Mapping the Immune Cell Microenvironment with Spatial Profiling in Muscle Tissue Injected with the Venom of Daboia russelii

 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
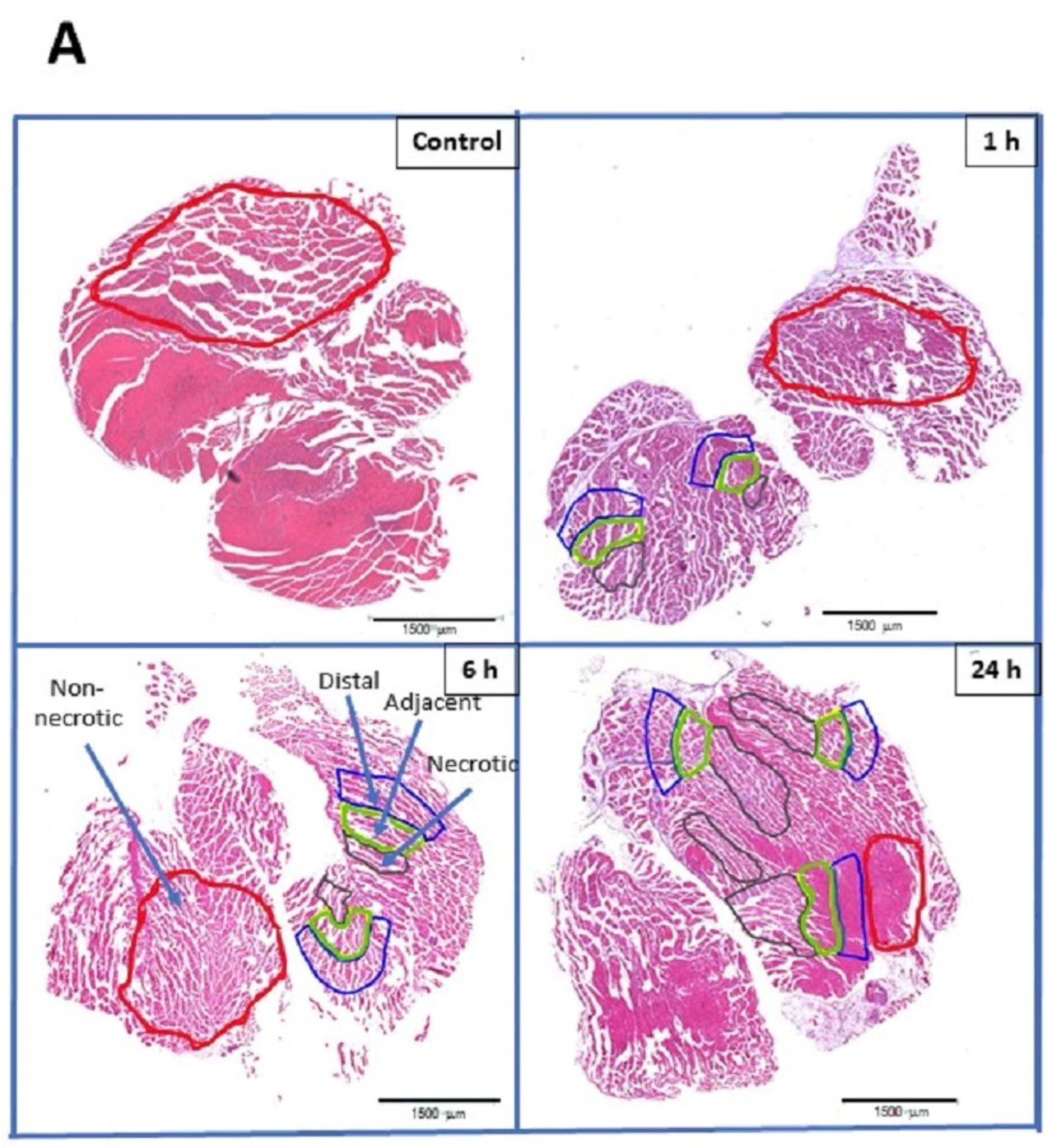
2.1. The Number of Inflammatory Cells Varies Depending on the Severity of Tissue Damage
2.2. GeoMx® DSP Allowed the Quantification of Markers of Different Inflammatory Cells
3. Discussion
4. Materials and Methods
4.1. Venom
4.2. Experimental Model
4.3. Histology and Immunohistochemistry
4.4. GeoMx® Digital Spatial Profiler
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gutiérrez, J.M. Snakebite Envenomation as a Neglected Tropical Disease: New Impetus for Confronting an Old Scourge. In Handbook of Venoms and Toxins of Reptiles; CRC Press: Boca Raton, FL, USA, 2021; pp. 471–484. [Google Scholar] [CrossRef]
- Warrell, D.A. Snake Bite. Lancet 2010, 375, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite Envenoming. Nat. Rev. Dis. Prim. 2017, 3, 17063. [Google Scholar] [CrossRef] [PubMed]
- Ownby, C.L.; Fletcher, J.E.; Colberg, T.R. Cardiotoxin 1 from Cobra (Naja naja atra) Venom Causes Necrosis of Skeletal Muscle in vivo. Toxicon 1993, 31, 697–709. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Ownby, C.L. Skeletal Muscle Degeneration Induced by Venom Phospholipases A 2: Insights into the Mechanisms of Local and Systemic Myotoxicity. Toxicon 2003, 42, 915–931. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Rucavado, A. Snake Venom Metalloproteinases: Their Role in the Pathogenesis of Local Tissue Damage. Biochimie 2000, 82, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.; Moreira, V.; Gutiérrez, J.M. Venoms. In Inflammation—From Molecular and Cellular Mechanisms to the Clinic; Wiley: Hoboken, NJ, USA, 2017; pp. 99–128. [Google Scholar] [CrossRef]
- Moreira, V.; Teixeira, C.; Borges Da Silva, H.; D’Império Lima, M.R.; Dos-Santos, M.C. The Role of TLR2 in the Acute Inflammatory Response Induced by Bothrops atrox Snake Venom. Toxicon 2016, 118, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Rucavado, A.; Nicolau, C.A.; Escalante, T.; Kim, J.; Herrera, C.; Gutiérrez, J.M.; Fox, J.W. Viperid Envenomation Wound Exudate Contributes to Increased Vascular Permeability via a DAMPs/TLR-4 Mediated Pathway. Toxins 2016, 8, 349. [Google Scholar] [CrossRef] [Green Version]
- Fontana, B.C.; Soares, A.M.; Zuliani, J.P.; Gonçalves, G.M. Role of Toll-like Receptors in Local Effects in a Model of Experimental Envenoming Induced by Bothrops jararacussu Snake Venom and by Two Phospholipases A2. Toxicon 2022, 214, 145–154. [Google Scholar] [CrossRef]
- Moreira, V.; Dos-Santos, M.C.; Nascimento, N.G.; da Silva, H.B.; Fernandes, C.M.; D’Império Lima, M.R.; Teixeira, C. Local Inflammatory Events Induced by Bothrops atrox Snake Venom and the Release of Distinct Classes of Inflammatory Mediators. Toxicon 2012, 60, 12–20. [Google Scholar] [CrossRef]
- Rucavado, A.; Escalante, T.; Kalogeropoulos, K.; Camacho, E.; Gutiérrez, J.M.; Fox, J.W. Analysis of Wound Exudates Reveals Differences in the Patterns of Tissue Damage and Inflammation Induced by the Venoms of Daboia russelii and Bothrops asper in Mice. Toxicon 2020, 186, 94–104. [Google Scholar] [CrossRef]
- Rucavado, A.; Escalante, T.; Teixeira, C.F.P.; Fernándes, C.M.; Díaz, C.; Gutiérrez, J.M. Increments in Cytokines and Matrix Metalloproteinases in Skeletal Muscle after Injection of Tissue-Damaging Toxins from the Venom of the Snake Bothrops asper. Mediat. Inflamm. 2002, 11, 121–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petricevich, V.L.; Teixeira, C.F.P.; Tambourgi, D.V.; Gutiérrez, J.M. Increments in Serum Cytokine and Nitric Oxide Levels in Mice Injected with Bothrops asper and Bothrops jararaca Snake Venoms. Toxicon 2000, 38, 1253–1266. [Google Scholar] [CrossRef]
- Alsolaiss, J.; Evans, C.A.; Oluoch, G.O.; Casewell, N.R.; Harrison, R.A. Profiling the Murine Acute Phase and Inflammatory Responses to African Snake Venom: An Approach to Inform Acute Snakebite Pathology. Toxins 2022, 14, 229. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, N.; Escalante, T.; Gutiérrez, J.M.; Rucavado, A. Skin Pathology Induced by Snake Venom Metalloproteinase: Acute Damage, Revascularization, and Re-Epithelization in a Mouse Ear Model. J. Invest. Dermatol. 2008, 128, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Hernández, R.; Cabalceta, C.; Saravia-Otten, P.; Chaves, A.; Gutiérrez, J.M.; Rucavado, A. Poor Regenerative Outcome after Skeletal Muscle Necrosis Induced by Bothrops asper Venom: Alterations in Microvasculature and Nerves. PLoS ONE 2011, 6, e19834. [Google Scholar] [CrossRef]
- Ranéia e Silva, P.A.; da Costa Neves, A.; da Rocha, C.B.; Moura-da-Silva, A.M.; Faquim-Mauro, E.L. Differential Macrophage Subsets in Muscle Damage Induced by a K49-PLA2 from Bothrops jararacussu Venom Modulate the Time Course of the Regeneration Process. Inflammation 2019, 42, 1542–1554. [Google Scholar] [CrossRef] [PubMed]
- Zuliani, J.P.; Gutiérrez, J.M.; Silva, L.L.C.E.; Sampaio, S.C.; Lomonte, B.; Pereira Teixeira, C.D.F. Activation of Cellular Functions in Macrophages by Venom Secretory Asp-49 and Lys-49 Phospholipases A(2). Toxicon 2005, 46, 523–532. [Google Scholar] [CrossRef]
- Leiguez, E.; Zuliani, J.P.; Cianciarullo, A.M.; Fernandes, C.M.; Gutierrez, J.M.; Teixeira, C. A Group IIA-Secreted Phospholipase A2 from Snake Venom Induces Lipid Body Formation in Macrophages: The Roles of Intracellular Phospholipases A2 and Distinct Signaling Pathways. J. Leukoc. Biol. 2011, 90, 155–166. [Google Scholar] [CrossRef]
- Boeno, C.N.; Paloschi, M.V.; Lopes, J.A.; Souza Silva, M.D.; Evangelista, J.R.; dos Reis, V.P.; da Setúbal, S.S.; Soares, A.M.; Zuliani, J.P. Dynamics of Action of a Lys-49 and an Asp-49 PLA2s on Inflammasome NLRP3 Activation in Murine Macrophages. Int. Immunopharmacol. 2022, 112, 109194. [Google Scholar] [CrossRef]
- Landucci, E.C.T.; Castro, R.C.; Pereira, M.F.; Cintra, A.C.O.; Giglio, J.R.; Marangoni, S.; Oliveira, B.; Cirino, G.; Antunes, E.; de Nucci, G. Mast Cell Degranulation Induced by Two Phospholipase A2 Homologues: Dissociation between Enzymatic and Biological Activities. Eur. J. Pharmacol. 1998, 343, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, P.; Bao, Y.; Serrano, S.M.T.; Laing, G.D.; Theakston, R.D.G.; Gutiérrez, J.M.; Escalante, T.; Zigrino, P.; Moura-Da-Silva, A.M.; Nischt, R.; et al. Role of the Snake Venom Toxin Jararhagin in Proinflammatory Pathogenesis: In Vitro and in Vivo Gene Expression Analysis of the Effects of the Toxin. Arch. Biochem. Biophys. 2005, 441, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Janovits, P.M.; Leiguez, E.; Portas, V.; Teixeira, C. A Metalloproteinase Induces an Inflammatory Response in Preadipocytes with the Activation of COX Signalling Pathways and Participation of Endogenous Phospholipases A2. Biomolecules 2021, 11, 921. [Google Scholar] [CrossRef] [PubMed]
- Viana, M.N.; Leiguez, E.; Gutiérrez, J.M.; Rucavado, A.; Markus, R.P.; Marçola, M.; Teixeira, C.; Fernandes, C.M. A Representative Metalloprotease Induces PGE2 Synthesis in Fibroblast-like Synoviocytes via the NF-ΚB/COX-2 Pathway with Amplification by IL-1β and the EP4 Receptor. Sci. Rep. 2020, 10, 3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, J.M.; Rucavado, A.; Escalante, T.; Lomonte, B.; Angulo, Y.; Fox, J.W. Tissue Pathology Induced by Snake Venoms: How to Understand a Complex Pattern of Alterations from a Systems Biology Perspective? Toxicon 2010, 55, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Marx, V. Method of the Year: Spatially Resolved Transcriptomics. Nat. Methods 2021, 18, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Merritt, C.R.; Ong, G.T.; Church, S.E.; Barker, K.; Danaher, P.; Geiss, G.; Hoang, M.; Jung, J.; Liang, Y.; McKay-Fleisch, J.; et al. Multiplex Digital Spatial Profiling of Proteins and RNA in Fixed Tissue. Nat. Biotechnol. 2020, 38, 586–599. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Rucavado, A.; Chaves, F.; Díaz, C.; Escalante, T. Experimental Pathology of Local Tissue Damage Induced by Bothrops asper Snake Venom. Toxicon 2009, 54, 958–975. [Google Scholar] [CrossRef]
- Mora, R.; Valverde, B.; Díaz, C.; Lomonte, B.; Gutiérrez, J.M. A Lys49 Phospholipase A(2) Homologue from Bothrops asper Snake Venom Induces Proliferation, Apoptosis and Necrosis in a Lymphoblastoid Cell Line. Toxicon 2005, 45, 651–660. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Chaves, F.; Cerdas, L. Inflammatory Infiltrate in Skeletal Muscle Injected with Bothrops asper Venom—PubMed. Rev. Biol. Trop. 1986, 34, 209–214. [Google Scholar]
- Brigitte, M.; Schilte, C.; Plonquet, A.; Baba-Amer, Y.; Henri, A.; Charlier, C.; Tajbakhsh, S.; Albert, M.; Gherardi, R.K.; Chrétien, F. Muscle Resident Macrophages Control the Immune Cell Reaction in a Mouse Model of Notexin-Induced Myoinjury. Arthritis Rheum. 2010, 62, 268–279. [Google Scholar] [CrossRef]
- Mortaz, E.; Alipoor, S.D.; Adcock, I.M.; Mumby, S.; Koenderman, L. Update on Neutrophil Function in Severe Inflammation. Front. Immunol. 2018, 9, 2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, P.X.; Kubes, P. The Neutrophil’s Role During Health and Disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.F.P.; Zamunér, S.R.; Zuliani, J.P.; Fernandes, C.M.; Cruz-Hofling, M.A.; Fernandes, I.; Chaves, F.; Gutiérrez, J.M. Neutrophils Do Not Contribute to Local Tissue Damage, but Play a Key Role in Skeletal Muscle Regeneration, in Mice Injected with Bothrops asper Snake Venom. Muscle Nerve 2003, 28, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Arnold, L.; Henry, A.; Poron, F.; Baba-Amer, Y.; van Rooijen, N.; Plonquet, A.; Gherardi, R.K.; Chazaud, B. Inflammatory Monocytes Recruited after Skeletal Muscle Injury Switch into Antiinflammatory Macrophages to Support Myogenesis. J. Exp. Med. 2007, 204, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- Freitas, A.P.; Clissa, P.B.; Soto, D.R.; Câmara, N.O.S.; Faquim-Mauro, E.L. The Modulatory Effect of Crotoxin and Its Phospholipase A2 Subunit from Crotalus durissus terrificus Venom on Dendritic Cells Interferes with the Generation of Effector CD4+ T Lymphocytes. Immunol. Lett. 2021, 240, 56–70. [Google Scholar] [CrossRef]
- Page, G.; Chevrel, G.; Miossec, P. Anatomic Localization of Immature and Mature Dendritic Cell Subsets in Dermatomyositis and Polymyositis: Interaction with Chemokines and Th1 Cytokine-Producing Cells. Arthritis Rheum. 2004, 50, 199–208. [Google Scholar] [CrossRef]
- Greenberg, S.A.; Pinkus, G.S.; Amato, A.A.; Pinkus, J.L. Myeloid Dendritic Cells in Inclusion-Body Myositis and Polymyositis. Muscle Nerve 2007, 35, 17–23. [Google Scholar] [CrossRef]
- Ladislau, L.; Portilho, D.M.; Courau, T.; Solares-Pérez, A.; Negroni, E.; Lainé, J.; Klatzmann, D.; Bonomo, A.; Allenbach, Y.; Benveniste, O.; et al. Activated Dendritic Cells Modulate Proliferation and Differentiation of Human Myoblasts. Cell Death Dis. 2018, 9, 551. [Google Scholar] [CrossRef]
- Dirks, A.J.; Leeuwenburgh, C. The Role of Apoptosis in Age-Related Skeletal Muscle Atrophy. Sport. Med. 2005, 35, 473–483. [Google Scholar] [CrossRef]
- Marzetti, E.; Leeuwenburgh, C. Skeletal Muscle Apoptosis, Sarcopenia and Frailty at Old Age. Exp. Gerontol. 2006, 41, 1234–1238. [Google Scholar] [CrossRef]
- Lukjanenko, L.; Jung, M.J.; Hegde, N.; Perruisseau-Carrier, C.; Migliavacca, E.; Rozo, M.; Karaz, S.; Jacot, G.; Schmidt, M.; Li, L.; et al. Loss of Fibronectin from the Aged Stem Cell Niche Affects the Regenerative Capacity of Skeletal Muscle in Mice. Nat. Med. 2016, 22, 897–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, J.; Arce, V.; Brenes, F.; Chaves, F. Changes in Myofibrillar Components after Skeletal Muscle Necrosis Induced by a Myotoxin Isolated from the Venom of the Snake Bothrops asper. Exp. Mol. Pathol. 1990, 52, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Vater, R.; Cullen, M.J.; Harris, J.B. The Fate of Desmin and Titin during the Degeneration and Regeneration of the Soleus Muscle of the Rat. Acta Neuropathol. 1992, 84, 278–288. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, A.K.; Pramoonjago, P.; Rucavado, A.; Moskaluk, C.; Silva, D.T.; Escalante, T.; Gutiérrez, J.M.; Fox, J.W. Mapping the Immune Cell Microenvironment with Spatial Profiling in Muscle Tissue Injected with the Venom of Daboia russelii. Toxins 2023, 15, 208. https://doi.org/10.3390/toxins15030208
de Oliveira AK, Pramoonjago P, Rucavado A, Moskaluk C, Silva DT, Escalante T, Gutiérrez JM, Fox JW. Mapping the Immune Cell Microenvironment with Spatial Profiling in Muscle Tissue Injected with the Venom of Daboia russelii. Toxins. 2023; 15(3):208. https://doi.org/10.3390/toxins15030208
Chicago/Turabian Stylede Oliveira, Ana K., Patcharin Pramoonjago, Alexandra Rucavado, Christopher Moskaluk, Dilza T. Silva, Teresa Escalante, José María Gutiérrez, and Jay W. Fox. 2023. "Mapping the Immune Cell Microenvironment with Spatial Profiling in Muscle Tissue Injected with the Venom of Daboia russelii" Toxins 15, no. 3: 208. https://doi.org/10.3390/toxins15030208