Harnessing the Power of Venomous Animal-Derived Toxins against COVID-19

 ,
,  ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
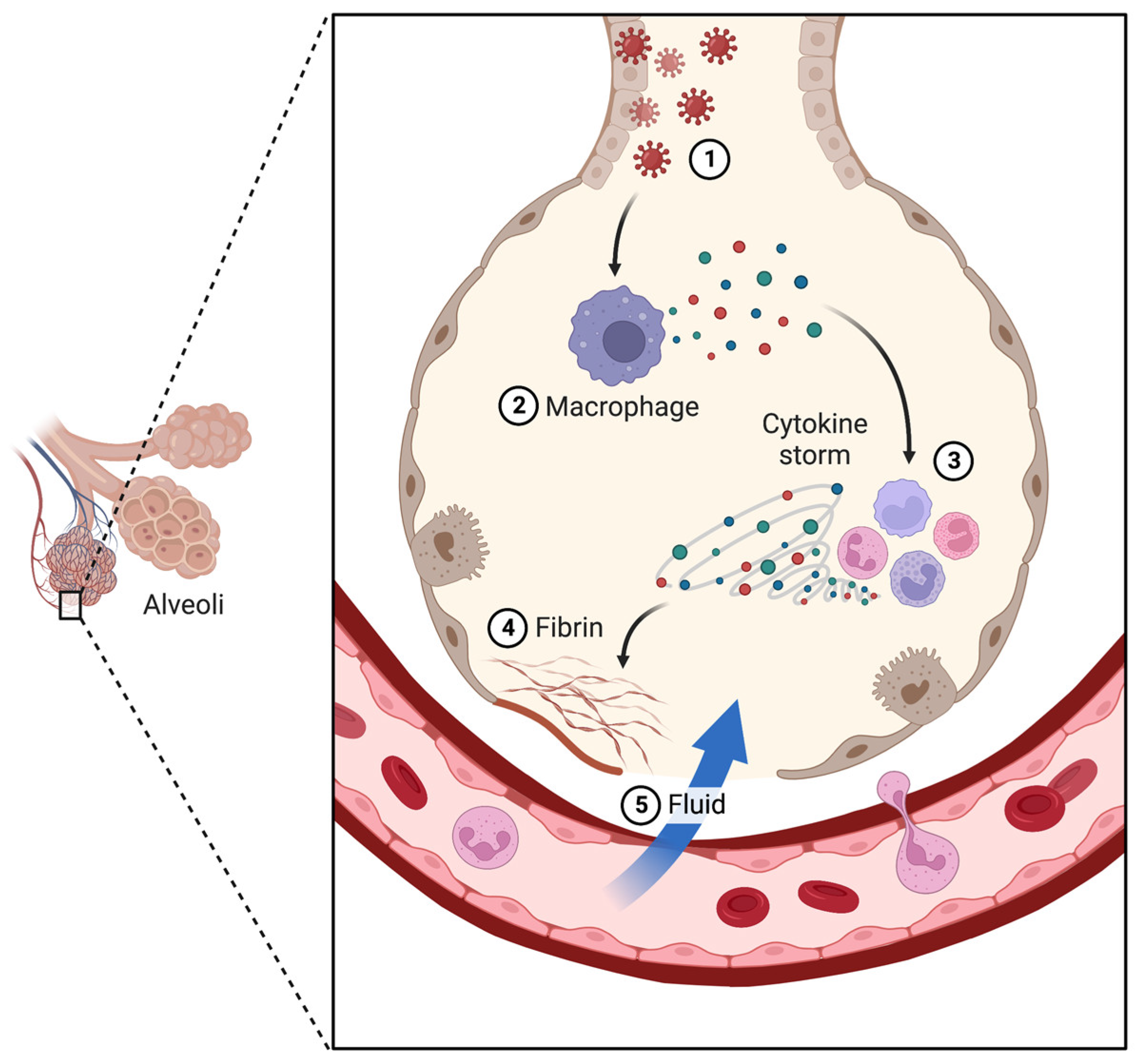
2. COVID-19 Disease
3. Available Treatments for COVID-19
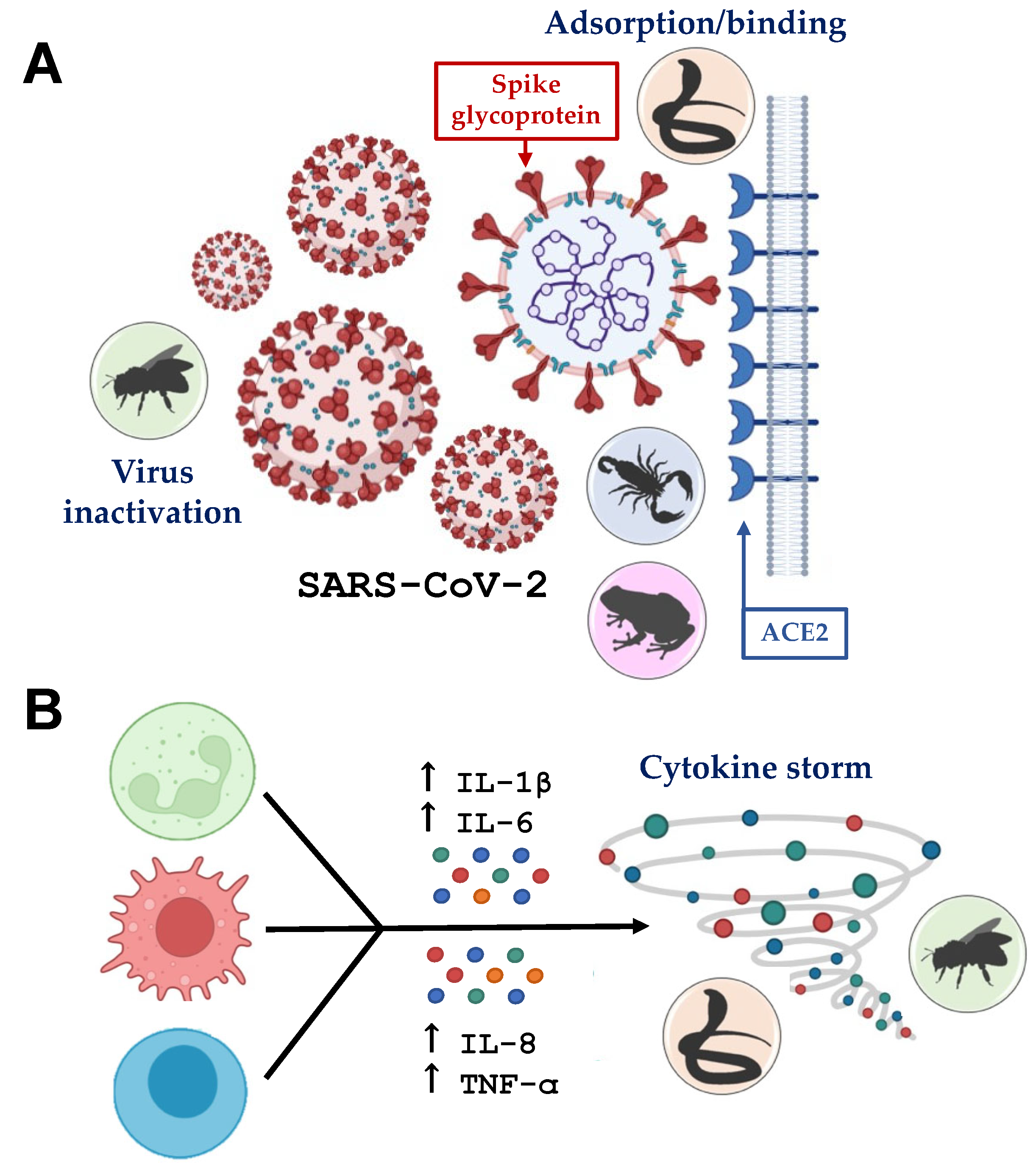
4. Crosstalk of Animal-Derived Toxins and COVID-19

{kind=link}
{kind=link}
| Toxins | Species | Animal | Mechanism | Year | Ref. |
|---|---|---|---|---|---|
| Bee venom | Apis mellifera | Bee | Hypothesis: attenuate cytokine storm caused by SARS-CoV-2. | 2020 | [71,72] |
| Prophylactic context for COVID-19. | |||||
| Melittin | Apis mellifera | Bee | In vitro assay using VERO cells: neutralizes the SARS-CoV-2 virus. | 2022 | [73] |
| Dermaseptin-S9 | Phyllomedusa sauvagii | Frog | Inhibitor of SARS-CoV-2 spike glycoprotein by protein-peptide analysis in silico by docking. | 2020 | [75] |
| Meucin18 and its mutation | Mesobuthus eupeus | Scorpion | Inhibitor of SARS-CoV-2 spike glycoprotein. Protein-peptide analysis in silico by docking. | 2021 | [76] |
| Cobrotoxin | Naja naja atra | Snake | Inhibitory effect on the cytokine storm caused by SARS-CoV-2 in COVID-19. | 2020 | [70] |
| Dimeric peptides from BthTX-I (PLA2) | Bothrops jararacussu | Snake | Inhibitory activity against the Papain-like protease of SARS-CoV-2. | 2021 | [69] |
| PLA2 | Vipera nikolskii | Snake | Inhibition of SARS-CoV-2 spike glycoprotein-mediated cell-cell infusion. | 2021 | [68] |
5. Venom-Derived Toxins as COVID-19 Therapy: New Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Utkin, Y.; Siniavin, A.; Kasheverov, I.; Tsetlin, V. Antiviral Effects of Animal Toxins: Is There a Way to Drugs? Int. J. Mol. Sci. 2022, 23, 73634. [Google Scholar] [CrossRef]
- Govender, R.D.; Hashim, M.J.; Khan, M.A.; Mustafa, H.; Khan, G. Global Epidemiology of HIV/AIDS: A Resurgence in North America and Europe. JEGH 2021, 11, 296. [Google Scholar] [CrossRef]
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 17 January 2023).
- Shariare, M.H.; Parvez, M.A.K.; Karikas, G.A.; Kazi, M. The Growing Complexity of COVID-19 Drug and Vaccine Candidates: Challenges and Critical Transitions. J. Infect. Public Health 2021, 14, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Herzig, V.; Cristofori-Armstrong, B.; Israel, M.R.; Nixon, S.A.; Vetter, I.; King, G.F. Animal Toxins—Nature’s Evolutionary-Refined Toolkit for Basic Research and Drug Discovery. Biochem. Pharmacol. 2020, 181, 114096. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Cerni, F.A.; Pinheiro-Junior, E.L.; Zoccal, K.F.; Bordon, K.d.C.F.; Amorim, F.G.; Peigneur, S.; Vriens, K.; Thevissen, K.; Cammue, B.P.A.; et al. Non-Disulfide-Bridged Peptides from Tityus Serrulatus Venom: Evidence for Proline-Free ACE-Inhibitors. Peptides 2016, 82, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Knerr, J.M.; Argemi, L.; Bordon, K.C.F.; Pucca, M.B.; Cerni, F.A.; Arantes, E.C.; Çalışkan, F.; Laustsen, A.H. Scorpion Venom: Detriments and Benefits. Biomedicines 2020, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Ahmadi, S.; Cerni, F.A.; Ledsgaard, L.; Sørensen, C.V.; McGeoghan, F.T.S.; Stewart, T.; Schoof, E.; Lomonte, B.; auf dem Keller, U.; et al. Unity Makes Strength: Exploring Intraspecies and Interspecies Toxin Synergism between Phospholipases A2 and Cytotoxins. Front. Pharmacol. 2020, 11, 611. [Google Scholar] [CrossRef]
- Da Mata, É.C.G.; Mourão, C.B.F.; Rangel, M.; Schwartz, E.F. Antiviral Activity of Animal Venom Peptides and Related Compounds. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 3. [Google Scholar] [CrossRef]
- Vilas Boas, L.C.P.; Campos, M.L.; Berlanda, R.L.A.; de Carvalho Neves, N.; Franco, O.L. Antiviral Peptides as Promising Therapeutic Drugs. Cell. Mol. Life Sci. 2019, 76, 3525–3542. [Google Scholar] [CrossRef]
- Sample, C.J.; Hudak, K.E.; Barefoot, B.E.; Koci, M.D.; Wanyonyi, M.S.; Abraham, S.; Staats, H.F.; Ramsburg, E.A. A Mastoparan-Derived Peptide Has Broad-Spectrum Antiviral Activity against Enveloped Viruses. Peptides 2013, 48, 96–105. [Google Scholar] [CrossRef]
- Chen, N.; Xu, S.; Zhang, Y.; Wang, F. Animal Protein Toxins: Origins and Therapeutic Applications. Biophys. Rep. 2018, 4, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Fauci, A.S.; Lane, H.C.; Redfield, R.R. Covid-19—Navigating the Uncharted. N. Engl. J. Med. 2020, 382, 1268–1269. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus–Infected Pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The Species Severe Acute Respiratory Syndrome-Related Coronavirus: Classifying 2019-NCoV and Naming It SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- Holshue, M.L.; DeBolt, C.; Lindquist, S.; Lofy, K.H.; Wiesman, J.; Bruce, H.; Spitters, C.; Ericson, K.; Wilkerson, S.; Tural, A.; et al. First Case of 2019 Novel Coronavirus in the United States. N. Engl. J. Med. 2020, 382, 929–936. [Google Scholar] [CrossRef]
- Tarighi, P.; Eftekhari, S.; Chizari, M.; Sabernavaei, M.; Jafari, D.; Mirzabeigi, P. A Review of Potential Suggested Drugs for Coronavirus Disease (COVID-19) Treatment. Eur. J. Pharmacol. 2021, 895, 173890. [Google Scholar] [CrossRef]
- Ko, W.-C.; Rolain, J.-M.; Lee, N.-Y.; Chen, P.-L.; Huang, C.-T.; Lee, P.-I.; Hsueh, P.-R. Arguments in Favour of Remdesivir for Treating SARS-CoV-2 Infections. Int. J. Antimicrob. Agents 2020, 55, 105933. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, Z.; Chen, W.; Cai, Z.; Ge, X.; Zhu, H.; Jiang, T.; Tan, W.; Peng, Y. Predicting the Receptor-Binding Domain Usage of the Coronavirus Based on Kmer Frequency on Spike Protein. Infect. Genet. Evol. 2018, 61, 183–184. [Google Scholar] [CrossRef]
- Yang, J.; Petitjean, S.J.L.; Koehler, M.; Zhang, Q.; Dumitru, A.C.; Chen, W.; Derclaye, S.; Vincent, S.P.; Soumillion, P.; Alsteens, D. Molecular Interaction and Inhibition of SARS-CoV-2 Binding to the ACE2 Receptor. Nat. Commun. 2020, 11, 4541. [Google Scholar] [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Zhou, S.; Graham, R.L.; Pruijssers, A.J.; Agostini, M.L.; Leist, S.R.; Schäfer, A.; Dinnon, K.H.; Stevens, L.J.; et al. An Orally Bioavailable Broad-Spectrum Antiviral Inhibits SARS-CoV-2 in Human Airway Epithelial Cell Cultures and Multiple Coronaviruses in Mice. Sci. Transl. Med. 2020, 12, eabb5883. [Google Scholar] [CrossRef]
- Dömling, A.; Gao, L. Chemistry and Biology of SARS-CoV-2. Chem 2020, 6, 1283–1295. [Google Scholar] [CrossRef]
- Da Silva, S.J.R.; Alves da Silva, C.T.; Mendes, R.P.G.; Pena, L. Role of Nonstructural Proteins in the Pathogenesis of SARS-CoV-2. J. Med. Virol. 2020, 92, 1427–1429. [Google Scholar] [CrossRef]
- Shin, D.; Mukherjee, R.; Grewe, D.; Bojkova, D.; Baek, K.; Bhattacharya, A.; Schulz, L.; Widera, M.; Mehdipour, A.R.; Tascher, G.; et al. Papain-like Protease Regulates SARS-CoV-2 Viral Spread and Innate Immunity. Nature 2020, 587, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Cascella, M.; Rajnik, M.; Aleem, A.; Dulebohn, S.C.; Di Napoli, R. Features, Evaluation, and Treatment of Coronavirus (COVID-19). In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Gao, Y.-M.; Xu, G.; Wang, B.; Liu, B.-C. Cytokine Storm Syndrome in Coronavirus Disease 2019: A Narrative Review. J. Intern. Med. 2021, 289, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological Findings of COVID-19 Associated with Acute Respiratory Distress Syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Caricchio, R.; Gallucci, M.; Dass, C.; Zhang, X.; Gallucci, S.; Fleece, D.; Bromberg, M.; Criner, G.J.; Temple University COVID-19 Research Group. Preliminary Predictive Criteria for COVID-19 Cytokine Storm. Ann. Rheum. Dis. 2021, 80, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.J.; Peltan, I.D.; Jensen, P.; Hoda, D.; Hunter, B.; Silver, A.; Starr, N.; Buckel, W.; Grisel, N.; Hummel, E.; et al. Clinical Criteria for COVID-19-Associated Hyperinflammatory Syndrome: A Cohort Study. Lancet Rheumatol. 2020, 2, e754–e763. [Google Scholar] [CrossRef]
- Chen, L.Y.C.; Quach, T.T.T. COVID-19 Cytokine Storm Syndrome: A Threshold Concept. Lancet Microbe 2021, 2, e49–e50. [Google Scholar] [CrossRef]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and Immunological Features of Severe and Moderate Coronavirus Disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef]
- Coopersmith, C.M.; Antonelli, M.; Bauer, S.R.; Deutschman, C.S.; Evans, L.E.; Ferrer, R.; Hellman, J.; Jog, S.; Kesecioglu, J.; Kissoon, N.; et al. The Surviving Sepsis Campaign: Research Priorities for Coronavirus Disease 2019 in Critical Illness. Crit. Care Med. 2021, 49, 598–622. [Google Scholar] [CrossRef]
- Gandhi, R.T.; Lynch, J.B.; Del Rio, C. Mild or Moderate COVID-19. N. Engl. J. Med. 2020, 383, 1757–1766. [Google Scholar] [CrossRef] [PubMed]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-Approved Drug Ivermectin Inhibits the Replication of SARS-CoV-2 in Vitro. Antivir. Res. 2020, 178, 104787. [Google Scholar] [CrossRef] [PubMed]
- Khuroo, M.S. Chloroquine and Hydroxychloroquine in Coronavirus Disease 2019 (COVID-19). Facts, Fiction and the Hype: A Critical Appraisal. Int. J. Antimicrob. Agents 2020, 56, 106101. [Google Scholar] [CrossRef] [PubMed]
- The RECOVERY Collaborative Group Effect of Hydroxychloroquine in Hospitalized Patients with COVID-19. N. Engl. J. Med. 2020, 383, 2030–2040. [CrossRef]
- Zhang, R.; Mylonakis, E. In Inpatients with COVID-19, None of Remdesivir, Hydroxychloroquine, Lopinavir, or Interferon β-1a Differed from Standard Care for in-Hospital Mortality. Ann. Intern. Med. 2021, 174, JC17. [Google Scholar] [CrossRef]
- Mitjà, O.; Corbacho-Monné, M.; Ubals, M.; Tebé, C.; Peñafiel, J.; Tobias, A.; Ballana, E.; Alemany, A.; Riera-Martí, N.; Pérez, C.A.; et al. Hydroxychloroquine for Early Treatment of Adults with Mild Coronavirus Disease 2019: A Randomized, Controlled Trial. Clin. Infect. Dis. 2021, 73, e4073–e4081. [Google Scholar] [CrossRef]
- Boulware, D.R.; Pullen, M.F.; Bangdiwala, A.S.; Pastick, K.A.; Lofgren, S.M.; Okafor, E.C.; Skipper, C.P.; Nascene, A.A.; Nicol, M.R.; Abassi, M.; et al. A Randomized Trial of Hydroxychloroquine as Postexposure Prophylaxis for COVID-19. N. Engl. J. Med. 2020, 383, 517–525. [Google Scholar] [CrossRef]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir–Ritonavir in Adults Hospitalized with Severe COVID-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, A.; Singh, R.; Misra, A. Molnupiravir in COVID-19: A Systematic Review of Literature. Diabetes Metab. Syndr. 2021, 15, 102329. [Google Scholar] [CrossRef]
- Jayk Bernal, A.; Gomes da Silva, M.M.; Musungaie, D.B.; Kovalchuk, E.; Gonzalez, A.; Delos Reyes, V.; Martín-Quirós, A.; Caraco, Y.; Williams-Diaz, A.; Brown, M.L.; et al. Molnupiravir for Oral Treatment of COVID-19 in Nonhospitalized Patients. N. Engl. J. Med. 2022, 386, 509–520. [Google Scholar] [CrossRef]
- Mahase, E. COVID-19: Pfizer’s Paxlovid Is 89% Effective in Patients at Risk of Serious Illness, Company Reports. BMJ 2021, 375, n2713. [Google Scholar] [CrossRef] [PubMed]
- Goldman, J.D.; Lye, D.C.B.; Hui, D.S.; Marks, K.M.; Bruno, R.; Montejano, R.; Spinner, C.D.; Galli, M.; Ahn, M.-Y.; Nahass, R.G.; et al. Remdesivir for 5 or 10 Days in Patients with Severe COVID-19. N. Engl. J. Med. 2020, 383, 1827–1837. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, R.L.; Vaca, C.E.; Paredes, R.; Mera, J.; Webb, B.J.; Perez, G.; Oguchi, G.; Ryan, P.; Nielsen, B.U.; Brown, M.; et al. Early Remdesivir to Prevent Progression to Severe COVID-19 in Outpatients. N. Engl. J. Med. 2022, 386, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Spinner, C.D.; Gottlieb, R.L.; Criner, G.J.; Arribas López, J.R.; Cattelan, A.M.; Soriano Viladomiu, A.; Ogbuagu, O.; Malhotra, P.; Mullane, K.M.; Castagna, A.; et al. Effect of Remdesivir vs Standard Care on Clinical Status at 11 Days in Patients with Moderate COVID-19: A Randomized Clinical Trial. JAMA 2020, 324, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the Treatment of COVID-19-Final Report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef]
- Joyner, M.J.; Bruno, K.A.; Klassen, S.A.; Kunze, K.L.; Johnson, P.W.; Lesser, E.R.; Wiggins, C.C.; Senefeld, J.W.; Klompas, A.M.; Hodge, D.O.; et al. Safety Update: COVID-19 Convalescent Plasma in 20,000 Hospitalized Patients. Mayo Clin. Proc. 2020, 95, 1888–1897. [Google Scholar] [CrossRef]
- Simonovich, V.A.; Burgos Pratx, L.D.; Scibona, P.; Beruto, M.V.; Vallone, M.G.; Vázquez, C.; Savoy, N.; Giunta, D.H.; Pérez, L.G.; Sánchez, M.D.L.; et al. A Randomized Trial of Convalescent Plasma in COVID-19 Severe Pneumonia. N. Engl. J. Med. 2021, 384, 619–629. [Google Scholar] [CrossRef]
- Weinreich, D.M.; Sivapalasingam, S.; Norton, T.; Ali, S.; Gao, H.; Bhore, R.; Musser, B.J.; Soo, Y.; Rofail, D.; Im, J.; et al. REGN-COV2, a Neutralizing Antibody Cocktail, in Outpatients with COVID-19. N. Engl. J. Med. 2021, 384, 238–251. [Google Scholar] [CrossRef]
- Wong, S.K.; Li, W.; Moore, M.J.; Choe, H.; Farzan, M. A 193-Amino Acid Fragment of the SARS Coronavirus S Protein Efficiently Binds Angiotensin-Converting Enzyme 2. J. Biol. Chem. 2004, 279, 3197–3201. [Google Scholar] [CrossRef] [PubMed]
- Van den Brink, E.N.; Ter Meulen, J.; Cox, F.; Jongeneelen, M.A.C.; Thijsse, A.; Throsby, M.; Marissen, W.E.; Rood, P.M.L.; Bakker, A.B.H.; Gelderblom, H.R.; et al. Molecular and Biological Characterization of Human Monoclonal Antibodies Binding to the Spike and Nucleocapsid Proteins of Severe Acute Respiratory Syndrome Coronavirus. J. Virol. 2005, 79, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Yan, X.; Guo, X.; Cao, W.; Han, W.; Qi, C.; Feng, J.; Yang, D.; Gao, G.; Jin, G. A Human SARS-CoV Neutralizing Antibody against Epitope on S2 Protein. Biochem. Biophys. Res. Commun. 2005, 333, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Jahanshahlu, L.; Rezaei, N. Monoclonal Antibody as a Potential Anti-COVID-19. Biomed. Pharmacother. 2020, 129, 110337. [Google Scholar] [CrossRef]
- Wang, C.; Li, W.; Drabek, D.; Okba, N.M.A.; van Haperen, R.; Osterhaus, A.D.M.E.; van Kuppeveld, F.J.M.; Haagmans, B.L.; Grosveld, F.; Bosch, B.-J. A Human Monoclonal Antibody Blocking SARS-CoV-2 Infection. Nat. Commun. 2020, 11, 2251. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, F.; Shen, C.; Peng, W.; Li, D.; Zhao, C.; Li, Z.; Li, S.; Bi, Y.; Yang, Y.; et al. A Noncompeting Pair of Human Neutralizing Antibodies Block COVID-19 Virus Binding to Its Receptor ACE2. Science 2020, 368, 1274–1278. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 MRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cárdenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against COVID-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and Efficacy of the ChAdOx1 NCoV-19 Vaccine (AZD1222) against SARS-CoV-2: An Interim Analysis of Four Randomised Controlled Trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Shinde, V.; Bhikha, S.; Hoosain, Z.; Archary, M.; Bhorat, Q.; Fairlie, L.; Lalloo, U.; Masilela, M.S.L.; Moodley, D.; Hanley, S.; et al. Efficacy of NVX-CoV2373 COVID-19 Vaccine against the B.1.351 Variant. N. Engl. J. Med. 2021, 384, 1899–1909. [Google Scholar] [CrossRef]
- Dunkle, L.M.; Kotloff, K.L.; Gay, C.L.; Áñez, G.; Adelglass, J.M.; Barrat Hernández, A.Q.; Harper, W.L.; Duncanson, D.M.; McArthur, M.A.; Florescu, D.F.; et al. Efficacy and Safety of NVX-CoV2373 in Adults in the United States and Mexico. N. Engl. J. Med. 2022, 386, 531–543. [Google Scholar] [CrossRef]
- Bordon, K.d.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef]
- Cecilio, A.B.; Caldas, S.; Oliveira, R.A.D.; Santos, A.S.B.; Richardson, M.; Naumann, G.B.; Schneider, F.S.; Alvarenga, V.G.; Estevão-Costa, M.I.; Fuly, A.L.; et al. Molecular Characterization of Lys49 and Asp49 Phospholipases A2 from Snake Venom and Their Antiviral Activities against Dengue Virus. Toxins 2013, 5, 1780–1798. [Google Scholar] [CrossRef] [PubMed]
- Fenard, D.; Lambeau, G.; Valentin, E.; Lefebvre, J.C.; Lazdunski, M.; Doglio, A. Secreted Phospholipases A(2), a New Class of HIV Inhibitors That Block Virus Entry into Host Cells. J. Clin. Investig. 1999, 104, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhao, Z.; Zhou, D.; Chen, Y.; Hong, W.; Cao, L.; Yang, J.; Zhang, Y.; Shi, W.; Cao, Z.; et al. Virucidal Activity of a Scorpion Venom Peptide Variant Mucroporin-M1 against Measles, SARS-CoV and Influenza H5N1 Viruses. Peptides 2011, 32, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Ogando, N.S.; Dalebout, T.J.; Zevenhoven-Dobbe, J.C.; Limpens, R.W.A.L.; van der Meer, Y.; Caly, L.; Druce, J.; de Vries, J.J.C.; Kikkert, M.; Bárcena, M.; et al. SARS-Coronavirus-2 Replication in Vero E6 Cells: Replication Kinetics, Rapid Adaptation and Cytopathology. J. Gen. Virol. 2020, 101, 925–940. [Google Scholar] [CrossRef] [PubMed]
- Ae, S.; Ma, S.; Ma, N.; Ds, K.; Sd, G.; Va, G.; Va, M.; Vg, S.; Av, O.; Scr, L.; et al. Snake Venom Phospholipase A 2 s Exhibit Strong Virucidal Activity against SARS-CoV-2 and Inhibit the Viral Spike Glycoprotein Interaction with ACE2. Cell. Mol. Life Sci. CMLS 2021, 78, 7777–7794. [Google Scholar] [CrossRef]
- Freire, M.C.L.C.; Noske, G.D.; Bitencourt, N.V.; Sanches, P.R.S.; Santos-Filho, N.A.; Gawriljuk, V.O.; de Souza, E.P.; Nogueira, V.H.R.; de Godoy, M.O.; Nakamura, A.M.; et al. Non-Toxic Dimeric Peptides Derived from the Bothropstoxin-I Are Potent SARS-CoV-2 and Papain-like Protease Inhibitors. Molecules 2021, 26, 4896. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Reid, P.F.; Qin, Z.-H. Cobrotoxin Could Be an Effective Therapeutic for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1258–1260. [Google Scholar] [CrossRef]
- Kasozi, K.I.; Niedbała, G.; Alqarni, M.; Zirintunda, G.; Ssempijja, F.; Musinguzi, S.P.; Usman, I.M.; Matama, K.; Hetta, H.F.; Mbiydzenyuy, N.E.; et al. Bee Venom—A Potential Complementary Medicine Candidate for SARS-CoV-2 Infections. Front. Public Health 2020, 8, 594458. [Google Scholar] [CrossRef]
- Block, J. High Risk COVID-19: Potential Intervention at Multiple Points in the COVID-19 Disease Process Via Prophylactic Treatment with Azithromycin or Bee Derived Products. Preprints 2020, 2020040013. [Google Scholar] [CrossRef]
- Enayathullah, M.G.; Parekh, Y.; Banu, S.; Ram, S.; Nagaraj, R.; Kumar, B.K.; Idris, M.M. Gramicidin S and Melittin: Potential Anti-Viral Therapeutic Peptides to Treat SARS-CoV-2 Infection. Sci. Rep. 2022, 12, 3446. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Cerni, F.A.; Oliveira, I.S.; Jenkins, T.P.; Argemí, L.; Sørensen, C.V.; Ahmadi, S.; Barbosa, J.E.; Laustsen, A.H. Bee Updated: Current Knowledge on Bee Venom and Bee Envenoming Therapy. Front. Immunol. 2019, 10, 2090. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, R. In Silico Prediction of Anti–SARS-CoV-2 Effect of Dermaseptin Peptides from Amphibian Origin. Trends Pept. Protein Sci. 2020, 5, 1–9 (e6). [Google Scholar] [CrossRef]
- Mahnam, K.; Lotfi, M.; Shapoorabadi, F.A. Examining the Interactions Scorpion Venom Peptides (HP1090, Meucin-13, and Meucin-18) with the Receptor Binding Domain of the Coronavirus Spike Protein to Design a Mutated Therapeutic Peptide. J. Mol. Graph. Model. 2021, 107, 107952. [Google Scholar] [CrossRef]
- Alqathama, A.A.; Ahmad, R.; Alsaedi, R.B.; Alghamdi, R.A.; Abkar, E.H.; Alrehaly, R.H.; Abdalla, A.N. The Vital Role of Animal, Marine, and Microbial Natural Products against COVID-19. Pharm. Biol. 2022, 60, 509–524. [Google Scholar] [CrossRef]
- Chérifi, F.; Laraba-Djebari, F. Bioactive Molecules Derived from Snake Venoms with Therapeutic Potential for the Treatment of Thrombo-Cardiovascular Disorders Associated with COVID-19. Protein J. 2021, 40, 799–841. [Google Scholar] [CrossRef]
- Gouda, A.S.; Mégarbane, B. Snake Venom-derived Bradykinin-potentiating Peptides: A Promising Therapy for COVID-19? Drug Dev. Res. 2020, 82, 38–48. [Google Scholar] [CrossRef]
- Kalita, B.; Saviola, A.J.; Samuel, S.P.; Mukherjee, A.K. State-of-the-Art Review—A Review on Snake Venom-Derived Antithrombotics: Potential Therapeutics for COVID-19-Associated Thrombosis? Int. J. Biol. Macromol. 2021, 192, 1040–1057. [Google Scholar] [CrossRef]
- Kini, R.M. Anticoagulant Proteins from Snake Venoms: Structure, Function and Mechanism. Biochem. J. 2006, 397, 377–387. [Google Scholar] [CrossRef]
- Silva, M.C.; Lopes Silva, T.; Silva, M.V.; Mota, C.M.; Santiago, F.M.; Fonseca, K.C.; Oliveira, F.; Mineo, T.W.P.; Mineo, J.R. Interaction between TNF and BmooMP-Alpha-I, a Zinc Metalloprotease Derived from Bothrops moojeni Snake Venom, Promotes Direct Proteolysis of This Cytokine: Molecular Modeling and Docking at a Glance. Toxins 2016, 8, 223. [Google Scholar] [CrossRef]
- Hu, Y.; Meng, B.; Yin, S.; Yang, M.; Li, Y.; Liu, N.; Li, S.; Liu, Y.; Sun, D.; Wang, S.; et al. Scorpion Venom Peptide HsTx2 Suppressed PTZ-Induced Seizures in Mice via the Circ_0001293/MiR-8114/TGF-Β2 Axis. J. Neuroinflamm. 2022, 19, 284. [Google Scholar] [CrossRef]
- Tao, J.; Yin, S.; Song, Y.; Zeng, L.; Li, S.; Liu, N.; Sun, H.; Fu, Z.; Wang, Y.; Li, Y.; et al. Novel Scorpion Venom Peptide HsTx2 Ameliorates Cerebral Ischemic Brain Injury in Rats via the MAPK Signaling Pathway. Biochem. Biophys. Res. Commun. 2021, 534, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Kocyigit, A.; Guler, E.M.; Kaleli, S. Anti-Inflammatory and Antioxidative Properties of Honey Bee Venom on Freund’s Complete Adjuvant-Induced Arthritis Model in Rats. Toxicon 2019, 161, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Yao, L.; Zhang, B.; Zhang, S.; Guo, J. Anti-Inflammatory Effects of Neurotoxin-Nna, a Peptide Separated from the Venom of Naja Naja Atra. BMC Complement. Altern. Med. 2013, 13, 86. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-Z.; He, H.; Han, R.; Zhu, J.-L.; Kou, J.-Q.; Ding, X.-L.; Qin, Z.-H. The Protective Effects of Cobra Venom from Naja Naja Atra on Acute and Chronic Nephropathy. Evid.-Based Complement. Altern. Med. 2013, 2013, e478049. [Google Scholar] [CrossRef]
- Zhu, J.; Cui, K.; Kou, J.; Wang, S.; Xu, Y.; Ding, Z.; Han, R.; Qin, Z. Naja Naja Atra Venom Protects against Manifestations of Systemic Lupus Erythematosus in MRL/Lpr Mice. Evid.-Based Complement. Altern. Med. 2014, 2014, e969482. [Google Scholar] [CrossRef]
- Avalo, Z.; Barrera, M.C.; Agudelo-Delgado, M.; Tobón, G.J.; Cañas, C.A. Biological Effects of Animal Venoms on the Human Immune System. Toxins 2022, 14, 344. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, I.; Ferreira, I.; Jacob, B.; Cardenas, K.; Cerni, F.; Baia-da-Silva, D.; Arantes, E.; Monteiro, W.; Pucca, M. Harnessing the Power of Venomous Animal-Derived Toxins against COVID-19. Toxins 2023, 15, 159. https://doi.org/10.3390/toxins15020159
Oliveira I, Ferreira I, Jacob B, Cardenas K, Cerni F, Baia-da-Silva D, Arantes E, Monteiro W, Pucca M. Harnessing the Power of Venomous Animal-Derived Toxins against COVID-19. Toxins. 2023; 15(2):159. https://doi.org/10.3390/toxins15020159
Chicago/Turabian StyleOliveira, Isadora, Isabela Ferreira, Beatriz Jacob, Kiara Cardenas, Felipe Cerni, Djane Baia-da-Silva, Eliane Arantes, Wuelton Monteiro, and Manuela Pucca. 2023. "Harnessing the Power of Venomous Animal-Derived Toxins against COVID-19" Toxins 15, no. 2: 159. https://doi.org/10.3390/toxins15020159
APA StyleOliveira, I., Ferreira, I., Jacob, B., Cardenas, K., Cerni, F., Baia-da-Silva, D., Arantes, E., Monteiro, W., & Pucca, M. (2023). Harnessing the Power of Venomous Animal-Derived Toxins against COVID-19. Toxins, 15(2), 159. https://doi.org/10.3390/toxins15020159