Exosome Liberation by Human Neutrophils under L-Amino Acid Oxidase of Calloselasma rhodostoma Venom Action

 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
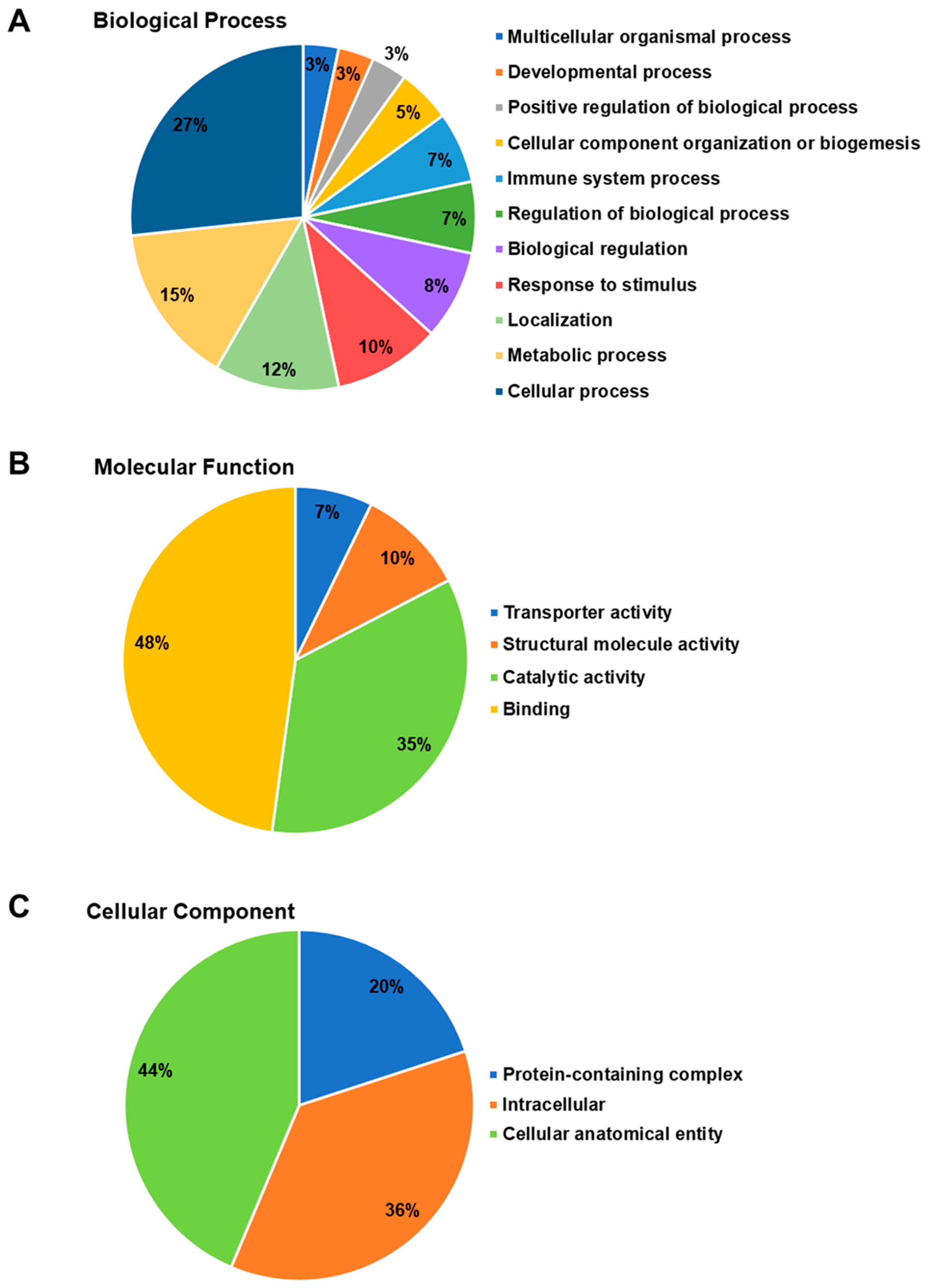
2. Results
3. Discussion
- Desmin, encoded by the DES gene, represents a muscle-specific ɪɪɪ-type intermediate filament crucial for proper muscle structure and function [68];
- Antithrombin ɪɪɪ, encoded by the SERPINC1 gene, is a major serine protease inhibitor in plasma that regulates the blood clotting cascade by neutralizing thrombin, thereby inhibiting thrombosis [71];
- Kininogen-1, encoded by the KNG1 gene, is involved in blood clotting, inhibits thrombocyte aggregation, and has pro-inflammatory and pro-oxidant properties [83];
- Peptidase D, encoded by the PEPD gene, plays a role in proline recycling and may be a limiting factor for collagen production [88];
4. Material and Methods
4.1. Chemicals and Reagents
4.2. Cr-LAAO Isolation
4.3. Isolation and Activation of Neutrophils
4.4. Exosomes Isolation
4.5. Protein Profile of Neutrophil Supernatants
4.6. Western Blot Protein Expression
4.7. Proteomic Analysis
4.8. Mass Spectrometry Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Wang, L.; Sun, X.; Li, F. SERPINB4 Promotes Keratinocyte Inflammation via P38MAPK Signaling Pathway. J. Immunol. Res. 2023, 2023, 3397940. [Google Scholar] [CrossRef] [PubMed]
- Du, X.-Y.; Clemetson, K.J. Snake Venom L-Amino Acid Oxidases. Toxicon 2002, 40, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Zuliani, J.; Kayano, A.; Zaqueo, K.; Neto, A.; Sampaio, S.; Soares, A.; Stabeli, R. Snake Venom L-Amino Acid Oxidases: Some Consideration About Their Functional Characterization. Protein Pept. Lett. 2009, 16, 908–912. [Google Scholar] [CrossRef] [PubMed]
- da Silva, S.L.; Calderon, L.A.; Zanchi, F.B.; Mendes, M.M.; Grabner, A.N.; Zuliani, J.P.; Fernandes, C.F.C.; Izidoro, L.F.M.; Costa, T.R.; Stábeli, R.G.; et al. Snake Venom L-Amino Acid Oxidases: Trends in Pharmacology and Biochemistry. Biomed. Res. Int. 2014, 2014, 196754. [Google Scholar] [CrossRef]
- Paloschi, M.V.; Pontes, A.S.; Soares, A.M.; Zuliani, J.P. An Update on Potential Molecular Mechanisms Underlying the Actions of Snake Venom L-Amino Acid Oxidases (LAAOs). Curr. Med. Chem. 2018, 25, 2520–2530. [Google Scholar] [CrossRef]
- Guo, C.; Liu, S.; Yao, Y.; Zhang, Q.; Sun, M.-Z. Past Decade Study of Snake Venom L-Amino Acid Oxidase. Toxicon 2012, 60, 302–311. [Google Scholar] [CrossRef]
- Zuliani, J.P.; Paloschi, M.V.; Pontes, A.S.; Boeno, C.N.; Lopes, J.A.; Setubal, S.S.; Zanchi, F.B.; Soares, A.M. Reptile Venom L-Amino Acid Oxidases—Structure and Function. In Handbook of Venoms and Toxins of Reptiles; CRC Press: Boca Raton, FL, USA, 2021; Volume 2, pp. 413–430. [Google Scholar] [CrossRef]
- Suhr, S.M.; Kim, D.S. Identification of the Snake Venom Substance That Induces Apoptosis. Biochem. Biophys. Res. Commun. 1996, 224, 134–139. [Google Scholar] [CrossRef]
- Ahn, M.Y.; Lee, B.M.; Kim, Y.S. Characterization and Cytotoxicity of L-Amino Acid Oxidase from the Venom of King Cobra (Ophiophagus hannah). Int. J. Biochem. Cell Biol. 1997, 29, 911–919. [Google Scholar] [CrossRef]
- Li, Z.-Y.; Yu, T.-F.; Lian, E.C.-Y. Purification and Characterization of L-Amino Acid Oxidase from King Cobra (Ophiophagus hannah) Venom and Its Effects on Human Platelet Aggregation. Toxicon 1994, 32, 1349–1358. [Google Scholar] [CrossRef]
- Sakurai, Y.; Shima, M.; Matsumoto, T.; Takatsuka, H.; Nishiya, K.; Kasuda, S.; Fujimura, Y.; Yoshioka, A. Anticoagulant Activity of M-LAO, l-Amino Acid Oxidase Purified from Agkistrodon Halys Blomhoffii, through Selective Inhibition of Factor IX. Biochim. Biophys. Acta-Proteins Proteom. 2003, 1649, 51–57. [Google Scholar] [CrossRef]
- Abe, Y.; Shimoyama, Y.; Munakata, H.; Ito, J.; Nagata, N.; Ohtsuki, K. Characterization of an Apoptosis-Inducing Factor in Habu Snake Venom as a Glycyrrhizin (GL)-Binding Protein Potently Inhibited by GL in Vitro. Biol. Pharm. Bull. 1998, 21, 924–927. [Google Scholar] [CrossRef] [PubMed]
- Stábeli, R.G.; Marcussi, S.; Carlos, G.B.; Pietro, R.C.L.R.; Selistre-de-Araújo, H.S.; Giglio, J.R.; Oliveira, E.B.; Soares, A.M. Platelet Aggregation and Antibacterial Effects of an L-Amino Acid Oxidase Purified from Bothrops Alternatus Snake Venom. Bioorg. Med. Chem. 2004, 12, 2881–2886. [Google Scholar] [CrossRef] [PubMed]
- França, S.C.; Kashima, S.; Roberto, P.G.; Marins, M.; Ticli, F.K.; Pereira, J.O.; Astolfi-Filho, S.; Stábeli, R.G.; Magro, A.J.; Fontes, M.R.M.; et al. Molecular Approaches for Structural Characterization of Bothrops L-Amino Acid Oxidases with Antiprotozoal Activity: CDNA Cloning, Comparative Sequence Analysis, and Molecular Modeling. Biochem. Biophys. Res. Commun. 2007, 355, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Tempone, A.G.; Andrade, H.F.; Spencer, P.J.; Lourenço, C.O.; Rogero, J.R.; Nascimento, N. Bothrops Moojeni Venom Kills Leishmania Spp. with Hydrogen Peroxide Generated by Its -Amino Acid Oxidase. Biochem. Biophys. Res. Commun. 2001, 280, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Fung, S.Y.; Lee, M.L.; Tan, N.H. Molecular Mechanism of Cell Death Induced by King Cobra (Ophiophagus hannah) Venom l-Amino Acid Oxidase. Toxicon 2015, 96, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Pontes, A.S.; da S. Setúbal, S.; Xavier, C.V.; Lacouth-Silva, F.; Kayano, A.M.; Pires, W.L.; Nery, N.M.; Boeri de Castro, O.; da Silva, S.D.; Calderon, L.A.; et al. Effect of L-Amino Acid Oxidase from Calloselasma Rhodosthoma Snake Venom on Human Neutrophils. Toxicon 2014, 80, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Pontes, A.S.; Setúbal, S.D.S.; Nery, N.M.; Da Silva, F.S.; Da Silva, S.D.; Fernandes, C.F.C.; Stábeli, R.G.; Soares, A.M.; Zuliani, J.P. P38 MAPK Is Involved in Human Neutrophil Chemotaxis Induced by L-Amino Acid Oxidase from Calloselasma Rhodosthoma. Toxicon 2016, 119, 106–116. [Google Scholar] [CrossRef]
- Paloschi, M.V.; Boeno, C.N.; Lopes, J.A.; Eduardo dos Santos da Rosa, A.; Pires, W.L.; Pontes, A.S.; da Silva Setúbal, S.; Soares, A.M.; Zuliani, J.P. Role of L-Amino Acid Oxidase Isolated from Calloselasma Rhodostoma Venom on Neutrophil NADPH Oxidase Complex Activation. Toxicon 2018, 145, 48–55. [Google Scholar] [CrossRef]
- Paloschi, M.V.; Lopes, J.A.; Boeno, C.N.; Silva, M.D.S.; Evangelista, J.R.; Pontes, A.S.; da Silva Setúbal, S.; Rego, C.M.A.; Néry, N.M.; Ferreira e Ferreira, A.A.; et al. Cytosolic Phospholipase A2-α Participates in Lipid Body Formation and PGE2 Release in Human Neutrophils Stimulated with an l-Amino Acid Oxidase from Calloselasma Rhodostoma Venom. Sci. Rep. 2020, 10, 10976. [Google Scholar] [CrossRef]
- Paloschi, M.V.; Boeno, C.N.; Lopes, J.A.; Rego, C.M.A.; Silva, M.D.S.; Santana, H.M.; Serrath, S.N.; Ikenohuchi, Y.J.; Farias, B.J.C.; Felipin, K.P.; et al. Reactive Oxygen Species-Dependent-NLRP3 Inflammasome Activation in Human Neutrophils Induced by l-Amino Acid Oxidase Derived from Calloselasma Rhodostoma Venom. Life Sci. 2022, 308, 120962. [Google Scholar] [CrossRef]
- Torii, S.; Naito, M.; Tsuruo, T. Apoxin I, a Novel Apoptosis-Inducing Factor with L-Amino Acid Oxidase Activity Purified from Western Diamondback Rattlesnake Venom. J. Biol. Chem. 1997, 272, 9539–9542. [Google Scholar] [CrossRef] [PubMed]
- Suhr, S.-M.; Kim, D.-S. Comparison of the Apoptotic Pathways Induced by L-Amino Acid Oxidase and Hydrogen Peroxide. J. Biochem. 1999, 125, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.R.; Menaldo, D.L.; Zoccal, K.F.; Burin, S.M.; Aissa, A.F.; de Castro, F.A.; Faccioli, L.H.; Greggi Antunes, L.M.; Sampaio, S.V. CR-LAAO, an L-Amino Acid Oxidase from Calloselasma Rhodostoma Venom, as a Potential Tool for Developing Novel Immunotherapeutic Strategies against Cancer. Sci. Rep. 2017, 7, 42673. [Google Scholar] [CrossRef] [PubMed]
- van der Pol, E.; Böing, A.N.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, Functions, and Clinical Relevance of Extracellular Vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [CrossRef]
- Majumdar, R.; Tavakoli Tameh, A.; Arya, S.B.; Parent, C.A. Exosomes Mediate LTB4 Release during Neutrophil Chemotaxis. PLoS Biol. 2021, 19, e3001271. [Google Scholar] [CrossRef]
- Higginbotham, J.N.; Demory Beckler, M.; Gephart, J.D.; Franklin, J.L.; Bogatcheva, G.; Kremers, G.-J.; Piston, D.W.; Ayers, G.D.; McConnell, R.E.; Tyska, M.J.; et al. Amphiregulin Exosomes Increase Cancer Cell Invasion. Curr. Biol. 2011, 21, 779–786. [Google Scholar] [CrossRef]
- Atayde, V.D.; Hassani, K.; da Silva Lira Filho, A.; Borges, A.R.; Adhikari, A.; Martel, C.; Olivier, M. Leishmania Exosomes and Other Virulence Factors: Impact on Innate Immune Response and Macrophage Functions. Cell. Immunol. 2016, 309, 7–18. [Google Scholar] [CrossRef]
- Karasu, E.; Eisenhardt, S.U.; Harant, J.; Huber-Lang, M. Extracellular Vesicles: Packages Sent with Complement. Front. Immunol. 2018, 9, 721. [Google Scholar] [CrossRef]
- Han, Q.-F.; Li, W.-J.; Hu, K.-S.; Gao, J.; Zhai, W.-L.; Yang, J.-H.; Zhang, S.-J. Exosome Biogenesis: Machinery, Regulation, and Therapeutic Implications in Cancer. Mol. Cancer 2022, 21, 207. [Google Scholar] [CrossRef]
- Ludwig, A.-K.; Giebel, B. Exosomes: Small Vesicles Participating in Intercellular Communication. Int. J. Biochem. Cell Biol. 2012, 44, 11–15. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, W.; Zhang, H.; Zhang, F.; Chen, L.; Ma, L.; Larcher, L.M.; Chen, S.; Liu, N.; Zhao, Q.; et al. Progress, Opportunity, and Perspective on Exosome Isolation—Efforts for Efficient Exosome-Based Theranostics. Theranostics 2020, 10, 3684–3707. [Google Scholar] [CrossRef] [PubMed]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of Extracellular Vesicles (EV): Exosomes, Microvesicles, Retrovirus-like Vesicles, and Apoptotic Bodies. J. Neurooncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The Biology, Function, and Biomedical Applications of Exosomes. Science (80-) 2020, 367. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, N.A.; Sampey, G.C.; Lepene, B.; Akpamagbo, Y.; Barclay, R.A.; Iordanskiy, S.; Hakami, R.M.; Kashanchi, F. Presence of Viral RNA and Proteins in Exosomes from Cellular Clones Resistant to Rift Valley Fever Virus Infection. Front. Microbiol. 2016, 7, 139. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, Y.; Qian, X. Target-Specific Exosome Isolation through Aptamer-Based Microfluidics. Biosensors 2022, 12, 257. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, J.W. 3D Spheroid Cultures of Stem Cells and Exosome Applications for Cartilage Repair. Life 2022, 12, 939. [Google Scholar] [CrossRef]
- Ye, Q.; Li, Z.; Li, Y.; Li, Y.; Zhang, Y.; Gui, R.; Cui, Y.; Zhang, Q.; Qian, L.; Xiong, Y.; et al. Exosome-Derived MicroRNA: Implications in Melanoma Progression, Diagnosis and Treatment. Cancers 2022, 15, 80. [Google Scholar] [CrossRef]
- Aswad, H.; Forterre, A.; Wiklander, O.P.B.; Vial, G.; Danty-Berger, E.; Jalabert, A.; Lamazière, A.; Meugnier, E.; Pesenti, S.; Ott, C.; et al. Exosomes Participate in the Alteration of Muscle Homeostasis during Lipid-Induced Insulin Resistance in Mice. Diabetologia 2014, 57, 2155–2164. [Google Scholar] [CrossRef]
- Ratajczak, J.; Miekus, K.; Kucia, M.; Zhang, J.; Reca, R.; Dvorak, P.; Ratajczak, M.Z. Embryonic Stem Cell-Derived Microvesicles Reprogram Hematopoietic Progenitors: Evidence for Horizontal Transfer of MRNA and Protein Delivery. Leukemia 2006, 20, 847–856. [Google Scholar] [CrossRef]
- Distler, J.H.W.; Huber, L.C.; Gay, S.; Distler, O.; Pisetsky, D.S. Microparticles as Mediators of Cellular Cross-Talk in Inflammatory Disease. Autoimmunity 2006, 39, 683–690. [Google Scholar] [CrossRef]
- Boulanger, C.M.; Amabile, N.; Tedgui, A. Circulating Microparticles. Hypertension 2006, 48, 180–186. [Google Scholar] [CrossRef]
- Bobrie, A.; Colombo, M.; Raposo, G.; Théry, C. Exosome Secretion: Molecular Mechanisms and Roles in Immune Responses. Traffic 2011, 12, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, S.; Wu, S.; Chen, L. Exosomes Modulate the Viral Replication and Host Immune Responses in HBV Infection. Biomed. Res. Int. 2019, 2019, 2103943. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Ratajczak, J. Innate Immunity Communicates Using the Language of Extracellular Microvesicles. Stem Cell Rev. Rep. 2021, 17, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, L.; Han, L.; Li, Y.; Hu, L.; Wang, H.; Zou, F. TRX2/Rab35 Interaction Impairs Exosome Secretion by Inducing Rab35 Degradation. Int. J. Mol. Sci. 2022, 23, 6557. [Google Scholar] [CrossRef]
- Heo, J.; Kang, H. Exosome-Based Treatment for Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 1002. [Google Scholar] [CrossRef]
- Lässer, C.; Eldh, M.; Lötvall, J. Isolation and Characterization of RNA-Containing Exosomes. J. Vis. Exp. 2012. [Google Scholar] [CrossRef]
- Khashayar, P.; Amoabediny, G.; Larijani, B.; Hosseini, M.; Vanfleteren, J. Fabrication and Verification of Conjugated AuNP-Antibody Nanoprobe for Sensitivity Improvement in Electrochemical Biosensors. Sci. Rep. 2017, 7, 16070. [Google Scholar] [CrossRef]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-Gel Digestion for Mass Spectrometric Characterization of Proteins and Proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef]
- Forrest, D.M.; Batista, M.; Marchini, F.K.; Tempone, A.J.; Traub-Csekö, Y.M. Proteomic Analysis of Exosomes Derived from Procyclic and Metacyclic-like Cultured Leishmania Infantum Chagasi. J. Proteom. 2020, 227, 103902. [Google Scholar] [CrossRef]
- Carvalho, P.C.; Lima, D.B.; Leprevost, F.V.; Santos, M.D.M.; Fischer, J.S.G.; Aquino, P.F.; Moresco, J.J.; Yates, J.R.; Barbosa, V.C. Integrated Analysis of Shotgun Proteomic Data with PatternLab for Proteomics 4.0. Nat. Protoc. 2016, 11, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.J.H.; Yang, X.; Don, C.W.; Minami, E.; Liu, Y.-W.; Weyers, J.J.; Mahoney, W.M.; Van Biber, B.; Cook, S.M.; Palpant, N.J.; et al. Human Embryonic-Stem-Cell-Derived Cardiomyocytes Regenerate Non-Human Primate Hearts. Nature 2014, 510, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Eitan, E.; Suire, C.; Zhang, S.; Mattson, M.P. Impact of Lysosome Status on Extracellular Vesicle Content and Release. Ageing Res. Rev. 2016, 32, 65–74. [Google Scholar] [CrossRef]
- Yamashita, T.; Takahashi, Y.; Nishikawa, M.; Takakura, Y. Effect of Exosome Isolation Methods on Physicochemical Properties of Exosomes and Clearance of Exosomes from the Blood Circulation. Eur. J. Pharm. Biopharm. 2016, 98, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pols, M.S.; Klumperman, J. Trafficking and Function of the Tetraspanin CD63. Exp. Cell Res. 2009, 315, 1584–1592. [Google Scholar] [CrossRef]
- Kowal, E.J.K.; Ter-Ovanesyan, D.; Regev, A.; Church, G.M. Extracellular Vesicle Isolation and Analysis by Western Blotting. In Extracellular Vesicles. Methods in Molecular Biology; Humana Press: New York, NY, USA, 2017; pp. 143–152. [Google Scholar]
- Multia, E.; Tear, C.J.Y.; Palviainen, M.; Siljander, P.; Riekkola, M.-L. Fast Isolation of Highly Specific Population of Platelet-Derived Extracellular Vesicles from Blood Plasma by Affinity Monolithic Column, Immobilized with Anti-Human CD61 Antibody. Anal. Chim. Acta 2019, 1091, 160–168. [Google Scholar] [CrossRef]
- Palviainen, M.; Saraswat, M.; Varga, Z.; Kitka, D.; Neuvonen, M.; Puhka, M.; Joenväärä, S.; Renkonen, R.; Nieuwland, R.; Takatalo, M.; et al. Extracellular Vesicles from Human Plasma and Serum Are Carriers of Extravesicular Cargo—Implications for Biomarker Discovery. PLoS ONE 2020, 15, e0236439. [Google Scholar] [CrossRef]
- Goel, G.; Makkar, H.P.S.; Francis, G.; Becker, K. Phorbol Esters: Structure, Biological Activity, and Toxicity in Animals. Int. J. Toxicol. 2007, 26, 279–288. [Google Scholar] [CrossRef]
- Antebi, Y.E.; Nandagopal, N.; Elowitz, M.B. An Operational View of Intercellular Signaling Pathways. Curr. Opin. Syst. Biol. 2017, 1, 16–24. [Google Scholar] [CrossRef]
- Carmona, B.; Marinho, H.S.; Matos, C.L.; Nolasco, S.; Soares, H. Tubulin Post-Translational Modifications: The Elusive Roles of Acetylation. Biology 2023, 12, 561. [Google Scholar] [CrossRef]
- Boggs, A.E.; Vitolo, M.I.; Whipple, R.A.; Charpentier, M.S.; Goloubeva, O.G.; Ioffe, O.B.; Tuttle, K.C.; Slovic, J.; Lu, Y.; Mills, G.B.; et al. α-Tubulin Acetylation Elevated in Metastatic and Basal-like Breast Cancer Cells Promotes Microtentacle Formation, Adhesion, and Invasive Migration. Cancer Res. 2015, 75, 203–215. [Google Scholar] [CrossRef]
- Seeley, E.S.; Carrière, C.; Goetze, T.; Longnecker, D.S.; Korc, M. Pancreatic Cancer and Precursor Pancreatic Intraepithelial Neoplasia Lesions Are Devoid of Primary Cilia. Cancer Res. 2009, 69, 422–430. [Google Scholar] [CrossRef]
- Segelmark, M.; Persson, B.; Hellmark, T.; Wieslander, J. Binding and Inhibition of Myeloperoxidase (MPO): A Major Function of Ceruloplasmin? Clin. Exp. Immunol. 2003, 108, 167–174. [Google Scholar] [CrossRef]
- Hilhorst, M.; van Paassen, P.; van Rie, H.; Bijnens, N.; Heerings-Rewinkel, P.; van Breda Vriesman, P.; Cohen Tervaert, J.W. Complement in ANCA-Associated Glomerulonephritis. Nephrol. Dial. Transplant. 2017, 32, 1302–1313. [Google Scholar] [CrossRef]
- Hakroush, S.; Tampe, D.; Korsten, P.; Ströbel, P.; Tampe, B. Complement Components C3 and C4 Indicate Vasculitis Manifestations to Distinct Renal Compartments in ANCA-Associated Glomerulonephritis. Int. J. Mol. Sci. 2021, 22, 6588. [Google Scholar] [CrossRef]
- Baier, E.; Tampe, D.; Kluge, I.A.; Hakroush, S.; Tampe, B. Implication of Platelets and Complement C3 as Link between Innate Immunity and Tubulointerstitial Injury in Renal Vasculitis with MPO-ANCA Seropositivity. Front. Immunol. 2022, 13, 1054457. [Google Scholar] [CrossRef]
- Siegert, R.; Perrot, A.; Keller, S.; Behlke, J.; Michalewska-Włudarczyk, A.; Wycisk, A.; Tendera, M.; Morano, I.; Özcelik, C. A Myomesin Mutation Associated with Hypertrophic Cardiomyopathy Deteriorates Dimerisation Properties. Biochem. Biophys. Res. Commun. 2011, 405, 473–479. [Google Scholar] [CrossRef]
- Prill, K.; Carlisle, C.; Stannard, M.; Windsor Reid, P.J.; Pilgrim, D.B. Myomesin Is Part of an Integrity Pathway That Responds to Sarcomere Damage and Disease. PLoS ONE 2019, 14, e0224206. [Google Scholar] [CrossRef]
- Lamber, E.P.; Guicheney, P.; Pinotsis, N. The Role of the M-Band Myomesin Proteins in Muscle Integrity and Cardiac Disease. J. Biomed. Sci. 2022, 29, 18. [Google Scholar] [CrossRef]
- Hnia, K.; Ramspacher, C.; Vermot, J.; Laporte, J. Desmin in Muscle and Associated Diseases: Beyond the Structural Function. Cell Tissue Res. 2015, 360, 591–608. [Google Scholar] [CrossRef]
- Watanabe, Y.; Yamaguchi, Y.; Komitsu, N.; Ohta, S.; Azuma, Y.; Izuhara, K.; Aihara, M. Elevation of Serum Squamous Cell Carcinoma Antigen 2 in Patients with Psoriasis: Associations with Disease Severity and Response to the Treatment. Br. J. Dermatol. 2016, 174, 1327–1336. [Google Scholar] [CrossRef]
- Ghonemy, S.; Mohamed, B.; Elkashishy, K.; Ibrahim, A.-S.M. Squamous Cell Carcinoma Antigen in Psoriasis: An Immunohistochemical Study. J. Clin. Aesthet. Dermatol. 2021, 14, 50–53. [Google Scholar]
- Szabo, R.; Netzel-Arnett, S.; Hobson, J.P.; Antalis, T.M.; Bugge, T.H. Matriptase-3 Is a Novel Phylogenetically Preserved Membrane-Anchored Serine Protease with Broad Serpin Reactivity. Biochem. J. 2005, 390, 231–242. [Google Scholar] [CrossRef]
- Castro-Ferreira, R.; Fontes-Carvalho, R.; Falcão-Pires, I.; Leite-Moreira, A.F. Papel Da Titina Na Modulação Da Função Cardíaca e Suas Implicações Fisiopatológicas. Arq. Bras. Cardiol. 2011, 96, 332–339. [Google Scholar] [CrossRef]
- Kellermayer, D.; Smith, J.E.; Granzier, H. Titin Mutations and Muscle Disease. Pflügers Arch.-Eur. J. Physiol. 2019, 471, 673–682. [Google Scholar] [CrossRef]
- Loescher, C.M.; Hobbach, A.J.; Linke, W.A. Titin (TTN): From Molecule to Modifications, Mechanics, and Medical Significance. Cardiovasc. Res. 2022, 118, 2903–2918. [Google Scholar] [CrossRef]
- Pinotsis, N.; Chatziefthimiou, S.D.; Berkemeier, F.; Beuron, F.; Mavridis, I.M.; Konarev, P.V.; Svergun, D.I.; Morris, E.; Rief, M.; Wilmanns, M. Superhelical Architecture of the Myosin Filament-Linking Protein Myomesin with Unusual Elastic Properties. PLoS Biol. 2012, 10, e1001261. [Google Scholar] [CrossRef]
- Pernigo, S.; Fukuzawa, A.; Beedle, A.E.M.; Holt, M.; Round, A.; Pandini, A.; Garcia-Manyes, S.; Gautel, M.; Steiner, R.A. Binding of Myomesin to Obscurin-Like-1 at the Muscle M-Band Provides a Strategy for Isoform-Specific Mechanical Protection. Structure 2017, 25, 107–120. [Google Scholar] [CrossRef]
- Hang, C.; Song, Y.; Li, Y.; Zhang, S.; Chang, Y.; Bai, R.; Saleem, A.; Jiang, M.; Lu, W.; Lan, F.; et al. Knockout of MYOM1 in Human Cardiomyocytes Leads to Myocardial Atrophy via Impairing Calcium Homeostasis. J. Cell. Mol. Med. 2021, 25, 1661–1676. [Google Scholar] [CrossRef]
- Blachly-Dyson, E.; Zambronicz, E.B.; Yu, W.H.; Adams, V.; McCabe, E.R.; Adelman, J.; Colombini, M.; Forte, M. Cloning and Functional Expression in Yeast of Two Human Isoforms of the Outer Mitochondrial Membrane Channel, the Voltage-Dependent Anion Channel. J. Biol. Chem. 1993, 268, 1835–1841. [Google Scholar] [CrossRef] [PubMed]
- Hiller, S.; Garces, R.G.; Malia, T.J.; Orekhov, V.Y.; Colombini, M.; Wagner, G. Solution Structure of the Integral Human Membrane Protein VDAC-1 in Detergent Micelles. Science (80-) 2008, 321, 1206–1210. [Google Scholar] [CrossRef] [PubMed]
- Dadsena, S.; Bockelmann, S.; Mina, J.G.M.; Hassan, D.G.; Korneev, S.; Razzera, G.; Jahn, H.; Niekamp, P.; Müller, D.; Schneider, M.; et al. Ceramides Bind VDAC2 to Trigger Mitochondrial Apoptosis. Nat. Commun. 2019, 10, 1832. [Google Scholar] [CrossRef] [PubMed]
- Wade, R.; Gunning, P.; Eddy, R.; Shows, T.; Kedes, L. Nucleotide Sequence, Tissue-Specific Expression, and Chromosome Location of Human Carbonic Anhydrase III: The Human CAIII Gene Is Located on the Same Chromosome as the Closely Linked CAI and CAII Genes. Proc. Natl. Acad. Sci. USA 1986, 83, 9571–9575. [Google Scholar] [CrossRef]
- Elder, I.; Fisher, Z.; Laipis, P.J.; Tu, C.; McKenna, R.; Silverman, D.N. Structural and Kinetic Analysis of Proton Shuttle Residues in the Active Site of Human Carbonic Anhydrase III. Proteins Struct. Funct. Bioinform. 2007, 68, 337–343. [Google Scholar] [CrossRef]
- Cheng, X.; Liu, D.; Song, H.; Tian, X.; Yan, C.; Han, Y. Overexpression of Kininogen-1 Aggravates Oxidative Stress and Mitochondrial Dysfunction in DOX-Induced Cardiotoxicity. Biochem. Biophys. Res. Commun. 2021, 550, 142–150. [Google Scholar] [CrossRef]
- Schroeder, H.W.; Cavacini, L. Structure and Function of Immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef]
- McHeyzer-Williams, M.; Okitsu, S.; Wang, N.; McHeyzer-Williams, L. Molecular Programming of B Cell Memory. Nat. Rev. Immunol. 2012, 12, 24–34. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Y.R.; Liu, K.; Yin, Z.; Liu, R.; Xia, Y.; Tan, L.; Yang, P.; Lee, J.-H.; Li, X.; et al. KAT2A Coupled with the α-KGDH Complex Acts as a Histone H3 Succinyltransferase. Nature 2017, 552, 273–277. [Google Scholar] [CrossRef]
- Remacha, L.; Pirman, D.; Mahoney, C.E.; Coloma, J.; Calsina, B.; Currás-Freixes, M.; Letón, R.; Torres-Pérez, R.; Richter, S.; Pita, G.; et al. Recurrent Germline DLST Mutations in Individuals with Multiple Pheochromocytomas and Paragangliomas. Am. J. Hum. Genet. 2019, 104, 651–664. [Google Scholar] [CrossRef]
- Baszanowska, W.; Misiura, M.; Oscilowska, I.; Palka, J.; Miltyk, W. Extracellular Prolidase (PEPD) Induces Anabolic Processes through EGFR, Β1-Integrin, and IGF-1R Signaling Pathways in an Experimental Model of Wounded Fibroblasts. Int. J. Mol. Sci. 2021, 22, 942. [Google Scholar] [CrossRef]
- Jiao, Q.; Bai, Y.; Akaike, T.; Takeshima, H.; Ishikawa, Y.; Minamisawa, S. Sarcalumenin Is Essential for Maintaining Cardiac Function during Endurance Exercise Training. Am. J. Physiol. Circ. Physiol. 2009, 297, H576–H582. [Google Scholar] [CrossRef] [PubMed]
- Conte, E.; Dinoi, G.; Imbrici, P.; De Luca, A.; Liantonio, A. Sarcoplasmic Reticulum Ca2+ Buffer Proteins: A Focus on the Yet-To-Be-Explored Role of Sarcalumenin in Skeletal Muscle Health and Disease. Cells 2023, 12, 715. [Google Scholar] [CrossRef]
- Fu, H.; Subramanian, R.R.; Masters, S.C. 14-3-3 Proteins: Structure, Function, and Regulation. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 617–647. [Google Scholar] [CrossRef] [PubMed]
- Mhawech, P. 14-3-3 Proteins—An Update. Cell Res. 2005, 15, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Kleppe, R.; Martinez, A.; Døskeland, S.O.; Haavik, J. The 14-3-3 Proteins in Regulation of Cellular Metabolism. Semin. Cell Dev. Biol. 2011, 22, 713–719. [Google Scholar] [CrossRef]
- Kaplan, A.; Bueno, M.; Fournier, A.E. Extracellular Functions of 14-3-3 Adaptor Proteins. Cell. Signal. 2017, 31, 26–30. [Google Scholar] [CrossRef]
- Munier, C.C.; Ottmann, C.; Perry, M.W.D. 14-3-3 Modulation of the Inflammatory Response. Pharmacol. Res. 2021, 163, 105236. [Google Scholar] [CrossRef]
- Low, F.M.; Hampton, M.B.; Winterbourn, C.C. Peroxiredoxin 2 and Peroxide Metabolism in the Erythrocyte. Antioxid. Redox Signal. 2008, 10, 1621–1630. [Google Scholar] [CrossRef]
- Liu, J.; Su, G.; Gao, J.; Tian, Y.; Liu, X.; Zhang, Z. Effects of Peroxiredoxin 2 in Neurological Disorders: A Review of Its Molecular Mechanisms. Neurochem. Res. 2020, 45, 720–730. [Google Scholar] [CrossRef]
- Talbert, P.B.; Henikoff, S. Histone Variants at a Glance. J. Cell Sci. 2021, 134, jcs244749. [Google Scholar] [CrossRef]
- Buttress, T.; He, S.; Wang, L.; Zhou, S.; Saalbach, G.; Vickers, M.; Li, G.; Li, P.; Feng, X. Histone H2B.8 Compacts Flowering Plant Sperm through Chromatin Phase Separation. Nature 2022, 611, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Read, J.A.; Winter, V.J.; Eszes, C.M.; Sessions, R.B.; Brady, R.L. Structural Basis for Altered Activity of M- and H-Isozyme Forms of Human Lactate Dehydrogenase. Proteins 2001, 43, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-J.; Mahieu, N.G.; Huang, X.; Singh, M.; Crawford, P.A.; Johnson, S.L.; Gross, R.W.; Schaefer, J.; Patti, G.J. Lactate Metabolism Is Associated with Mammalian Mitochondria. Nat. Chem. Biol. 2016, 12, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Xiong, Y.; Qiao, T.; Li, X.; Jia, L.; Han, Y. Lactate Dehydrogenase A: A Key Player in Carcinogenesis and Potential Target in Cancer Therapy. Cancer Med. 2018, 7, 6124–6136. [Google Scholar] [CrossRef]
- Urbańska, K.; Orzechowski, A. Unappreciated Role of LDHA and LDHB to Control Apoptosis and Autophagy in Tumor Cells. Int. J. Mol. Sci. 2019, 20, 2085. [Google Scholar] [CrossRef]
- Park, J.S.; Saeed, K.; Jo, M.H.; Kim, M.W.; Lee, H.J.; Park, C.-B.; Lee, G.; Kim, M.O. LDHB Deficiency Promotes Mitochondrial Dysfunction Mediated Oxidative Stress and Neurodegeneration in Adult Mouse Brain. Antioxidants 2022, 11, 261. [Google Scholar] [CrossRef]
- Munder, M. Suppression of T-Cell Functions by Human Granulocyte Arginase. Blood 2006, 108, 1627–1634. [Google Scholar] [CrossRef]
- Niu, F.; Yu, Y.; Li, Z.; Ren, Y.; Li, Z.; Ye, Q.; Liu, P.; Ji, C.; Qian, L.; Xiong, Y. Arginase: An Emerging and Promising Therapeutic Target for Cancer Treatment. Biomed. Pharmacother. 2022, 149, 112840. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, X.; Friesen, T.J.; Kwak, J.W.; Pisarenko, T.; Mekvanich, S.; Velasco, M.A.; Randolph, T.W.; Kargl, J.; Houghton, A.M. Annexin A2/TLR2/MYD88 Pathway Induces Arginase 1 Expression in Tumor-Associated Neutrophils. J. Clin. Investig. 2022, 132, e153643. [Google Scholar] [CrossRef]
- Stanton, R.C. Glucose-6-Phosphate Dehydrogenase, NADPH, and Cell Survival. IUBMB Life 2012, 64, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Z.; Hoshino, A.; Zheng, H.D.; Morley, M.; Arany, Z.; Rabinowitz, J.D. NADPH Production by the Oxidative Pentose-Phosphate Pathway Supports Folate Metabolism. Nat. Metab. 2019, 1, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Cano, M.; Datta, S.; Wang, L.; Liu, T.; Flores-Bellver, M.; Sachdeva, M.; Sinha, D.; Handa, J.T. Nrf2 Deficiency Decreases NADPH from Impaired IDH Shuttle and Pentose Phosphate Pathway in Retinal Pigmented Epithelial Cells to Magnify Oxidative Stress-induced Mitochondrial Dysfunction. Aging Cell 2021, 20, e13444. [Google Scholar] [CrossRef] [PubMed]
- Tian, N.; Hu, L.; Lu, Y.; Tong, L.; Feng, M.; Liu, Q.; Li, Y.; Zhu, Y.; Wu, L.; Ji, Y.; et al. TKT Maintains Intestinal ATP Production and Inhibits Apoptosis-Induced Colitis. Cell Death Dis. 2021, 12, 853. [Google Scholar] [CrossRef] [PubMed]
- Baptista, I.; Karakitsou, E.; Cazier, J.-B.; Günther, U.L.; Marin, S.; Cascante, M. TKTL1 Knockdown Impairs Hypoxia-Induced Glucose-6-Phosphate Dehydrogenase and Glyceraldehyde-3-Phosphate Dehydrogenase Overexpression. Int. J. Mol. Sci. 2022, 23, 3574. [Google Scholar] [CrossRef] [PubMed]
- Hammers, C.M.; Stanley, J.R. Desmoglein-1, Differentiation, and Disease. J. Clin. Investig. 2013, 123, 1419–1422. [Google Scholar] [CrossRef] [PubMed]
- Kasperkiewicz, M.; Ellebrecht, C.T.; Takahashi, H.; Yamagami, J.; Zillikens, D.; Payne, A.S.; Amagai, M. Pemphigus. Nat. Rev. Dis. Prim. 2017, 3, 17026. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, M.; Koetsier, J.L.; Huffine, A.L.; Broussard, J.A.; Godsel, B.M.; Cohen-Barak, E.; Sprecher, E.; Wolfgeher, D.J.; Kron, S.J.; Godsel, L.M.; et al. Epidermal Stratification Requires Retromer-Mediated Desmoglein-1 Recycling. Dev. Cell 2022, 57, 2683–2698. [Google Scholar] [CrossRef]
- Flix, B.; de la Torre, C.; Castillo, J.; Casal, C.; Illa, I.; Gallardo, E. Dysferlin Interacts with Calsequestrin-1, Myomesin-2 and Dynein in Human Skeletal Muscle. Int. J. Biochem. Cell Biol. 2013, 45, 1927–1938. [Google Scholar] [CrossRef]
- Sun, D.; Wu, R.; Zheng, J.; Li, P.; Yu, L. Polyubiquitin Chain-Induced P62 Phase Separation Drives Autophagic Cargo Segregation. Cell Res. 2018, 28, 405–415. [Google Scholar] [CrossRef]
- Han, S.-W.; Jung, B.-K.; Ryu, K.-Y. Regulation of Polyubiquitin Genes to Meet Cellular Ubiquitin Requirement. BMB Rep. 2021, 54, 189–195. [Google Scholar] [CrossRef]
- Dósa, A.; Csizmadia, T. The Role of K63-Linked Polyubiquitin in Several Types of Autophagy. Biol. Futur. 2022, 73, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Niehaus, J.Z.; Good, M.; Jackson, L.E.; Ozolek, J.A.; Silverman, G.A.; Luke, C.J. Human SERPINB12 Is an Abundant Intracellular Serpin Expressed in Most Surface and Glandular Epithelia. J. Histochem. Cytochem. 2015, 63, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Rosa, J.-P.; Raslova, H.; Bryckaert, M. Filamin A: Key Actor in Platelet Biology. Blood 2019, 134, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Verdonschot, J.A.J.; Vanhoutte, E.K.; Claes, G.R.F.; Helderman-van den Enden, A.T.J.M.; Hoeijmakers, J.G.J.; Hellebrekers, D.M.E.I.; Haan, A.; Christiaans, I.; Lekanne Deprez, R.H.; Boen, H.M.; et al. A Mutation Update for the FLNC Gene in Myopathies and Cardiomyopathies. Hum. Mutat. 2020, 41, 1091–1111. [Google Scholar] [CrossRef]
- Yuen, M.; Ottenheijm, C.A.C. Nebulin: Big Protein with Big Responsibilities. J. Muscle Res. Cell Motil. 2020, 41, 103–124. [Google Scholar] [CrossRef]
- Wang, Z.; Grange, M.; Pospich, S.; Wagner, T.; Kho, A.L.; Gautel, M.; Raunser, S. Structures from Intact Myofibrils Reveal Mechanism of Thin Filament Regulation through Nebulin. Science (80-) 2022, 375. [Google Scholar] [CrossRef]
- Oi, N.; Yamamoto, H.; Langfald, A.; Bai, R.; Lee, M.-H.; Bode, A.M.; Dong, Z. LTA4H Regulates Cell Cycle and Skin Carcinogenesis. Carcinogenesis 2017, 38, 728–737. [Google Scholar] [CrossRef]
- Nordio, L.; Bazzocchi, C.; Genova, F.; Serra, V.; Longeri, M.; Franzo, G.; Rondena, M.; Stefanello, D.; Giudice, C. Molecular and Immunohistochemical Expression of LTA4H and FXR1 in Canine Oral Melanoma. Front. Vet. Sci. 2021, 8, 767887. [Google Scholar] [CrossRef]
- Lisowska-Myjak, B.; Jóźwiak-Kisielewska, A.; Łukaszkiewicz, J.; Skarżyńska, E. Vitamin D-Binding Protein as a Biomarker to Confirm Specific Clinical Diagnoses. Expert Rev. Mol. Diagn. 2020, 20, 49–56. [Google Scholar] [CrossRef]
- Wang, J.; Li, R.; Li, M.; Wang, C. Fibronectin and Colorectal Cancer: Signaling Pathways and Clinical Implications. J. Recept. Signal Transduct. 2021, 41, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ul Ain, Q.; Schulz, C.; Pircher, J. Role of Antimicrobial Peptide Cathelicidin in Thrombosis and Thromboinflammation. Front. Immunol. 2023, 14, 1151926. [Google Scholar] [CrossRef] [PubMed]
- Patten, J.; Wang, K. Fibronectin in Development and Wound Healing. Adv. Drug Deliv. Rev. 2021, 170, 353–368. [Google Scholar] [CrossRef]
- Dallacasagrande, V.; Hajjar, K.A. Annexin A2 in Inflammation and Host Defense. Cells 2020, 9, 1499. [Google Scholar] [CrossRef]
- Lim, H.I.; Hajjar, K.A. Annexin A2 in Fibrinolysis, Inflammation and Fibrosis. Int. J. Mol. Sci. 2021, 22, 6836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, Z.; Xia, S.; Yao, L.; Li, L.; Gan, Z.; Tang, H.; Guo, Q.; Yan, X.; Sun, Z. GDI2 Is a Novel Diagnostic and Prognostic Biomarker in Hepatocellular Carcinoma. Aging 2021, 13, 25304–25324. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, D.; Chen, G.-Y. Targeted Disruption of Gdi2 Causes Early Embryonic Lethality. Placenta 2022, 126, 17–25. [Google Scholar] [CrossRef]
- Punwani, D.; Pelz, B.; Yu, J.; Arva, N.C.; Schafernak, K.; Kondratowicz, K.; Makhija, M.; Puck, J.M. Coronin-1A: Immune Deficiency in Humans and Mice. J. Clin. Immunol. 2015, 35, 100–107. [Google Scholar] [CrossRef]
- Martorella, M.; Barford, K.; Winckler, B.; Deppmann, C.D. Emergent Role of Coronin-1a in Neuronal Signaling. Vitam. Horm. 2017, 104, 113–131. [Google Scholar]
- Tang, X.; Li, Q.; Li, L.; Jiang, J. Expression of Talin-1 in Endometriosis and Its Possible Role in Pathogenesis. Reprod. Biol. Endocrinol. 2021, 19, 42. [Google Scholar] [CrossRef]
- Zhao, Y.; Lykov, N.; Tzeng, C. Talin-1 Interaction Network in Cellular Mechanotransduction (Review). Int. J. Mol. Med. 2022, 49, 60. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Zhu, L.; Bromberger, T.; Yang, J.; Yang, Q.; Liu, J.; Plow, E.F.; Moser, M.; Qin, J. Mechanism of Integrin Activation by Talin and Its Cooperation with Kindlin. Nat. Commun. 2022, 13, 2362. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fold Change | |||
|---|---|---|---|
| Protein Description | Access Code | LAAO/Control | Control/LAAO |
| Alpha-actinin-2 | F6THM6 | 4.0756 | - |
| Myomesin-2 | P54296 | 3.3955 | - |
| ATP synthase subunit alpha, mitochondrial | P25705 | 2.9427 | - |
| Myosin-2 | Q9UKX2 | 2.8325 | - |
| Alpha-actinin-2 | P35609 | 2.8138 | - |
| Myosin-1 | P12882 | 2.7773 | - |
| Myosin-binding protein C, slow-type | F8VZY0 | 2.6963 | - |
| Myosin-6 | P13533 | 2.6811 | - |
| Myosin-7 | P12883 | 2.6417 | - |
| ATP synthase subunit beta, mitochondrial | P06576 | 2.5931 | - |
| Myosin-8 | P13535 | 2.5761 | - |
| Myosin-4 | Q9Y623 | 2.5727 | - |
| Alpha-actinin-3 | A0A087WSZ2 | 2.4419 | - |
| Myosin light chain 1/3, skeletal muscle isoform | P05976 | 2.3077 | - |
| Myosin-13 | Q9UKX3 | 2.2757 | - |
| ADP/ATP translocase 1 | P12235 | 2.2577 | - |
| Sarcoplasmic/endoplasmic reticulum calcium ATPase 2 | H7C5W9 | 2.0930 | - |
| Putative elongation factor 1-alpha-like 3 | Q5VTE0 | - | 2.2892 |
| Lactotransferrin | P02788 | - | 2.3342 |
| Coronin-1A | P31146 | - | 2.6324 |
| Myeloperoxidase | P05164 | - | 2.866 |
| Complement C3 | P01024 | - | 2.9114 |
| Immunoglobulin heavy constant gamma 1 | A0A0A0MS08 | - | 2.9392 |
| Tubulin beta-6 chain | Q9BUF5 | - | 3.4374 |
| Myosin-9 | P35579 | - | 3.5126 |
| Myeloblastin | P24158 | - | 3.5948 |
| Inter-alpha-trypsin inhibitor heavy chain H3 | Q06033 | - | 4.0529 |
| Triosephosphate isomerase | P60174 | - | 4.2686 |
| Glycogen phosphorylase, liver form | P06737 | - | 4.367 |
| Tubulin beta-4A chain | P04350 | - | 5.1224 |
| Transitional endoplasmic reticulum ATPase | P55072 | - | 5.5079 |
| Cathepsin G | P08311 | - | 5.5802 |
| Tubulin beta-4B chain | P68371 | - | 5.9516 |
| Tubulin beta chain | P07437 | - | 6.5033 |
| Tubulin alpha chain | F5H5D3 | - | 6.8008 |
| Filamin-A | P21333 | - | 7.1256 |
| Apolipoprotein B-100 | P04114 | - | 8.8368 |
| Tubulin alpha-4A chain | P68366 | - | 9.317 |
| Pigment epithelium-derived factor | P36955 | - | 9.9446 |
| Talin-1 | Q9Y490 | - | 12.962 |
| Adenylyl cyclase-associated protein 1 | Q01518 | - | 13.314 |
| Protein | Mechanism | Reference |
|---|---|---|
| Glucose-6-phosphate1-dehydrogenase; 6-phosphogluconate dehydrogenase; glucose-6-phosphate isomerase; transketolase. | Involved in the pentose phosphate pathway and plays an important role in the generation of NADPH. | [108,109,110,111,112] |
| desmoglein-1 | Key role in the structure and function of epithelial cells, like cell–cell adhesion. | [113,114,115] |
| Myomesin 2 | Formation and maintenance of the structure of the sarcomere. | [67,116] |
| Polyubiquitin C | Directly related to protein degradation through the proteasome. | [117,118,119] |
| Serpine B12 | A protein involved in the downregulation of endopeptidase activity, providing protection to epithelial cells. | [120] |
| Filamin C | Organization and stabilization of the cytoskeleton, mainly in muscle cells. | [121,122] |
| Nebulin | Acts mainly in the regulation of muscle structure and function, it is crucial for the proper functioning of skeletal muscles. | [123,124] |
| Leukotriene A4 hydrolase | Plays a central role in the synthesis of leukotrienes, which are an important inflammatory mediator for immune response. | [125,126] |
| Vitamin D binding protein | Plays a crucial role in the transport and regulation of vitamin D in the body. | [127] |
| Cathelicidin antimicrobial peptide | Plays a key role in host defense against microbial infections, including bacteria, viruses, fungi, and even parasites. | [128,129] |
| Fibronectin | Plays multiple roles in cell adhesion, migration, wound healing, embryonic development, and immune response, produced by various cells, including fibroblasts, hepatocytes, endothelial cells, and muscle cells. | [128,130] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrath, S.N.; Pontes, A.S.; Paloschi, M.V.; Silva, M.D.S.; Lopes, J.A.; Boeno, C.N.; Silva, C.P.; Santana, H.M.; Cardozo, D.G.; Ugarte, A.V.E.; et al. Exosome Liberation by Human Neutrophils under L-Amino Acid Oxidase of Calloselasma rhodostoma Venom Action. Toxins 2023, 15, 625. https://doi.org/10.3390/toxins15110625
Serrath SN, Pontes AS, Paloschi MV, Silva MDS, Lopes JA, Boeno CN, Silva CP, Santana HM, Cardozo DG, Ugarte AVE, et al. Exosome Liberation by Human Neutrophils under L-Amino Acid Oxidase of Calloselasma rhodostoma Venom Action. Toxins. 2023; 15(11):625. https://doi.org/10.3390/toxins15110625
Chicago/Turabian StyleSerrath, Suzanne N., Adriana S. Pontes, Mauro V. Paloschi, Milena D. S. Silva, Jéssica A. Lopes, Charles N. Boeno, Carolina P. Silva, Hallison M. Santana, Daniel G. Cardozo, Andrey V. E. Ugarte, and et al. 2023. "Exosome Liberation by Human Neutrophils under L-Amino Acid Oxidase of Calloselasma rhodostoma Venom Action" Toxins 15, no. 11: 625. https://doi.org/10.3390/toxins15110625
APA StyleSerrath, S. N., Pontes, A. S., Paloschi, M. V., Silva, M. D. S., Lopes, J. A., Boeno, C. N., Silva, C. P., Santana, H. M., Cardozo, D. G., Ugarte, A. V. E., Magalhães, J. G. S., Cruz, L. F., Setubal, S. S., Soares, A. M., Cavecci-Mendonça, B., Santos, L. D., & Zuliani, J. P. (2023). Exosome Liberation by Human Neutrophils under L-Amino Acid Oxidase of Calloselasma rhodostoma Venom Action. Toxins, 15(11), 625. https://doi.org/10.3390/toxins15110625