Gut-Faecal Microbial and Health-Marker Response to Dietary Fumonisins in Weaned Pigs

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Growth Rate and Clinical Signs
2.2. Serum Biochemical Endpoints
2.3. Antioxidant Parameters
2.4. Histopathological Findings
- 0 = no alteration, healthy condition;
- 1 = slight/small scale/few extent of pathological alteration;
- 2 = medium degree/medium scale/medium number of pathological modification;
- 3 = pronounced/extensive/numerous extent or occurrence of the alterations.
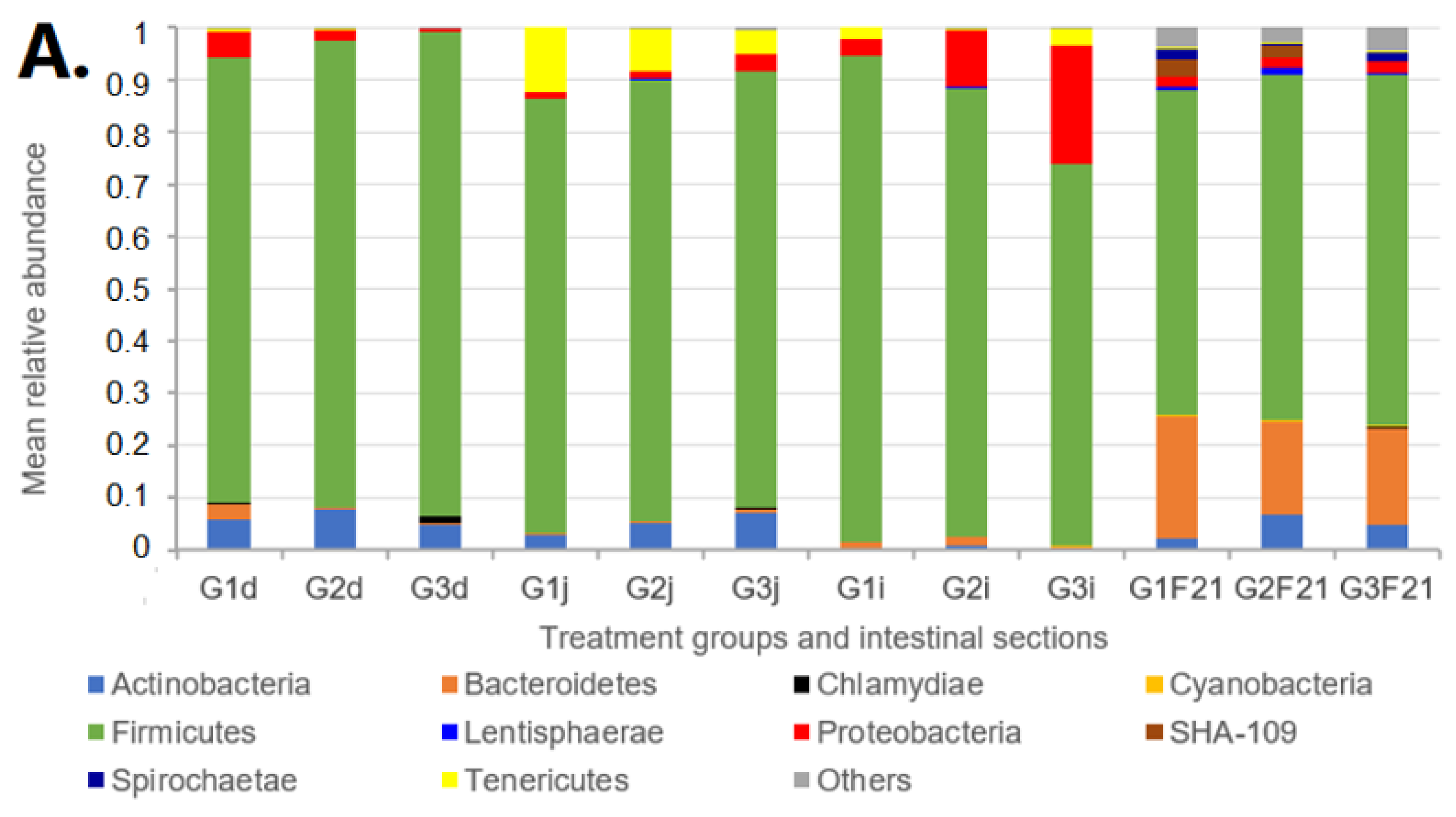
2.5. Microbiota Analyses
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals, Housing and Experimental Diets
5.2. Sampling and Laboratory Analysis
5.2.1. Blood Serum Biochemistry
5.2.2. Determination of Antioxidant Parameters and Lipid Peroxidation
5.2.3. Histology and Histopathology
- 0 = no alteration, healthy condition;
- 1 = slight/small scale/few extent of pathological alteration;
- 2 = medium degree/medium scale/medium number of pathological modification;
- 3 = pronounced/extensive/numerous extent or occurrence of the alterations.
5.2.4. Statistical Analysis
5.2.5. Microbiota Analysis and Statistical Analysis Procedure
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bolger, M.; Coker, R.D.; DiNovi, M.; Gaylor, D.; Gelderblom, W.; Olsen, M.; Paster, N.; Riley, R.T.; Shephard, G.; Speijers, G.J.A. Fumonisins. In Safety Evaluation of Certain Mycotoxins in Food; WHO Food Additives Series; WHO: Geneva, Switzerland, 2001; Volume 47, pp. 103–297. [Google Scholar]
- Bulder, A.S.; Arcella, D.; Bolger, M.; Carrington, C.; Kpodo, K.; Resnik, S.; Riley, R.T.; Wolterink, G.; Wu, F. Fumonisins (Addendum). Saf. Eval. Certain Food Addit. Contam. 2012, 65, 325–794. [Google Scholar]
- Szecsi, A.; Szekeres, A.; Bartok, T.; Oros, G.; Bartok, M.; Mesterhazy, A. Fumonisin B1-4-Producing Capacity of Hungarian Fusarium Verticillioides Isolates. World Mycotoxin J. 2010, 3, 67–76. [Google Scholar] [CrossRef]
- Lerda, D. Fumonisins in Foods from Cordoba (Argentina), Presence: Mini Review. Toxicol Open Access 2017, 3, 125. [Google Scholar] [CrossRef]
- Smith, G.W. Fumonisins. In Veterinary Toxicology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1003–1018. [Google Scholar]
- Harrison, L.R.; Colvin, B.M.; Greene, J.T.; Newman, L.E.; Cole, J.R. Pulmonary Edema and Hydrothorax in Swine Produced by Fumonisin B1, a Toxic Metabolite of Fusarium moniliforme. J. Vet. Diagn. Investig. 1990, 2, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Gelderblom, W.C.A.; Jaskiewicz, K.; Marasas, W.F.O.; Thiel, P.G.; Horak, R.M.; Vleggaar, R.; Kriek, N.P.J. Fumonisins—Novel Mycotoxins with Cancer-Promoting Activity Produced by Fusarium moniliforme. Appl. Environ. Microbiol. 1988, 54, 1806–1811. [Google Scholar] [CrossRef] [PubMed]
- Gelderblom, W.C.A.; Kriek, N.P.J.; Marasas, W.F.O.; Thiel, P.G. Toxicity and Carcinogenicity of the Fusarium moniliforme Metabolite, Fumonisin B1, in Rats. Carcinogenesis 1991, 12, 1247–1251. [Google Scholar] [CrossRef]
- Voss, K.A.; Chamberlain, W.J.; Bacon, C.W.; Norred, W.P. A Preliminary Investigation on Renal and Hepatic Toxicity in Rats Fed Purified Fumonisin B1. Nat. Toxins 1993, 1, 222–228. [Google Scholar] [CrossRef]
- Chilaka, C.A.; De Boevre, M.; Atanda, O.O.; De Saeger, S. The Status of Fusarium Mycotoxins in Sub-Saharan Africa: A Review of Emerging Trends and Post-Harvest Mitigation Strategies towards Food Control. Toxins 2017, 9, 19. [Google Scholar] [CrossRef]
- Ortiz, C.S.; Richards, C.; Terry, A.; Parra, J.; Shim, W.B. Genetic Variability and Geographical Distribution of Mycotoxigenic Fusarium verticillioides Strains Isolated from Maize Fields in Texas. Plant Pathol. J. 2015, 31, 203–211. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Fumonisin B1. IARC. Monogr. Eval. Carcinog. Risk Hum. 2002, 82, 275–366. [Google Scholar]
- Bouhet, S.; Oswald, I.P. The Intestine as a Possible Target for Fumonisin Toxicity. Mol. Nutr. Food Res. 2007, 51, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Schwartz-Zimmermann, H.E.; Caha, S.; Moll, W.D.; Schatzmayr, G.; Applegate, T.J. Dose-Dependent Effects on Sphingoid Bases and Cytokines in Chickens Fed Diets Prepared with Fusarium verticillioides Culture Material Containing Fumonisins. Toxins 2015, 7, 1253–1272. [Google Scholar] [CrossRef] [PubMed]
- Swamy, H.V.L.N.; Smith, T.K.; MacDonald, E.J.; Karrow, N.A.; Woodward, B.; Boermans, H.J. Effects of Feeding a Blend of Grains Naturally Contaminated with Fusarium Mycotoxins on Growth and Immunological Measurements of Starter Pigs, and the Efficacy of a Polymeric Glucomannan Mycotoxin Adsorbent. J. Anim. Sci. 2003, 81, 2792–2803. [Google Scholar] [CrossRef] [PubMed]
- Gbore, F.A.; Yinusa, R.I. Salleh Evaluation of Subchronic Dietary Fumonisin B 1 on Nutrient Digestibility and Growth Performance of Rats. Afr. J. Biotechnol. 2010, 9, 6442–6447. [Google Scholar] [CrossRef]
- Lessard, M.; Boudry, G.; Sève, B.; Oswald, I.P.; Lallès, J.P. Intestinal Physiology and Peptidase Activity in Male Pigs Are Modulated by Consumption of Corn Culture Extracts Containing Fumonisins. J. Nutr. 2009, 139, 1303–1307. [Google Scholar] [CrossRef] [PubMed]
- Gbore, F.A.; Egbunike, G.N. Influence of Dietary Fumonisin B1 on Nutrient Utilization by Growing Pigs. Livest. Res. Rural Dev. 2007, 19, 93. [Google Scholar]
- Conway, P.L. Function and Regulation of the Gastrointestinal Microbiota of the Pig. Publ. Assoc. Anim. Prod. 1994, 80, 231. [Google Scholar]
- Ren, Z.; Guo, C.; Yu, S.; Zhu, L.; Wang, Y.; Hu, H.; Deng, J. Progress in Mycotoxins Affecting Intestinal Mucosal Barrier Function. Int. J. Mol. Sci. 2019, 20, 2777. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Kim, I.H. The Impact of Weaning Stress on Gut Health and the Mechanistic Aspects of Several Feed Additives Contributing to Improved Gut Health Function in Weanling Piglets—A Review. Animals 2021, 11, 2418. [Google Scholar] [CrossRef]
- Wei, X.; Tsai, T.; Howe, S.; Zhao, J. Weaning Induced Gut Dysfunction and Nutritional Interventions in Nursery Pigs: A Partial Review. Animals 2021, 11, 1279. [Google Scholar] [CrossRef]
- Wang, J.; Tang, L.; Glenn, T.C.; Wang, J.S. Aflatoxin B1 Induced Compositional Changes in Gut Microbial Communities of Male F344 rats. Toxicol. Sci. 2016, 150, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yang, S.; Zhang, X.; Huang, S.; Wang, N.; Wang, M.; Long, M.; He, J. Zearalenone Changes the Diversity and Composition of Caecum Microbiota in Weaned Rabbit. BioMed Res. Int. 2018, 2018, 3623274. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.W.; Dinesh, R.; Yoshinari, T.; Holtrop, G.; Richardson, A.J.; Duncan, G.; MacDonald, S.; Lloyd, A.; Tarbin, J. Masked Trichothecene and Zearalenone Mycotoxins Withstand Digestion and Absorption in the Upper GI Tract but Are Efficiently Hydrolyzed by Human Gut Microbiota in Vitro. Mol. Nutr. Food Res. 2017, 61, 1600680. [Google Scholar] [CrossRef] [PubMed]
- Oswald, I.P.; Desautels, C.; Laffitte, J.; Fournout, S.; Peres, S.Y.; Odin, M.; Le, P.; Le Bars, J.; Fairbrother, J.M. Mycotoxin Fumonisin B 1 Increases Intestinal Colonization by Pathogenic Escherichia Coli in Pigs. Appl. Environ. Microbiol. 2003, 69, 5870–5874. [Google Scholar] [CrossRef] [PubMed]
- Burel, C.; Tanguy, M.; Guerre, P.; Boilletot, E.; Cariolet, R.; Queguiner, M.; Postollec, G.; Pinton, P.; Salvat, G.; Oswald, I.P.; et al. Effect of Low Dose of Fumonisins on Pig Health: Immune Status, Intestinal Microbiota and Sensitivity to Salmonella. Toxins 2013, 5, 841–864. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, Z.; Jiang, L.; Chen, Z.; Sun, H. Response of Fecal Bacterial Flora to the Exposure of Fumonisin B1 in Balb/c Mice. Toxins 2021, 13, 612. [Google Scholar] [CrossRef]
- Yu, S.; Jia, B.; Lin, H.; Zhang, S.; Yu, D.; Liu, N.; Wu, A. Effects of Fumonisin B and Hydrolyzed Fumonisin B on Growth and Intestinal Microbiota in Broilers. Toxins 2022, 14, 163. [Google Scholar] [CrossRef]
- Zeebone, Y.Y.; Kovács, M.; Bóta, B.; Zdeněk, V.; Taubner, T.; Halas, V. Dietary Fumonisin May Compromise the Nutritive Value of Feed and Distort Copper and Zinc Digestibility and Retention in Weaned Piglets. J. Anim. Physiol. Anim. Nutr. 2022, 107, 504–517. [Google Scholar] [CrossRef]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Risks for Animal Health Related to the Presence of Fumonisins, Their Modified Forms and Hidden Forms in Feed. EFSA J. 2018, 16, e05242. [Google Scholar] [CrossRef]
- Terciolo, C.; Bracarense, A.P.; Souto, P.C.M.C.; Cossalter, A.M.; Dopavogui, L.; Loiseau, N.; Oliveira, C.A.F.; Pinton, P.; Oswald, I.P. Fumonisins at Doses below EU Regulatory Limits Induce Histological Alterations in Piglets. Toxins 2019, 11, 548. [Google Scholar] [CrossRef]
- Haschek, W.M.; Gumprecht, L.A.; Smith, G.; Tumbleson, M.E.; Constable, P.D. Fumonisin Toxicosis in Swine: An Overview of Porcine Pulmonary Edema and Current Perspectives. Environ. Health Perspect. 2001, 109, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Colvin, B.M.; Cooley, A.J.; Beaver, R.W. Fumonisin Toxicosis in Swine: Clinical and Pathologic Findings. J. Vet. Diagn. Investig. 1993, 5, 232–541. [Google Scholar] [CrossRef] [PubMed]
- Schertz, H.; Dänicke, S.; Frahm, J.; Schatzmayr, D.; Dohnal, I.; Bichl, G.; Schwartz-Zimmermann, H.E.; Colicchia, S.; Breves, G.; Teifke, J.P.; et al. Biomarker Evaluation and Toxic Effects of an Acute Oral and Systemic Fumonisin Exposure of Pigs with a Special Focus on Dietary Fumonisin Esterase Supplementation. Toxins 2018, 10, 296. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.J.; Harvey, J.W.; Bruss, M.L. Clinical Biochemistry of Domestic Animals; Academic Press: Cambridge, MA, USA, 2008; ISBN 0080568823. [Google Scholar]
- Dilkin, P.; Direito, G.; Simas, M.M.S.; Mallmann, C.A.; Corrêa, B. Toxicokinetics and Toxicological Effects of Single Oral Dose of Fumonisin B1 Containing Fusarium verticillioides Culture Material in Weaned Piglets. Chem. Biol. Interact. 2010, 185, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Fodor, J.; Meyer, K.; Riedlberger, M.; Bauer, J.; Horn, P.; Kovacs, F.; Kovacs, M. Distribution and Elimination of Fumonisin Analogues in Weaned Piglets after Oral Administration of Fusarium verticillioides Fungal Culture. Food Addit. Contam. 2006, 23, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Kócsó, D.J.; Szabó-Fodor, J.; Mézes, M.; Balogh, K.; Ferenczi, S.; Szabó, A.; Bóta, B.; Kovács, M. Fumonisin B 1 Exposure Increases Hsp70 Expression in the Lung and Kidney of Rats without Inducing Significant Oxidative Stress. Acta Vet. Hung. 2018, 66, 394–407. [Google Scholar] [CrossRef] [PubMed]
- Kubasova, T.; Davidova-Gerzova, L.; Babak, V.; Cejkova, D.; Montagne, L.; Le-Floc’h, N.; Rychlik, I. Effects of Host Genetics and Environmental Conditions on Fecal Microbiota Composition of Pigs. PLoS ONE 2018, 13, e0201901. [Google Scholar] [CrossRef]
- Che, D.; Adams, S.; Wei, C.; Gui-Xin, Q.; Atiba, E.M.; Hailong, J. Effects of Astragalus Membranaceus Fiber on Growth Performance, Nutrient Digestibility, Microbial Composition, VFA Production, Gut PH, and Immunity of Weaned Pigs. Microbiologyopen 2019, 8, e00712. [Google Scholar] [CrossRef]
- Gebhardt, J.T.; Thomson, K.A.; Woodworth, J.C.; Dritz, S.S.; Tokach, M.D.; Derouchey, J.M.; Goodband, R.D.; Jones, C.K.; Cochrane, R.A.; Niederwerder, M.C.; et al. Effect of Dietary Medium-Chain Fatty Acids on Nursery Pig Growth Performance, Fecal Microbial Composition, and Mitigation Properties against Porcine Epidemic Diarrhea Virus Following Storage. J. Anim. Sci. 2020, 98, skz358. [Google Scholar] [CrossRef]
- Kim, H.B.; Borewicz, K.; White, B.A.; Singer, R.S.; Sreevatsan, S.; Tu, Z.J.; Isaacson, R.E. Longitudinal Investigation of the Age-Related Bacterial Diversity in the Feces of Commercial Pigs. Vet. Microbiol. 2011, 153, 124–133. [Google Scholar] [CrossRef]
- Looft, T.; Johnson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D. In-Feed Antibiotic Effects on the Swine Intestinal Microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef] [PubMed]
- Ramayo-Caldas, Y.; Mach, N.; Lepage, P.; Levenez, F.; Denis, C.; Lemonnier, G.; Leplat, J.J.; Billon, Y.; Berri, M.; Doré, J.; et al. Phylogenetic Network Analysis Applied to Pig Gut Microbiota Identifies an Ecosystem Structure Linked with Growth Traits. ISME J. 2016, 10, 2973–2977. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Downes, J.; Dewhirst, F.E.; Tanner, A.C.R.; Wade, W.G. Description of Alloprevotella rava gen. nov., sp. Nov., Isolated from the Human Oral Cavity, and Reclassification of Prevotella tannerae Moore et Al. 1994 as Alloprevotella tannerae gen. nov., comb. Nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 1214–1218. [Google Scholar] [CrossRef]
- Ren, W.; Chen, S.; Zhang, L.; Liu, G.; Hussain, T.; Hao, X.; Yin, J.; Duan, J.; Tan, B.; Wu, G.; et al. Interferon Tau Affects Mouse Intestinal Microbiota and Expression of IL-17. Mediat. Inflamm. 2016, 2016, 2839232. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Oliphant, K.; Allen-Vercoe, E. Macronutrient Metabolism by the Human Gut Microbiome: Major Fermentation By-Products and Their Impact on Host Health. Microbiome 2019, 7, 91. [Google Scholar] [CrossRef]
- Zhong, X.; Zhang, Z.; Wang, S.; Cao, L.; Zhou, L.; Sun, A.; Zhong, Z.; Nabben, M. Microbial-Driven Butyrate Regulates Jejunal Homeostasis in Piglets during the Weaning Stage. Front. Microbiol. 2019, 10, 3335. [Google Scholar] [CrossRef]
- Dou, S.; Gadonna-Widehem, P.; Rome, V.; Hamoudi, D.; Rhazi, L.; Lakhal, L.; Larcher, T.; Bahi-Jaber, N.; Pinon-Quintana, A.; Guyonvarch, A.; et al. Characterisation of Early-Life Fecal Microbiota in Susceptible and Healthy Pigs to Post-Weaning Diarrhoea. PLoS ONE 2017, 12, e0169851. [Google Scholar] [CrossRef]
- Mateos, I.; Combes, S.; Pascal, G.; Cauquil, L.; Barilly, C.; Cossalter, A.M.; Laffitte, J.; Botti, S.; Pinton, P.; Oswald, I.P. Fumonisin-Exposure Impairs Age-Related Ecological Succession of Bacterial Species in Weaned Pig Gut Microbiota. Toxins 2018, 10, 230. [Google Scholar] [CrossRef]
- De Angelis, M.; Calasso, M.; Cavallo, N.; Di Cagno, R.; Gobbetti, M. Functional Proteomics within the Genus Lactobacillus. Proteomics 2016, 16, 946–962. [Google Scholar] [CrossRef] [PubMed]
- Gresse, R.; Durand, F.C.; Dunière, L.; Blanquet-Diot, S.; Forano, E. Microbiota Composition and Functional Profiling throughout the Gastrointestinal Tract of Commercial Weaning Piglets. Microorganisms 2019, 7, 343. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.; Koh, S.; Oh, Y.; Cho, H. Exposure to Low Concentrations of Mycotoxins Triggers Unique Responses from the Pig Gut Microbiome. Korean J. Vet. Serv. 2020, 43, 39–44. [Google Scholar]
- Dang, H.A.; Zsolnai, A.; Kovacs, M.; Bors, I.; Bonai, A.; Bota, B.; Szabóoo-Fodor, J. In Vitro Interaction between Fumonisin B1 and the Intestinal Microflora of Pigs. Pol. J. Microbiol. 2017, 66, 245–250. [Google Scholar] [CrossRef]
- Kim, S.W.; Holanda, D.M.; Gao, X.; Park, I.; Yiannikouris, A. Effcacy of a Yeast Cellwall Extract to Mitigate the Effect of Naturally Co-Occurring Mycotoxins Contaminating Feed Ingredients Fed to Young Pigs: Impact on Gut Health, Microbiome, and Growth. Toxins 2019, 11, 633. [Google Scholar] [CrossRef]
- De Angelis, M.; Siragusa, S.; Berloco, M.; Caputo, L.; Settanni, L.; Alfonsi, G.; Amerio, M.; Grandi, A.; Ragni, A.; Gobbetti, M. Selection of Potential Probiotic Lactobacilli from Pig Feces to Be Used as Additives in Pelleted Feeding. Res. Microbiol. 2006, 157, 792–801. [Google Scholar] [CrossRef]
- Ahlberg, S.H.; Joutsjoki, V.; Korhonen, H.J. Potential of lactic acid bacteria in aflatoxin risk mitigation. Int. J. Food. Microbiol. 2015, 207, 87–102. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, X.; Zhang, J.; Zhang, J.; Zhang, B. The Mechanism of Lactobacillus Strains for Their Ability to Remove Fumonisins B1 and B2. Food Chem. Toxicol. 2016, 97, 40–46. [Google Scholar] [CrossRef]
- Festa, S.; Coppotelli, B.M.; Morelli, I.S. Bacterial Diversity and Functional Interactions between Bacterial Strains from a Phenanthrene-Degrading Consortium Obtained from a Chronically Contaminated-Soil. Int. Biodeterior. Biodegrad. 2013, 85, 42–51. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y.; Gong, A.; Liu, N.; Chen, S.; Zhao, X.; Li, X.; Chen, L.; Zhou, C.; Wang, J. Biodegradation of Mycotoxin Fumonisin B1 by a Novel Bacterial Consortium SAAS79. Appl. Microbiol. Biotechnol. 2019, 103, 7129–7140. [Google Scholar] [CrossRef]
- Ali, O.; Mézes, M.; Balogh, K.; Kovács, M.; Turbók, J.; Szabó, A. Fumonisin B Series Mycotoxins’ Dose Dependent Effects on the Porcine Hepatic and Pulmonary Phospholipidome. Toxins 2022, 14, 803. [Google Scholar] [CrossRef] [PubMed]
- Zeebone, Y.Y.; Kovács, M.; Bóta, B.; Halas, V. Effects of Dietary Fumonisins on Nutrients Digestibility in Weanling Pigs. Acta Fytotech. Zootech. 2020, 23, 23–28. [Google Scholar] [CrossRef]
- Fodor, J.; Kametier, L.; Kovács, M. Practical Aspects of Fumonisin Production under Laboratory Conditions. Mycotoxin Res. 2006, 22, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Botsoglou, N.A.; Fletouris, D.J.; Papageorgiou, G.E.; Vassilopoulos, V.N.; Mantis, A.J.; Trakatellis, A.G. Rapid, Sensitive, and Specific Thiobarbituric Acid Method for Measuring Lipid Peroxidation in Animal Tissue, Food, and Feedstuff Samples. J. Agric. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of Total, Protein-Bound, and Nonprotein Sulfhydryl Groups in Tissue with Ellman’s Reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.A.; Burk, R.F. Species, Tissue and Subcellular Distribution of Non-Se-Dependent Glutathione Peroxidase Activity. J. Nutr. 1978, 108, 211–215. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Bushnell, B.; Rood, J.; Singer, E. BBMerge—Accurate Paired Shotgun Read Merging via Overlap. PLoS ONE 2017, 12, e0185056. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Ju, F.; Zhang, T. 16S RRNA Gene High-Throughput Sequencing Data Mining of Microbial Diversity and Interactions. Appl. Microbiol. Biotechnol. 2015, 99, 4119–4129. [Google Scholar] [CrossRef] [PubMed]
- Hazra, A.; Gogtay, N. Biostatistics Series Module 3: Comparing Groups: Numerical Variables. Indian J. Dermatol. 2016, 61, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Xu, Z.Z.; Peddada, S.; Amir, A.; Bittinger, K.; Gonzalez, A.; Lozupone, C.; Zaneveld, J.R.; Vázquez-Baeza, Y.; Birmingham, A.; et al. Normalization and Microbial Differential Abundance Strategies Depend upon Data Characteristics. Microbiome 2017, 5, 27. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Sun, J. ScienceDirect Hypothesis Testing and Statistical Analysis of Microbiome. Genes Dis. 2017, 4, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xu, Y.; Chen, X.; Fang, C.; Zhao, L.; Chen, F. The Maturing Development of Gut Microbiota in Commercial Piglets during the Weaning Transition. Front. Microbiol. 2017, 8, 1688. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Serum Parameters | Control (G1) | 15 mg/kg FBs (G2) | 30 mg/kg FBs (G3) | p-Value |
|---|---|---|---|---|
| Total protein, g/L | 56.2 ± 2.7 a | 56.8 ± 2.2 a | 60.9 ± 2.8 b | 0.012 |
| Albumin, g/L | 35.6 ± 3.6 | 34.4 ± 1.8 | 35.7 ± 3.8 | 0.703 |
| AST, U/L | 57.5 ± 18.2 a | 87.8 ± 22.9 a | 336.5 ± 269.6 b | 0.001 |
| ALT, U/L | 65.7 ± 16.7 | 86.3 ± 22.89 | 100.8 ± 55.3 | 0.117 |
| GGT, U/L | 37 ± 7.0 a | 53.2 ± 18.5 ab | 122.8 ± 102.2 b | 0.011 |
| ALKP, U/L | 270.7 ± 40.6 a | 406 ± 212.3 ab | 1356.5 ± 1384.6 b | 0.008 |
| CK, U/L | 1382.5 ± 686.5 a | 1351.5 ± 852.2 a | 4449.8 ± 3315.5 b | 0.019 |
| LDH, U/L | 1192 ± 165.7 a | 1323.8 ± 212.7 a | 2272.3 ± 693.7 b | 0.003 |
| GFR, ml/min | 90 ± 0.0 a | 88.7 ± 3.3 a | 76.4 ± 12.6 b | 0.029 |
| Urea, mmol/L | 3.3 ± 0.6 | 4.3 ± 0.9 | 3.9 ± 0.1 | 0.212 |
| Cholesterol, mmol/L | 2.2 ± 0.1 a | 3 ± 0.5 a | 4.5 ± 0.9 b | 0.001 |
| Creatinine, µmol/L | 88 ± 8.17 a | 88 ± 8.17 a | 102.3 ± 4.5 b | 0.019 |
| Ca, mmol/L | 2.7 ± 0.13 a | 2.7 ± 0.18 ab | 2.9 ± 0.11 b | 0.029 |
| Mg, mmol/L | 1.0 ± 0.12 a | 1.0 ± 0.10 a | 1.2 ± 0.12 b | 0.018 |
| Fe, mmol/L | 26.1 ± 4.8 | 21.7 ± 3.4 | 24.1 ± 6.7 | 0.353 |
| Na, mmol/L | 147.3 ± 4.1 | 147 ± 2.6 | 143.5 ± 3.0 | 0.116 |
| Cl, mmol/L | 101.5 ± 5.7 | 116.8 ± 41.3 | 97.3 ± 2.9 | 0.183 |
| Organ Pathological Observation | Control (G1) | 15 mg/kg FBs (G2) | 30 mg/kg FBs (G3) | |
|---|---|---|---|---|
| n | ∑ | |||
| Liver | ||||
| Decrease of staining intensity of liver cells | 0 | 6 | 6 | 12 |
| Single liver cell death | 0 | 6 | 6 | 12 |
| Swelling of MPS cells | 0 | 6 | 6 | 12 |
| Proliferation of MPS cells | 0 | 6 | 6 | 12 |
| Kidney | ||||
| Tubular epithelium detachment | 0 | 0 | 6 | 6 |
| Lymphocytic infiltration | 1 | 1 | 6 | 8 |
| Lung | ||||
| Interstitial lymphocytic infiltration | 2 | 3 | 4 | 9 |
| Pleural fibrosis | 0 | 2 | 3 | 5 |
| Ileum | ||||
| GALT lymphocyte depletion | 0 | 0 | 3 | 3 |
| Sample Origin | Genus-Level Colonization Pattern | Group Effects | p-Value |
|---|---|---|---|
| Duodenum | Bacteroidales: Prevotellaceae: Alloprevotella | G1G3 ↓ | 0.013 |
| Duodenum | Campylobacteraceae: Campylobacter | G1G3, G2G3 ↓ | 0.016, 0.012 |
| Duodenum | Firmicutes: Clostridiales: Lachnospiraceae: Incertae Sedis | G1G3, G2G3 ↓ | 0.009, 0.035 |
| Jejunum | Firmicutes: Erysipelotrichaceae: Turicibacter | G1G3 ↓ | 0.001 |
| Ileum | Clostridiales: Clostridiaceae: Clostridium sensu stricto 1. | G1G3 ↓ | 0.009 |
| Faeces | Firmicutes: Erysipelotrichaceae: Solobacterium | G1G3, G2G3 ↑ | 0.040, 0.003 |
| Faeces | Firmicutes: Clostridiales: Ruminococcaceae: Faecalibacterium | G2G3 ↑ | 0.020 |
| Faeces | Firmicutes: Clostridiales: Ruminococcaceae: Anaerofilum | G1G3, G2G3 ↑ | 0.027, 0.008 |
| Faeces | Firmicutes: Clostridiales: Ruminococcaceae: Ruminococcus | G2G3 ↑ | 0.011 |
| Faeces | Firmicutes: Clostridiales: Ruminococcaceae: Subdoligranulum | G2G3 ↑ | 0.015 |
| Faeces | Firmicutes: Clostridiales: Lachnospiraceae: Pseudobutyrivibrio | G1G3, G2G3 ↑ | 0.031, 0.027 |
| Faeces | Firmicutes: Clostridiales: Lachnospiraceae: Coprococcus | G2G3 ↑ | 0.04 |
| Faeces | Firmicutes: Clostridiales: Lachnospiraceae: Roseburia | G2G3 ↑ | 0.011 |
| Item, g/kg | |
| Crude protein | 175 |
| Crude fat | 33 |
| Crude fibre | 37 |
| Crude ash | 50 |
| Starch | 418 |
| Lysine | 11.1 |
| Methionine | 3.7 |
| Ca | 6.5 |
| P | 5.0 |
| Item, mg/kg | |
| Manganese | 40 |
| Zinc | 110 |
| Iron | 87 |
| Copper | 9.4 |
| Selenium | 0.30 |
| Iodine | 0.6 |
| DE, MJ/kg | 14.70 |
| ME, MJ/kg | 14.10 |
| Mycotoxins | Quantification (m/z) | Confirmation (m/z) |
|---|---|---|
| FB1 | 722.4 (+) | 760.3 (+), 720.3 (−) |
| FB2 | 706.4 (+) | 744.3 (+), 704.4 (−) |
| DON | 335.0 (+) | 297.0 (+), 340.9 (−) |
| ZEN | 317.0 (−) | 357.0 (+), 319.0 (+) |
| T-2 toxin | 505.1 (+) | 484.2 (+), 589.0 (+) |
| Mycotoxins | LOD, mg/kg | Control (G1) | 15 mg/kg FBs Diet (G2) | 30 mg/kg FBs Diet (G3) |
|---|---|---|---|---|
| FB1 | 0.031 | nd | 15.4 * | 29.75 * |
| FB2 | 0.051 | nd | ||
| FB3 | - | nd | ||
| DON | 0.053 | nd | nd | nd |
| ZEN | 0.005 | nd | nd | nd |
| T-2 toxin | 0.011 | nd | nd | nd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeebone, Y.Y.; Bóta, B.; Halas, V.; Libisch, B.; Olasz, F.; Papp, P.; Keresztény, T.; Gerőcs, A.; Ali, O.; Kovács, M.; et al. Gut-Faecal Microbial and Health-Marker Response to Dietary Fumonisins in Weaned Pigs. Toxins 2023, 15, 328. https://doi.org/10.3390/toxins15050328
Zeebone YY, Bóta B, Halas V, Libisch B, Olasz F, Papp P, Keresztény T, Gerőcs A, Ali O, Kovács M, et al. Gut-Faecal Microbial and Health-Marker Response to Dietary Fumonisins in Weaned Pigs. Toxins. 2023; 15(5):328. https://doi.org/10.3390/toxins15050328
Chicago/Turabian StyleZeebone, Yarsmin Yunus, Brigitta Bóta, Veronika Halas, Balázs Libisch, Ferenc Olasz, Péter Papp, Tibor Keresztény, Annamária Gerőcs, Omeralfaroug Ali, Melinda Kovács, and et al. 2023. "Gut-Faecal Microbial and Health-Marker Response to Dietary Fumonisins in Weaned Pigs" Toxins 15, no. 5: 328. https://doi.org/10.3390/toxins15050328