. 2018, 17, 3503–3516. [Google Scholar] [CrossRef]
- Ascoët, S.; Touchard, A.; Téné, N.; Lefranc, B.; Leprince, J.; Paquet, F.; Jouvensal, L.; Barassé, V.; Treilhou, M.; Billet, A.; et al. The mechanism underlying toxicity of a venom peptide against insects reveals how ants are master at disrupting membranes. iScience 2023, 26, 106157. [Google Scholar] [CrossRef]
- Duraisamy, K.; Singh, K.; Kumar, M.; Lefranc, B.; Bonnafé, E.; Treilhou, M.; Leprince, J.; Chow, B.K.C. P17 induces chemotaxis and differentiation of monocytes via MRGPRX2-mediated mast cell-line activation. J. Allergy Clin. Immunol. 2021, 149, 275–291. [Google Scholar] [CrossRef]
- Robinson, S.D.; Deuis, J.R.; Touchard, A.; Keramidas, A.; Mueller, A.; Schroeder, C.I.; Barassé, V.; Walker, A.A.; Brinkwirth, N.; Jami, S.; et al. Ant venoms contain vertebrate-selective pain-causing sodium channel toxins. Nat. Commun. 2023, 14, 2977. [Google Scholar] [CrossRef]
- Greer, J.A.; Moreau, C.S. Phylogenetic analysis and trait evolution of ant cocoons. Insect Syst. Evol. 2021, 53, 60–77. [Google Scholar] [CrossRef]
- Guo, S.; Herzig, V.; King, G.F. Dipteran toxicity assays for determining the oral insecticidal activity of venoms and toxins. Toxicon 2018, 150, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Dekan, Z.; Headey, S.J.; Scanlon, M.; Baldo, B.A.; Lee, T.; Aguilar, M.; Deuis, J.R.; Vetter, I.; Elliott, A.G.; Amado, M. Δ-Myrtoxin-Mp1a is a helical heterodimer from the venom of the jack jumper ant that has antimicrobial, membrane-disrupting, and nociceptive activities. Angew. Chem. 2017, 56, 8495–8499. [Google Scholar] [CrossRef]
- Nixon, S.A.; Robinson, S.D.; Agwa, A.J.; Walker, A.A.; Choudhary, S.; Touchard, A.; Undheim, E.A.B.; Robertson, A.; Vetter, I.; Schroeder, C.I.; et al. Multipurpose peptides: The venoms of Amazonian stinging ants contain anthelmintic ponericins with diverse predatory and defensive activities. Biochem. Pharmacol. 2021, 192, 114693. [Google Scholar] [CrossRef]
- Touchard, A.; Mendel, H.C.H.C.; Boulogne, I.; Herzig, V.; Braga Emidio, N.; King, G.F.G.F.; Triquigneaux, M.; Jaquillard, L.; Beroud, R.; De Waard, M.; et al. Heterodimeric insecticidal peptide provides new insights into the molecular and functional diversity of ant venoms. ACS Pharmacol. Transl. Sci. 2020, 3, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; King, M.A.; Donovan, G.R.; Alewood, D.; Alewood, P. Cytotoxicity of pilosulin 1, a peptide from the venom of the jumper ant Myrmecia pilosula. Biochim. Biophys. Acta Gen. Subj. 2006, 1425, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Barassé, V.; Téné, N.; Klopp, C.; Paquet, F.; Tysklind, N.; Troispoux, V.; Lalägue, H.; Orivel, J.; Lefranc, B.; Leprince, J.; et al. Venomics survey of six myrmicine ants provides insights into the molecular and structural diversity of their peptide toxins. Insect Biochem. Mol. Biol. 2022, 151, 103876. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Rullmann, J.A.C.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef]
- Guido-Patiño, J.C.; Plisson, F. Profiling hymenopteran venom toxins: Protein families, structural landscape, biological activities, and pharmacological benefits. Toxicon X 2022, 14, 100119. [Google Scholar] [CrossRef]
- Undheim, E.A.B.; Grimm, L.L.; Low, C.-F.; Morgenstern, D.; Herzig, V.; Zobel-Thropp, P.; Pineda, S.S.; Habib, R.; Dziemborowicz, S.; Fry, B.G.; et al. Weaponization of a hormone: Convergent recruitment of hyperglycemic hormone into the venom of arthropod predators. Structure 2015, 23, 1283–1292. [Google Scholar] [CrossRef]
- Holm, L. Benchmarking fold detection by DaliLite v. 5. Bioinformatics 2019, 35, 5326–5327. [Google Scholar] [CrossRef] [PubMed]
- Sakata, I.; Hayashi, M.; Nakamuta, K. Tetramorium Tsushimae Ants use methyl branched hydrocarbons of aphids for partner recognition. J. Chem. Ecol. 2017, 43, 966–970. [Google Scholar] [CrossRef] [PubMed]
- Bende, N.S.; Dziemborowicz, S.; Mobli, M.; Herzig, V.; Gilchrist, J.; Wagner, J.; Nicholson, G.M.; King, G.F.; Bosmans, F. A distinct sodium channel voltage-sensor locus determines insect selectivity of the spider toxin Dc1a. Nat. Commun. 2014, 5, 4350. [Google Scholar] [CrossRef] [PubMed]
- Tabakmakher, V.M.; Krylov, N.A.; Kuzmenkov, A.I.; Efremov, R.G.; Vassilevski, A.A. Kalium 2.0, a comprehensive database of polypeptide ligands of potassium channels. Sci. Data 2019, 6, 73. [Google Scholar] [CrossRef]
- Kuzmenkov, A.I.; Peigneur, S.; Nasburg, J.A.; Mineev, K.S.; Nikolaev, M.V.; Pinheiro-Junior, E.L.; Arseniev, A.S.; Wulff, H.; Tytgat, J.; Vassilevski, A.A. Apamin structure and pharmacology revisited. Front. Pharmacol. 2022, 13, 977440. [Google Scholar] [CrossRef]
- Jin, W.; Lu, Z. A novel high-affinity inhibitor for inward-rectifier K+ channels. Biochemistry 1998, 37, 13291–13299. [Google Scholar] [CrossRef]
- Mouhat, S.; Jouirou, B.; Mosbah, A.; De Waard, M.; Sabatier, J. Diversity of folds in animal toxins acting on ion channels. Biochem. J. 2004, 378, 717–726. [Google Scholar] [CrossRef]
- Chen, Z.-Y.; Zeng, D.-Y.; Hu, Y.-T.; He, Y.-W.; Pan, N.; Ding, J.-P.; Cao, Z.-J.; Liu, M.-L.; Li, W.-X.; Yi, H. Structural and functional diversity of acidic scorpion potassium channel toxins. PLoS ONE 2012, 7, e35154. [Google Scholar] [CrossRef]
- Zaharenko, A.J.; Ferreira, W.A., Jr.; Oliveira, J.S.; Richardson, M.; Pimenta, D.C.; Konno, K.; Portaro, F.C.V.; de Freitas, J.C. Proteomics of the neurotoxic fraction from the sea anemone Bunodosoma cangicum venom: Novel peptides belonging to new classes of toxins. Comp. Biochem. Physiol. Part D Genom. Proteom. 2008, 3, 219–225. [Google Scholar] [CrossRef]
- Massilia, G.R.; Schininà, M.E.; Ascenzi, P.; Polticelli, F. Contryphan-Vn: A novel peptide from the venom of the Mediterranean snail Conus ventricosus. Biochem. Biophys. Res. Commun. 2001, 288, 908–913. [Google Scholar] [CrossRef]
- Jiménez-Vargas, J.M.; Restano-Cassulini, R.; Possani, L.D. Toxin modulators and blockers of HERG K+ channels. Toxicon 2012, 60, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Norton, R.S.; Chandy, K.G. Venom-derived peptide inhibitors of voltage-gated potassium channels. Neuropharmacology 2017, 127, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Luo, Z.; Lei, S.; Li, S.; Li, X.; Yuan, C. Effects and mechanism of gating modifier spider toxins on the HERG channel. Toxicon 2021, 189, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Chung, S.-H. Computational studies of venom peptides targeting potassium channels. Toxins 2015, 7, 5194–5211. [Google Scholar] [CrossRef]
- Sanches, K.; Prypoten, V.; Chandy, K.G.; Chalmers, D.K.; Norton, R.S. Interaction of the inhibitory peptides ShK and HmK with the voltage-gated potassium channel KV1.3: Role of conformational dynamics. J. Chem. Inf. Model. 2023, 63, 3043–3053. [Google Scholar] [CrossRef]
- Dauplais, M.; Lecoq, A.; Song, J.; Cotton, J.; Jamin, N.; Gilquin, B.; Roumestand, C.; Vita, C.; de Medeiros, C.C.; Rowan, E.G. On the convergent evolution of animal toxins: Conservation of a diad of functional residues in potassium channel-blocking toxins with unrelated structures. J. Biol. Chem. 1997, 272, 4302–4309. [Google Scholar] [CrossRef]
- Mouhat, S.; De Waard, M.; Sabatier, J. Contribution of the functional dyad of animal toxins acting on voltage-gated KV1-type Channels. J. Pept. Sci. 2005, 11, 65–68. [Google Scholar] [CrossRef]
- Selvakumar, P.; Fernández-Mariño, A.I.; Khanra, N.; He, C.; Paquette, A.J.; Wang, B.; Huang, R.; Smider, V.V.; Rice, W.J.; Swartz, K.J. Structures of the T cell potassium channel KV1.3 with Immunoglobulin Modulators. Nat. Commun. 2022, 13, 3854. [Google Scholar] [CrossRef] [PubMed]
- Park, B.G.; Peigneur, S.; Esaki, N.; Yamaguchi, Y.; Ryu, J.H.; Tytgat, J.; Kim, J.I.L.; Sato, K. Solution structure and functional analysis of HelaTx1: The first toxin member of the κ-KTx5 subfamily. BMB Rep. 2020, 53, 260. [Google Scholar] [CrossRef]
- Batista, C.V.F.; Gómez-Lagunas, F.; de la Vega, R.C.R.; Hajdu, P.; Panyi, G.; Gáspár, R.; Possani, L.D. Two novel toxins from the Amazonian scorpion Tityus cambridgei that block KV1.3 and shaker BK+-channels with distinctly different affinities. Biochim. Biophys. Acta Proteins Proteom. 2002, 1601, 123–131. [Google Scholar] [CrossRef]
- Krishnarjuna, B.; Villegas-Moreno, J.; Mitchell, M.L.; Csoti, A.; Peigneur, S.; Amero, C.; Pennington, M.W.; Tytgat, J.; Panyi, G.; Norton, R.S. Synthesis, folding, structure and activity of a predicted peptide from the sea anemone Oulactis sp. with an ShKT fold. Toxicon 2018, 150, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Guschinskaya, N.; Ressnikoff, D.; Arafah, K.; Voisin, S.; Bulet, P.; Uzest, M.; Rahbé, Y. Insect mouthpart transcriptome unveils extension of cuticular protein repertoire and complex organization. iScience 2020, 23, 100828. [Google Scholar] [CrossRef]
- Sapountzis, P.; Duport, G.; Balmand, S.; Gaget, K.; Jaubert-Possamai, S.; Febvay, G.; Charles, H.; Rahbé, Y.; Colella, S.; Calevro, F. New insight into the RNA interference response against cathepsin-L gene in the pea aphid, Acyrthosiphon pisum: Molting or gut phenotypes specifically induced by injection or feeding treatments. Insect Biochem. Mol. Biol. 2014, 51, 20–32. [Google Scholar] [CrossRef]
- Ribeiro Lopes, M.; Gaget, K.; Renoz, F.; Duport, G.; Balmand, S.; Charles, H.; Callaerts, P.; Calevro, F. Bacteriocyte plasticity in pea aphids facing amino acid stress or starvation during development. Front. Physiol. 2022, 13, 982920. [Google Scholar] [CrossRef]
- Febvay, G.; Delobel, B.; Rahbé, Y. Influence of the amino acid balance on the improvement of an artificial diet for a biotype of Acyrthosiphon pisum (Homoptera: Aphididae). Can. J. Zool. 1988, 66, 2449–2453. [Google Scholar] [CrossRef]
- Radstake, F.D.W.; Raaijmakers, E.A.L.; Luttge, R.; Zinger, S.; Frimat, J.-P. CALIMA: The Semi-automated open-source calcium imaging analyzer. Comput. Methods Programs Biomed. 2019, 179, 104991. [Google Scholar] [CrossRef]
- Braunschweiler, L.; Ernst, R.R. Coherence transfer by isotropic mixing: Application to proton correlation spectroscopy. J. Magn. Reason. 1983, 53, 521–528. [Google Scholar] [CrossRef]
- Kumar, A. Two-dimensional nuclear overhauser effect in biomolecules. Proc. Indian Acad. Sci. Chem. Sci. 1985, 95, 1–8. [Google Scholar] [CrossRef]
- Schanda, P.; Kupče, Ē.; Brutscher, B. SOFAST-HMQC Experiments for recording two-dimensional deteronuclear correlation spectra of proteins within a few seconds. J. Biomol. NMR 2005, 33, 199–211. [Google Scholar] [CrossRef]
- Bodenhausen, G.; Ruben, D.J. Natural abundance nitrogen-15 NMR by enhanced heteronuclear spectroscopy. Chem. Phys. Lett. 1980, 69, 185–189. [Google Scholar] [CrossRef]
- Schleucher, J.; Schwendinger, M.; Sattler, M.; Schmidt, P.; Schedletzky, O.; Glaser, S.J.; Sørensen, O.W.; Griesinger, C.A. General enhancement scheme in heteronuclear multidimensional NMR employing pulsed field gradients. J. Biomol. NMR 1994, 4, 301–306. [Google Scholar] [CrossRef]
- Vranken, W.F.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins Struct. Funct. Bioinform. 2005, 59, 687–696. [Google Scholar] [CrossRef]
- Brunger, A.T. Version 1.2 of the crystallography and NMR system. Nat. Protoc. 2007, 2, 2728–2733. [Google Scholar] [CrossRef] [PubMed]
- Brünger, A.T.; Adams, P.D.; Clore, G.M.; Delano, W.L.; Gros, P.; Grosse-Kunstleve, R.W.; Jiang, J.S.; Kuszewsk, J.I.; Nilges, M.; Pannu, N.S. Crystallography & NMR System (CNS), Version 0.5; Yale University: New Haven, CT, USA, 1998. [Google Scholar]
- Rieping, W.; Habeck, M.; Bardiaux, B.; Bernard, A.; Malliavin, T.E.; Nilges, M. ARIA2: Automated NOE assignment and data integration in NMR structure calculation. Bioinformatics 2007, 23, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.-S.; Maguire, M.L.; Stevens, T.J.; Broadhurst, R.W. DANGLE: A bayesian inferential method for predicting protein backbone dihedral angles and secondary structure. J. Magn. Reson. 2010, 202, 223–233. [Google Scholar] [CrossRef]
- Hutchinson, E.G.; Thornton, J.M. PROMOTIF—A program to identify and analyze structural motifs in proteins. Protein Sci. 1996, 5, 212–220. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | MW # | Injected Dose ¥ |
|---|---|---|---|
| U2 | DPPPGFIGVR * | 1052.6 | 103 nmol. g−1 |
| U4 | GCSQFRRMRNLCG * | 1523.7 | 68 nmol. g−1 |
| U7 | AINCRRYPRHPKCRGVSA | 2081.1 | 52 nmol. g−1 |
| U8 | GMLDRILGAVKGFMGS | 1650.9 | 54 nmol. g−1 |
| U10 | GLGFLAKIMGKVGMRMIKKLVPEAAKVAVDQLSQQQ | 3882.2 | 28 nmol. g−1 |
| U11 | GKEKEKLKQCFKDMTLAAIDYAKHKVEKHLFKCI | 4018.1 | 27 nmol. g−1 |
| U13 | RPPQIGIFDQIDKGMAAFMDLFK * | 2636.4 | 45 nmol. g−1 |
| U14 | IPPNAVKSLQ * | 1064.6 | 95 nmol. g−1 |
| U15 | VFLTPDQIKAMIKRH * | 1795.0 | 48 nmol. g−1 |
| Peptide | Venom | Blowfly Species | PD50 [1 h] * | Cytotoxicity | Reference |
|---|---|---|---|---|---|
| Rm1a | Rhytidoponera metallica | L. caesar | 0.3 ± 0.0 | N.D. | [23] |
| Nc3a | Neoponera commutata | L. cuprina | 0.5 ± 0.0 | Yes | [27] |
| U11-Tb1a | Tetramorium bicarinatum | L. caesar | 1.8 ± 0.1 | No | This study |
| Pp1a | Pseudomyrmex penetrator | L. cuprina | 2.4 ± 0.6 | Yes | [28] |
| U20-Mri1a | Manica rubida | L. caesar | 2.9 ± 0.9 | N.D. | [15] |
| Ta2a | Tetramorium africanum | L. caesar | 7.1 ± 0.9 | N.D. | [23] |
| Ae1a | Anochetus emarginatus | L. cuprina | 8.9 ± 3.1 | N.D. | [10] |
| U10-Mri1c | Manica rubida | L. caesar | 10.5 ± 1.7 | N.D. | [15] |
| M-Tb1a | Tetramorium bicarinatum | L. caesar | 12.0 ± 1.0 | Yes | [21] |
| U10-Mri1a | Manica rubida | L. caesar | 12.1 ± 1.5 | N.D. | [15] |
| U9-Tb1a | Tetramorium bicarinatum | L. caesar | 13.1 ± 3.0 | Yes | [21] |
| Na1b | Neoponera apicalis | L. cuprina | 25.8 ± 13.9 | Yes | [27] |
| Nc3b | Neoponera commutata | L. cuprina | 26.4 ± 5.9 | Yes | [27] |
| Nc1a | Neoponera commutata | L. cuprina | 31.4 ± 6.3 | Yes | [27] |
| Nc2a | Neoponera commutata | L. cuprina | 38.1 ± 18.5 | Yes | [27] |
| U10-Mri1b | Manica rubida | L. caesar | 58.4 ± 7.9 | N.D. | [15] |
| U13-Mri1a | Manica rubida | L. caesar | 69.2 ± 8.6 | N.D. | [15] |
| Pc1a | Paraponera clavata | L. caesar | 74.3 ± 2.1 | N.D. | [23] |
| Ta3a | Tetramorium africanum | L. caesar | 77.8 ± 1.4 | N.D. | [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barassé, V.; Jouvensal, L.; Boy, G.; Billet, A.; Ascoët, S.; Lefranc, B.; Leprince, J.; Dejean, A.; Lacotte, V.; Rahioui, I.; et al. Discovery of an Insect Neuroactive Helix Ring Peptide from Ant Venom. Toxins 2023, 15, 600. https://doi.org/10.3390/toxins15100600
Barassé V, Jouvensal L, Boy G, Billet A, Ascoët S, Lefranc B, Leprince J, Dejean A, Lacotte V, Rahioui I, et al. Discovery of an Insect Neuroactive Helix Ring Peptide from Ant Venom. Toxins. 2023; 15(10):600. https://doi.org/10.3390/toxins15100600
Chicago/Turabian StyleBarassé, Valentine, Laurence Jouvensal, Guillaume Boy, Arnaud Billet, Steven Ascoët, Benjamin Lefranc, Jérôme Leprince, Alain Dejean, Virginie Lacotte, Isabelle Rahioui, and et al. 2023. "Discovery of an Insect Neuroactive Helix Ring Peptide from Ant Venom" Toxins 15, no. 10: 600. https://doi.org/10.3390/toxins15100600
APA StyleBarassé, V., Jouvensal, L., Boy, G., Billet, A., Ascoët, S., Lefranc, B., Leprince, J., Dejean, A., Lacotte, V., Rahioui, I., Sivignon, C., Gaget, K., Ribeiro Lopes, M., Calevro, F., Da Silva, P., Loth, K., Paquet, F., Treilhou, M., Bonnafé, E., & Touchard, A. (2023). Discovery of an Insect Neuroactive Helix Ring Peptide from Ant Venom. Toxins, 15(10), 600. https://doi.org/10.3390/toxins15100600