A Pore Forming Toxin-like Protein Derived from Chinese Red Belly Toad Bombina maxima Triggers the Pyroptosis of Hippomal Neural Cells and Impairs the Cognitive Ability of Mice

Abstract
:1. Introduction
2. Results
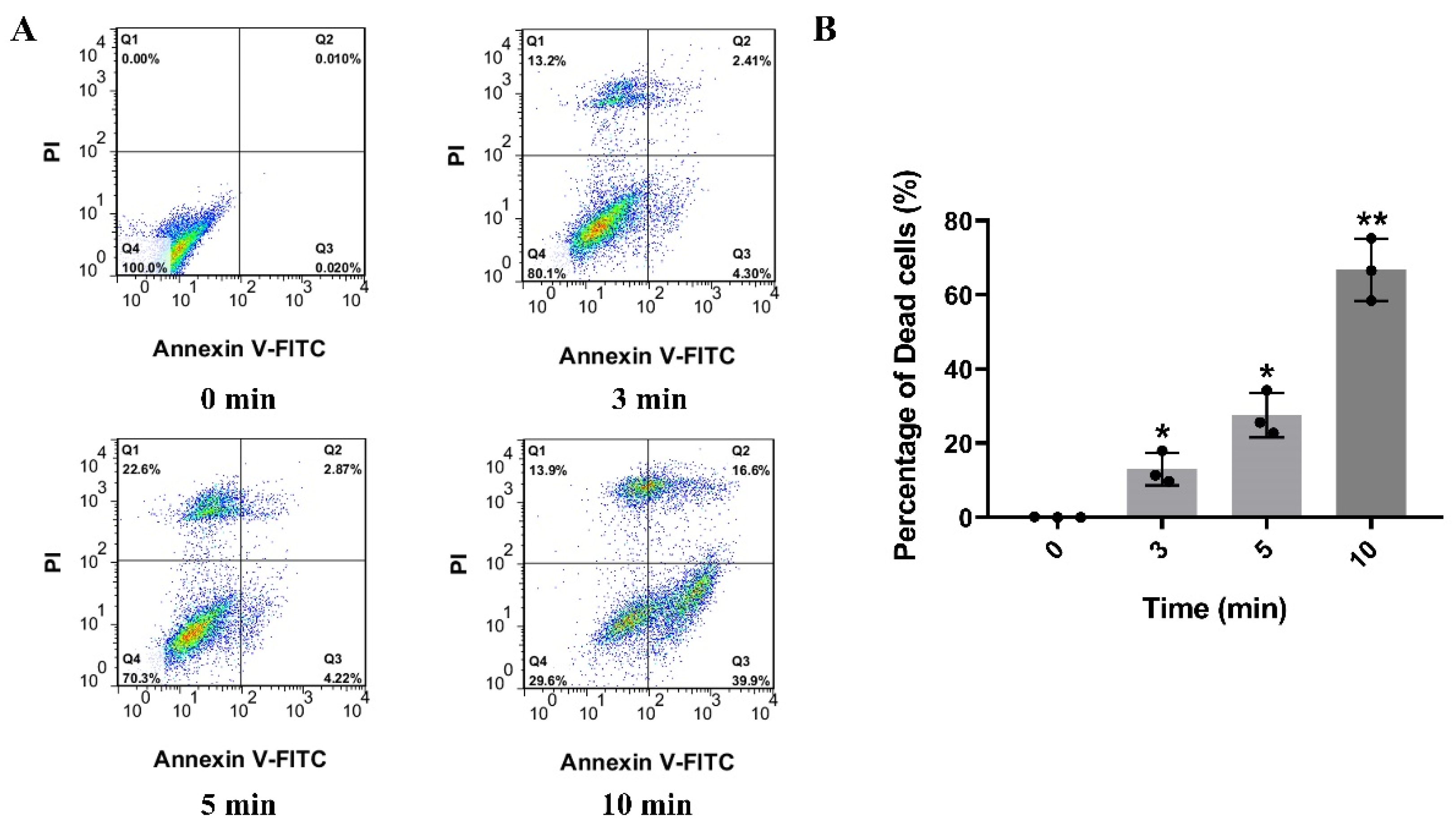
2.1. βγ-CAT Displays Strong Cytotoxicity and Induces Death of Mouse Hippocapmal Neuronal Cells
2.2. βγ-CAT Triggers Gasdermin-E-Dependent Pyroptosis of HT-22 Cells
2.3. The Pyroptosis of HT-22 Cells Triggered by βγ-CAT Depend on Its Membrane Binding, Oligomerization and Endocytosis
2.4. βγ-CAT Crosses the Blood-Brain Barrier and Reduces Cognitive Function of Mice
3. Discussion
4. Conclusions
- (1)
- βγ-CAT shows strong cytotoxicity to HT-22 hippocampal neuronal cells and triggers the GSDME-dependent pyroptosis of HT-22 cells.
- (2)
- The pyroptosis of HT-22 cells induced by βγ-CAT relies on the membrane binding, oligomerization, and endocytosis of βγ-CAT.
- (3)
- βγ-CAT can cross the BBB and affects the cognitive function of mice.
5. Materials and Methods
5.1. Animals
5.2. Cell Lines, Antibodies and Reagents
5.3. LDH Release Detection
5.4. MTT Assay
5.5. Flow Cytometry Detection of HT-22 Cell Death
5.6. PCR and RT-qPCR Assay
5.7. Western Blotting Assay
5.8. Enzyme-Linked Immuno Sorbent Assay (ELISA)
5.9. Flow Cytometry Detection of the Membrane Binding of βγ-CAT
5.10. Confocal Microscopy Assay of the Endocytosis and Co-Localization of βγ-CAT
5.11. In Vitro BBB Model Establishment and βγ-CAT Penetration Detection
5.12. In Vivo BBB Penetration Assay
5.13. Water Maze Assay for Assessment of Spatial Learning and Memory of Mice
5.14. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dal Peraro, M.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Cirauqui, N.; Abriata, L.A.-O.; van der Goot, F.A.-O.X.; Dal Peraro, M. Structural, physicochemical and dynamic features conserved within the aerolysin pore-forming toxin family. Sci. Rep. 2017, 7, 13932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczesny, P.; Iacovache, I.; Muszewska, A.; Ginalski, K.; van der Goot, F.G.; Grynberg, M. Extending the aerolysin family: From bacteria to vertebrates. PLoS ONE 2011, 6, e20349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cressiot, B.A.-O.; Ouldali, H.; Pastoriza-Gallego, M.; Bacri, L.A.-O.; Van der Goot, F.G.; Pelta, J. Aerolysin, a Powerful Protein Sensor for Fundamental Studies and Development of Upcoming Applications. ACS Sens. 2019, 4, 530–548. [Google Scholar] [CrossRef]
- Zhang, Y. Why do we study animal toxins? Zool. Res. 2015, 36, 183–222. [Google Scholar]
- Seni-Silva, A.A.-O.; Maleski, A.A.-O.; Souza, M.A.-O.; Falcao, M.A.-O.; Disner, G.A.-O.; Lopes-Ferreira, M.A.-O.; Lima, C.A.-O.X. Natterin-like depletion by CRISPR/Cas9 impairs zebrafish (Danio rerio) embryonic development. BMC Genom. 2022, 23, 123. [Google Scholar] [CrossRef]
- Lima, C.A.-O.X.; Disner, G.A.-O.; Falcão, M.A.P.; Seni-Silva, A.C.; Maleski, A.A.-O.; Souza, M.M.; Reis Tonello, M.C.; Lopes-Ferreira, M. The Natterin Proteins Diversity: A Review on Phylogeny, Structure, and Immune Function. Toxins 2021, 13, 538. [Google Scholar] [CrossRef]
- Chen, L.L.; Xie, J.; Cao, D.D.; Jia, N.; Li, Y.J.; Sun, H.; Li, W.F.; Hu, B.; Chen, Y.; Zhou, C.Z. The pore-forming protein Aep1 is an innate immune molecule that prevents zebrafish from bacterial infection. Dev. Comp. Immunol. 2018, 82, 49–54. [Google Scholar] [CrossRef]
- Pang, Y.; Gou, M.; Yang, K.; Lu, J.; Han, Y.; Teng, H.; Li, C.; Wang, H.; Liu, C.; Zhang, K.; et al. Crystal structure of a cytocidal protein from lamprey and its mechanism of action in the selective killing of cancer cells. Cell Commun. Signal. 2019, 17, 54. [Google Scholar] [CrossRef] [Green Version]
- Lisman, J.; Buzsáki, G.; Eichenbaum, H.; Nadel, L.; Ranganath, C.; Redish, A.D. Viewpoints: How the hippocampus contributes to memory, navigation and cognition. Nat. Neurosci. 2017, 20, 1434–1447. [Google Scholar] [CrossRef]
- Ekstrom, A.D.; Ranganath, C.A.-O. Space, time, and episodic memory: The hippocampus is all over the cognitive map. Hippocampus 2018, 28, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.J.; Chiu, I.M. Bacterial Signaling to the Nervous System through Toxins and Metabolites. J. Mol. Biol. 2017, 429, 587–605. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Lai, R. The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef] [PubMed]
- Pirazzini, M.; Montecucco, C.A.-O.; Rossetto, O. Toxicology and pharmacology of botulinum and tetanus neurotoxins: An update. Arch. Toxicol. 2022, 96, 1521–1539. [Google Scholar] [CrossRef]
- Wioland, L.; Dupont, J.; Bossu, J.-L.; Popoff, M.R.; Poulain, B. Attack of the nervous system by Clostridium perfringens Epsilon toxin: From disease to mode of action on neural cells. Toxicon 2013, 75, 122–135. [Google Scholar] [CrossRef]
- Liu, S.B.; He, Y.Y.; Zhang, Y.; Lee, W.H.; Qian, J.Q.; Lai, R.; Jin, Y. A novel non-lens betagamma-crystallin and trefoil factor complex from amphibian skin and its functional implications. PLoS ONE 2008, 3, e1770. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Xiang, Y.; Zeng, L.; Ma, X.; Lee, W.; Zhang, Y. Characterization of the bg-crystallin domains of bg-CAT, a non-lens bg-crystallin and trefoil factor complex, from the skin of the toad Bombina maxima. Biochimie 2011, 93, 1865–1872. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Yan, C.; Guo, X.L.; Zhou, K.F.; Li, S.A.; Gao, Q.; Wang, X.; Zhao, F.; Liu, J.; Lee, W.H. Host-derived, pore-forming toxin–like protein and trefoil factor complex protects the host against microbial infection. Proc. Natl. Acad. Sci. USA 2014, 111, 6702–6707. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Deng, C.J.; Duan, Y.L.; Ye, C.J.; Gong, D.H.; Guo, X.L.; Lee, W.; Zhou, J.; Li, S.A.; Zhang, Y. An Aerolysin-like Pore-Forming Protein Complex Targets Viral Envelope to Inactivate Herpes Simplex Virus Type 1. J. Immunol. 2021, 207, 888–901. [Google Scholar] [CrossRef]
- Guo, X.L.; Liu, L.Z.; Wang, Q.Q.; Liang, J.Y.; Lee, W.H.; Xiang, Y.; Li, S.A.; Zhang, Y. Endogenous pore-forming protein complex targets acidic glycosphingolipids in lipid rafts to initiate endolysosome regulation. Commun. Biol. 2019, 2, 59. [Google Scholar] [CrossRef]
- Wang, Q.; Bian, X.; Zeng, L.; Pan, F.; Liu, L.; Liang, J.; Wang, L.; Zhou, K.; Lee, W.; Xiang, Y.; et al. A cellular endolysosome-modulating pore-forming protein from a toad is negatively regulated by its paralog under oxidizing conditions. J. Biol. Chem. 2020, 295, 10293–10306. [Google Scholar] [CrossRef] [PubMed]
- Schnaar, R.L. Gangliosides of the Vertebrate Nervous System. J. Mol. Biol. 2016, 428, 3325–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, S.; Cantuti Castelvetri, L.; Simons, M. Metabolism and functions of lipids in myelin. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2015, 1851, 999–1005. [Google Scholar] [CrossRef]
- Bonetto, G.A.-O.; Di Scala, C.A.-O. Importance of Lipids for Nervous System Integrity: Cooperation between Gangliosides and Sulfatides in Myelin Stability. J. Neurosci. 2019, 39, 6218–6220. [Google Scholar] [CrossRef] [PubMed]
- Bertheloot, D.; Latz, E.A.-O.; Franklin, B.A.-O. Necroptosis, pyroptosis and apoptosis: An intricate game of cell death. Cell Mol. Immunol. 2021, 18, 1106–1121. [Google Scholar] [CrossRef]
- Li, J.; Cao, F.; Yin, H.-l.; Huang, Z.; Lin, Z.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Gao, W.; Shao, F. Pyroptosis: Gasdermin-Mediated Programmed Necrotic Cell Death. Trends Biochem. Sci. 2017, 42, 245–254. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Q.Q.; Zhao, Z.; Deng, C.J. Animal secretory endolysosome channel discovery. Zool. Res. 2021, 42, 141–152. [Google Scholar] [CrossRef]
- Opitz, B. Memory Function and the Hippocampus. Front. Neurol. Neurosci. 2014, 34, 51–59. [Google Scholar]
- Sweatt, J.D. Hippocampal function in cognition. Psychopharmacology 2004, 174, 99–110. [Google Scholar] [CrossRef]
- Brun, O.; Zoukimian, C.; Oliveira-Mendes, B.; Montnach, J.; Lauzier, B.; Ronjat, M.; Béroud, R.; Lesage, F.; Boturyn, D.; De Waard, M. Chemical Synthesis of a Functional Fluorescent-Tagged α-Bungarotoxin. Toxins 2022, 14, 79. [Google Scholar] [CrossRef] [PubMed]
- Haigh, J.L.; Williamson, D.J.; Poole, E.; Guo, Y.; Zhou, D.J.; Webb, M.E.; Deuchars, S.A.; Deuchars, J.; Turnbull, W.B. A versatile cholera toxin conjugate for neuronal targeting and tracing. Chem. Commun. 2020, 56, 6098–6101. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.G.; de Ávila, R.A.M.; Chávez-Olórtegui, C.D.; Lobato, F.C.F. Clostridium perfringens epsilon toxin: The third most potent bacterial toxin known. Anaerobe 2014, 30, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Linden, J.R.; Ma, Y.; Zhao, B.; Harris, J.M.; Rumah, K.R.; Schaeren-Wiemers, N.; Vartanian, T. Clostridium perfringens Epsilon Toxin Causes Selective Death of Mature Oligodendrocytes and Central Nervous System Demyelination. mBio 2015, 6, e02513. [Google Scholar] [CrossRef] [Green Version]
- Obermeier, B.A.-O.X.; Daneman, R.; Ransohoff, R.M. Development, maintenance and disruption of the blood-brain barrier. Nat. Med. 2013, 19, 1584–1596. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Gan, L.; Ren, L.; Lin, Y.; Ma, C.; Lin, X. Factors influencing the blood-brain barrier permeability. Brain Res. 2022, 1788, 147937. [Google Scholar] [CrossRef]
- Adler, D.; Linden, J.R.; Shetty, S.V.; Ma, Y.; Bokori-Brown, M.; Titball, R.W.; Vartanian, T. Clostridium perfringens Epsilon Toxin Compromises the Blood-Brain Barrier in a Humanized Zebrafish Model. iScience 2019, 15, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Schnaar, R.L. Brain gangliosides in axon–myelin stability and axon regeneration. FEBS Lett. 2010, 584, 1741–1747. [Google Scholar] [CrossRef] [Green Version]
- Profaci, C.P.; Munji, R.N.; Pulido, R.S.; Daneman, R. The blood-brain barrier in health and disease: Important unanswered questions. J. Exp. Med. 2020, 217, e20190062. [Google Scholar] [CrossRef] [Green Version]
- Mayo, L.; Trauger, S.A.; Blain, M.; Nadeau, M.; Patel, B.; Alvarez, J.I.; Mascanfroni, I.D.; Yeste, A.; Kivisäkk, P.; Kallas, K.; et al. Regulation of astrocyte activation by glycolipids drives chronic CNS inflammation. Nat. Med. 2014, 20, 1147–1156. [Google Scholar] [CrossRef] [Green Version]
- Goshi, N.; Morgan, R.K.; Lein, P.J.; Seker, E. A primary neural cell culture model to study neuron, astrocyte, and microglia interactions in neuroinflammation. J. Neuroinflamm. 2020, 17, 155. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.A. Targeting Central Nervous System Regeneration with Cell Type Specificity. Neurosurg. Clin. N. Am. 2021, 32, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu. Rev. Immunol. 2017, 35, 441–468. [Google Scholar] [CrossRef]
- McNamara, N.B.; Munro, D.A.D.; Bestard-Cuche, N.; Uyeda, A.; Bogie, J.F.J.; Hoffmann, A.; Holloway, R.K.; Molina-Gonzalez, I.; Askew, K.E.; Mitchell, S.; et al. Microglia regulate central nervous system myelin growth and integrity. Nature 2023, 613, 120–129. [Google Scholar] [CrossRef]
- Liu, D.; Dong, Z.; Xiang, F.; Liu, H.; Wang, Y.; Wang, Q.; Rao, J. Dendrobium Alkaloids Promote Neural Function After Cerebral Ischemia-Reperfusion Injury Through Inhibiting Pyroptosis Induced Neuronal Death in both In Vivo and In Vitro Models. Neurochem. Res. 2020, 45, 437–454. [Google Scholar] [CrossRef]
- Liu, Y.; Wen, D.; Gao, J.; Xie, B.; Yu, H.; Shen, Q.; Zhang, J.; Jing, W.; Cong, B.; Ma, C. Methamphetamine induces GSDME-dependent cell death in hippocampal neuronal cells through the endoplasmic reticulum stress pathway. Brain Res. Bull. 2020, 162, 73–83. [Google Scholar] [CrossRef]
- Aguayo, S.; Schuh, C.M.A.P.; Vicente, B.; Aguayo, L.G. Association between Alzheimer’s Disease and Oral and Gut Microbiota: Are Pore Forming Proteins the Missing Link? J. Alzheimer’s Dis. 2018, 65, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Di Scala, C.; Armstrong, N.; Chahinian, H.; Chabrière, E.; Fantini, J.; Yahi, N. AmyP53, a Therapeutic Peptide Candidate for the Treatment of Alzheimer’s and Parkinson’s Disease: Safety, Stability, Pharmacokinetics Parameters and Nose-to Brain Delivery. Int. J. Mol. Sci. 2022, 23, 13383. [Google Scholar] [CrossRef]
- Amodeo, G.F.; Pavlov, E.V. Amyloid β, α-synuclein and the c subunit of the ATP synthase: Can these peptides reveal an amyloidogenic pathway of the permeability transition pore? Biochim. Biophys. Acta (BBA) Biomembr. 2021, 1863, 183531. [Google Scholar] [CrossRef]
- Amodeo, G.F.; Lee, B.Y.; Krilyuk, N.; Filice, C.T.; Valyuk, D.; Otzen, D.E.; Noskov, S.; Leonenko, Z.; Pavlov, E.V. C subunit of the ATP synthase is an amyloidogenic calcium dependent channel-forming peptide with possible implications in mitochondrial permeability transition. Sci. Rep. 2021, 11, 8744. [Google Scholar] [CrossRef]
- Venko, K.; Novič, M.; Stoka, V.; Žerovnik, E. Prediction of Transmembrane Regions, Cholesterol, and Ganglioside Binding Sites in Amyloid-Forming Proteins Indicate Potential for Amyloid Pore Formation. Front. Mol. Neurosci. 2021, 14, 619496. [Google Scholar] [CrossRef]
- Sepehri, A.; Lazaridis, T. Putative Structures of Membrane-Embedded Amyloid β Oligomers. ACS Chem. Neurosci. 2023, 14, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Pastore, A.; Raimondi, F.; Rajendran, L.; Temussi, P.A. Why does the Aβ peptide of Alzheimer share structural similarity with antimicrobial peptides? Commun. Biol. 2020, 3, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatulian, S.A. Challenges and hopes for Alzheimer’s disease. Drug Discov. Today 2022, 27, 1027–1043. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.W. The chemistry of poisons in amphibian skin. Proc. Natl. Acad. Sci. USA 1995, 92, 9–13. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, L.A.; Course, L.S.; O’Connell, J.D.; Paulo, J.A.; Trauger, S.A.; Gygi, S.P.; Murray, A.W. Rapid toxin sequestration modifies poison frog physiology. J. Exp. Biol. 2021, 224, jeb230342. [Google Scholar] [CrossRef]
- Crowley, L.C.; Marfell, B.J.; Scott, A.P.; Waterhouse, N.J. Quantitation of Apoptosis and Necrosis by Annexin V Binding, Propidium Iodide Uptake, and Flow Cytometry. Cold Spring Harb. Protoc. 2016, 11, pdb.prot087288. [Google Scholar] [CrossRef]
- Ye, Q.Q.; Tian, H.F.; Chen, B.; Shao, J.R.; Qin, Y.; Wen, J.F. Giardia’s primitive GPL biosynthesis pathways with parasitic adaptation ‘patches’: Implications for Giardia’s evolutionary history and for finding targets against Giardiasis. Sci. Rep. 2017, 7, 9507. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, A.; Matsuoka, S. Spatial and Functional Crosstalk between the Mitochondrial Na+-Ca2+ Exchanger NCLX and the Sarcoplasmic Reticulum Ca2+ Pump SERCA in Cardiomyocytes. Int. J. Mol. Sci. 2022, 23, 7948. [Google Scholar] [CrossRef]
- Liang, W.; Xu, W.; Zhu, J.; Zhu, Y.; Gu, Q.; Li, Y.; Guo, C.; Huang, Y.; Yu, J.; Wang, W.; et al. Ginkgo biloba extract improves brain uptake of ginsenosides by increasing blood-brain barrier permeability via activating A1 adenosine receptor signaling pathway. J. Ethnopharmacol. 2020, 246, 112243. [Google Scholar] [CrossRef]
- Czupalla, C.J.; Liebner, S.; Devraj, K. In vitro models of the blood-brain barrier. Methods Mol. Biol 2014, 1135, 415–437. [Google Scholar] [PubMed]
- Devraj, K.; Guérit, S.; Macas, J.; Reiss, Y. An In Vivo Blood-brain Barrier Permeability Assay in Mice Using Fluorescently Labeled Tracers. J. Vis. Exp. 2018, 132, e57038. [Google Scholar]
- Sun, W.; Yang, Y.; Wu, Z.; Chen, X.; Li, W.; An, L. Chronic Cyanuric Acid Exposure Depresses Hippocampal LTP but Does Not Disrupt Spatial Learning or Memory in the Morris Water Maze. Neurotox. Res. 2021, 39, 1148–1159. [Google Scholar] [CrossRef]
- Dinel, A.L.; Lucas, C.; Guillemet, D.; Laye, S.; Pallet, V.; Joffre, C. Chronic Supplementation with a Mix of Salvia officinalis and Salvia lavandulaefolia Improves Morris Water Maze Learning in Normal Adult C57Bl/6J Mice. Nutrients 2020, 12, 1777. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Product Length |
|---|---|---|
| PCR | ||
| GSDMA-Forward | TCCCTCCTGGAGAAAAGCCA | 338 bp |
| GSDMA-Reverse | ACTTAGCACTGTCAGAGCCC | |
| GSDMB-Forward | TGGATGCCGGCACTACACAAC | 248 bp |
| GSDMB-Reverse | GGTAGTTCCCTCTTCAGCTTCC | |
| GSDMC-Forward | GATCTGAGGCCTGTCAAATGC | 524 bp |
| GSDMC-Reverse | TCTGTTTGCCACTGTCCACT | |
| GSDMD-Forward | CCGGAGTGTTTTGGCTCCTT | 260 bp |
| GSDMD-Reverse | ACCACAAACAGGTCATCCCC | |
| GSDME-Forward | GTCAGCGCACTAGCAGAAATG | 173 bp |
| GSDME-Reverse | ATGCCAAACCTCTCTGTGTC | |
| PJVK-Forward | GCTGACAAGTACCAACCCCT | 595 bp |
| PJVK-Reverse | CACAAATGTCGAAGGCACCG | |
| β-actin-Forward | CCACCATGTACCCAGGCATT | 253 bp |
| β-actin-Reverse | AGGGTGTAAAACGCAGCTCA | |
| RT-qPCR | ||
| GSDMA-Forward | TCCCTCCTGGAGAAAAGCCA | 261 bp |
| GSDMA-Reverse | GTGCTTCCAGGGTCACTTCG | |
| GSDMB-Forward | CCGTTAGAAGCCTTGTTGATGC | 180 bp |
| GSDMB-Reverse | CCGTTGAGTCTACATTATCCAG | |
| GSDMC-Forward | CAGATGCAACCAAATTCTGCC | 207 bp |
| GSDMC-Reverse | TGGTTTCGACATCCAGGTCA | |
| GSDMD-Forward | GATCAAGGAGGTAAGCGGCA | 195 bp |
| GSDMD-Reverse | CACTCCGGTTCTGGTTCTGG | |
| GSDME-Forward | AGTTTTCCTGGGGACTTGCT | 170 bp |
| GSDME-Reverse | CAATGTCAGCAGAGGCAAACAA | |
| PJVK-Forward | TCAGCGAAGCTGACAAGTACC | 300 bp |
| PJVK-Reverse | CCACCTCATGTTTGGTCACG | |
| β-actin-Forward | CCACCATGTACCCAGGCATT | 253 bp |
| β-actin-Reverse | AGGGTGTAAAACGCAGCTCA | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Q.; Wang, Q.; Lee, W.; Xiang, Y.; Yuan, J.; Zhang, Y.; Guo, X. A Pore Forming Toxin-like Protein Derived from Chinese Red Belly Toad Bombina maxima Triggers the Pyroptosis of Hippomal Neural Cells and Impairs the Cognitive Ability of Mice. Toxins 2023, 15, 191. https://doi.org/10.3390/toxins15030191
Ye Q, Wang Q, Lee W, Xiang Y, Yuan J, Zhang Y, Guo X. A Pore Forming Toxin-like Protein Derived from Chinese Red Belly Toad Bombina maxima Triggers the Pyroptosis of Hippomal Neural Cells and Impairs the Cognitive Ability of Mice. Toxins. 2023; 15(3):191. https://doi.org/10.3390/toxins15030191
Chicago/Turabian StyleYe, Qingqing, Qiquan Wang, Wenhui Lee, Yang Xiang, Jixue Yuan, Yun Zhang, and Xiaolong Guo. 2023. "A Pore Forming Toxin-like Protein Derived from Chinese Red Belly Toad Bombina maxima Triggers the Pyroptosis of Hippomal Neural Cells and Impairs the Cognitive Ability of Mice" Toxins 15, no. 3: 191. https://doi.org/10.3390/toxins15030191