Anxiolytic, Analgesic and Anti-Inflammatory Effects of Peptides Hmg 1b-2 and Hmg 1b-4 from the Sea Anemone Heteractis magnifica

 , , ,
, , ,  and
and 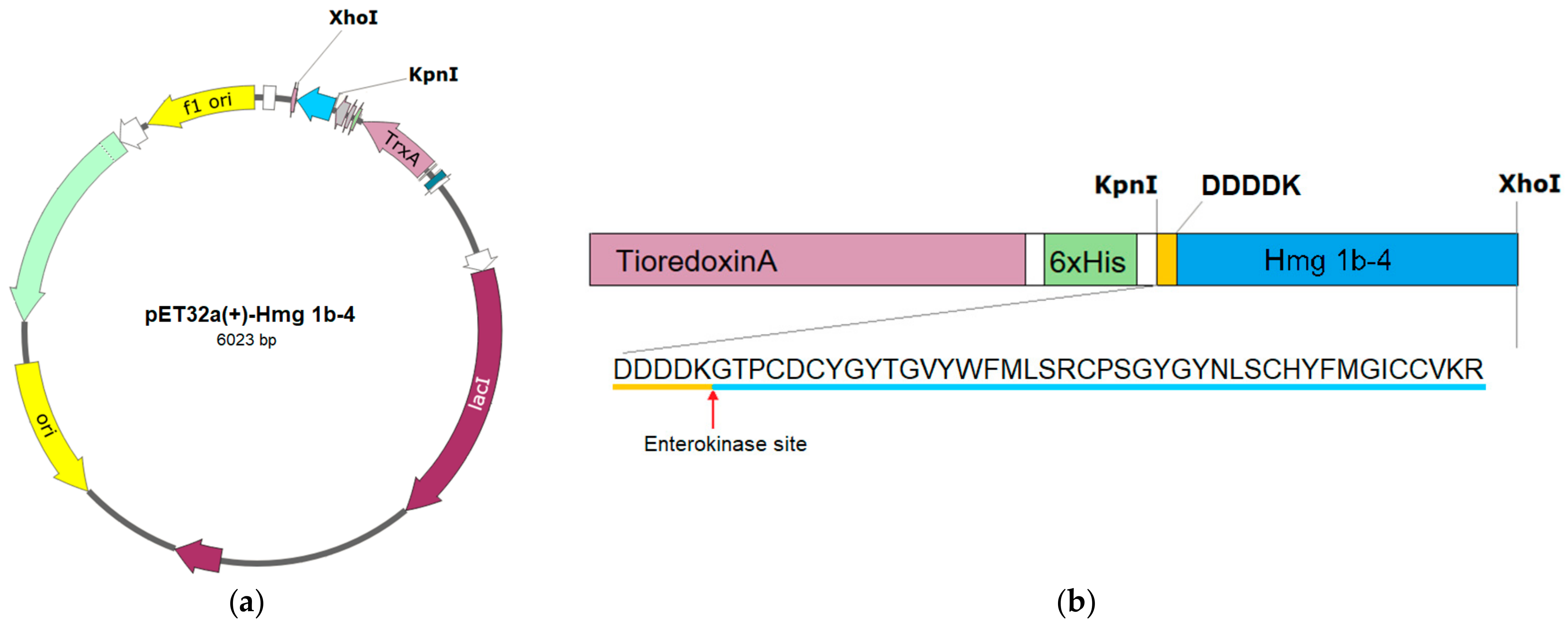{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
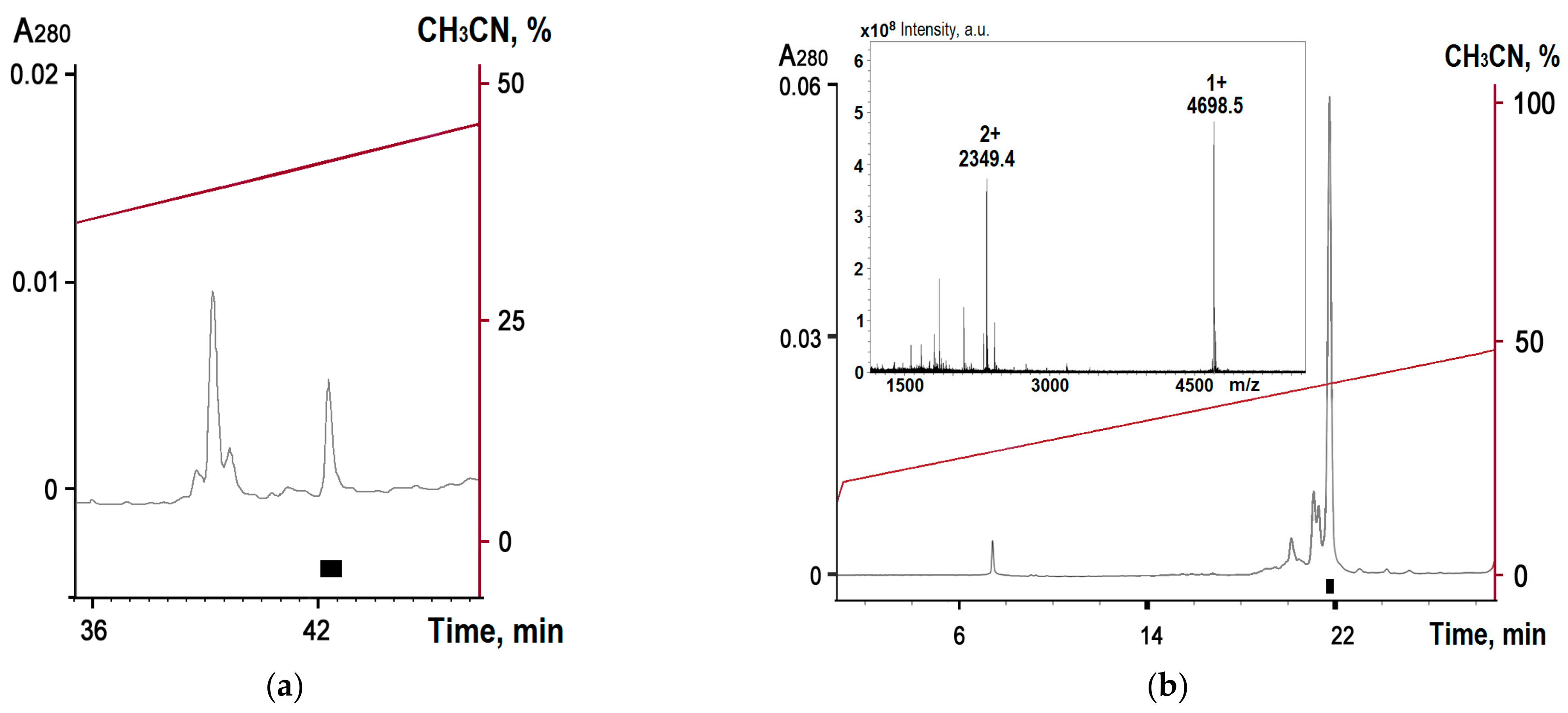
2.1. Obtaining Recombinant Hmg 1b-4
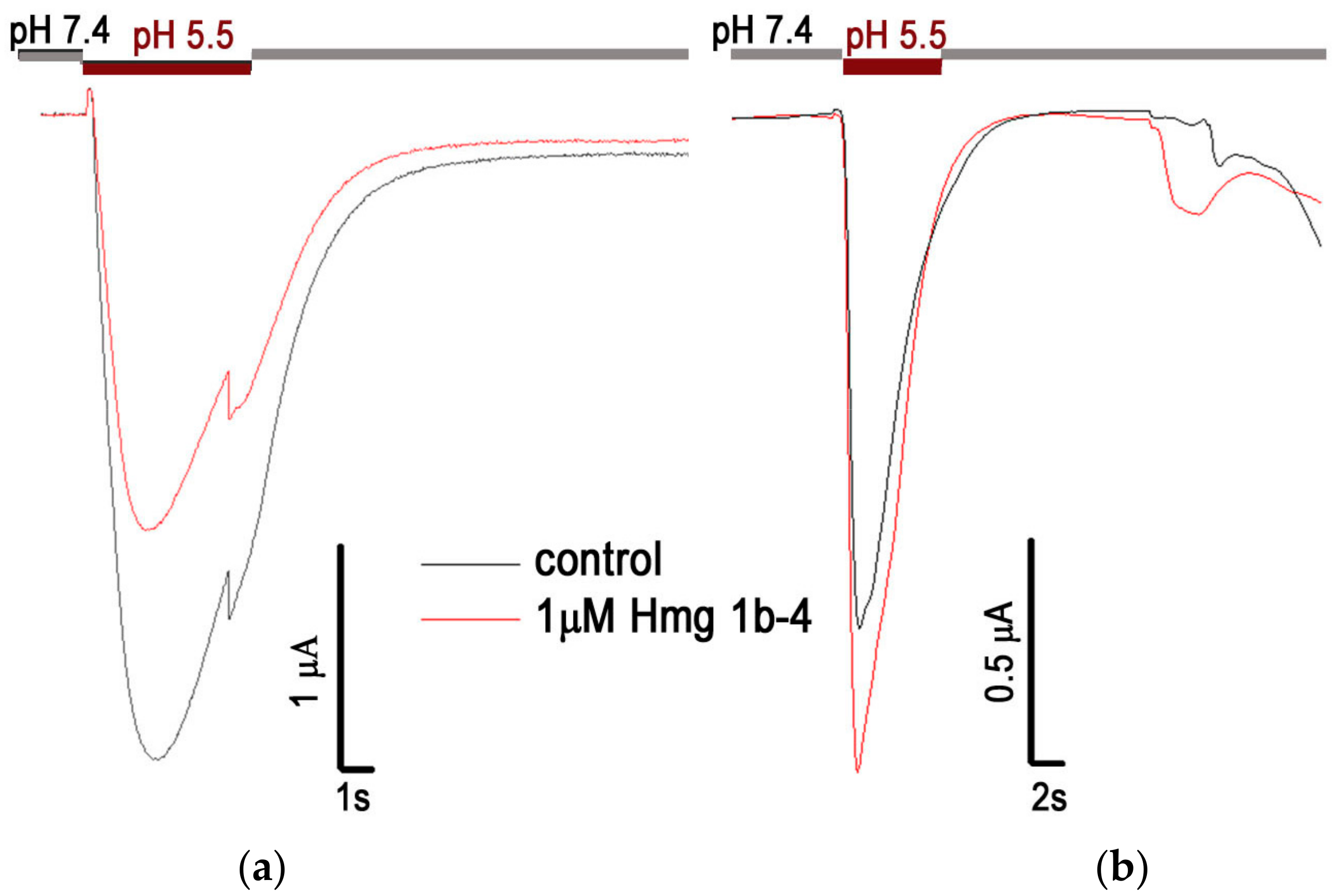
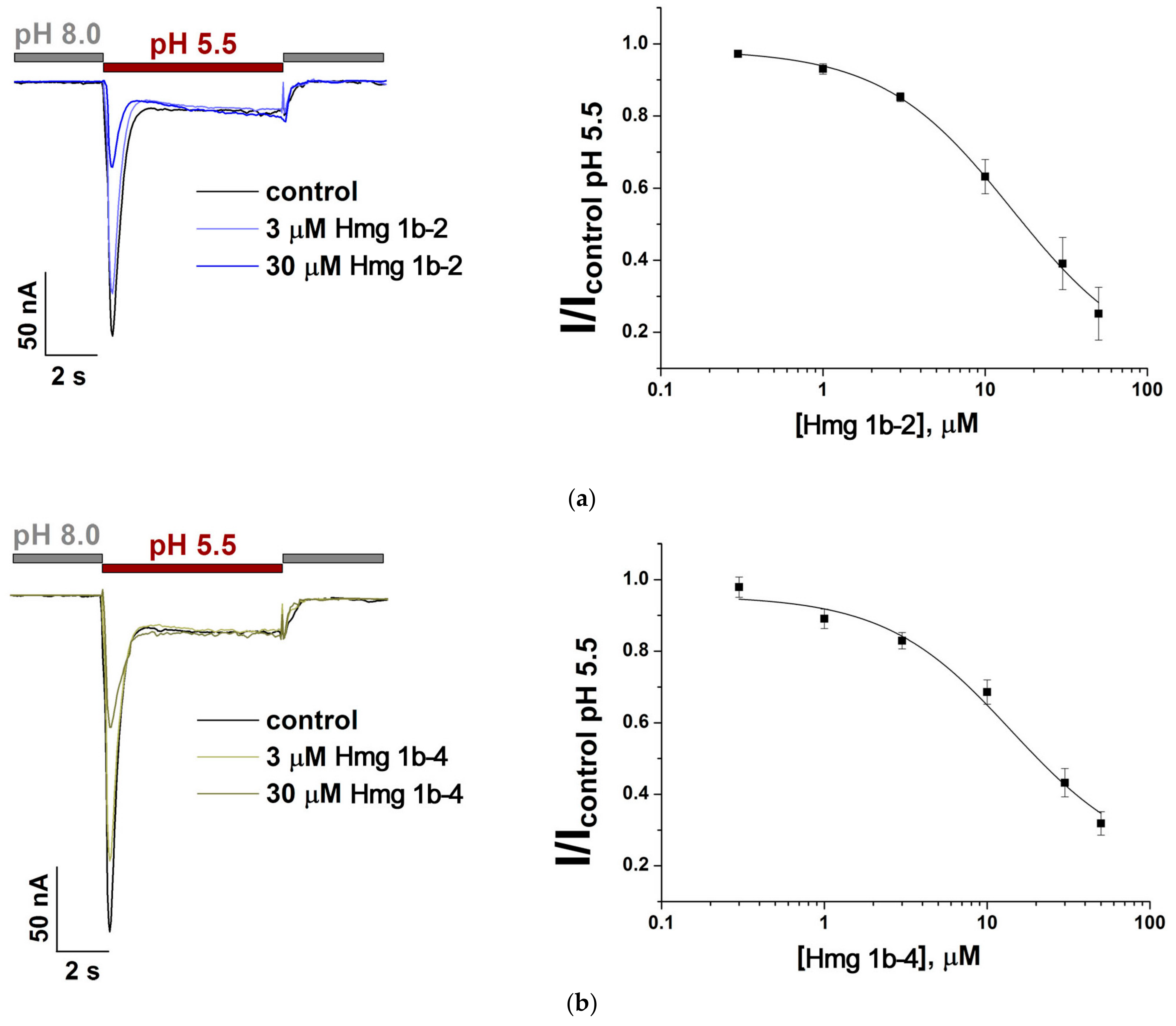
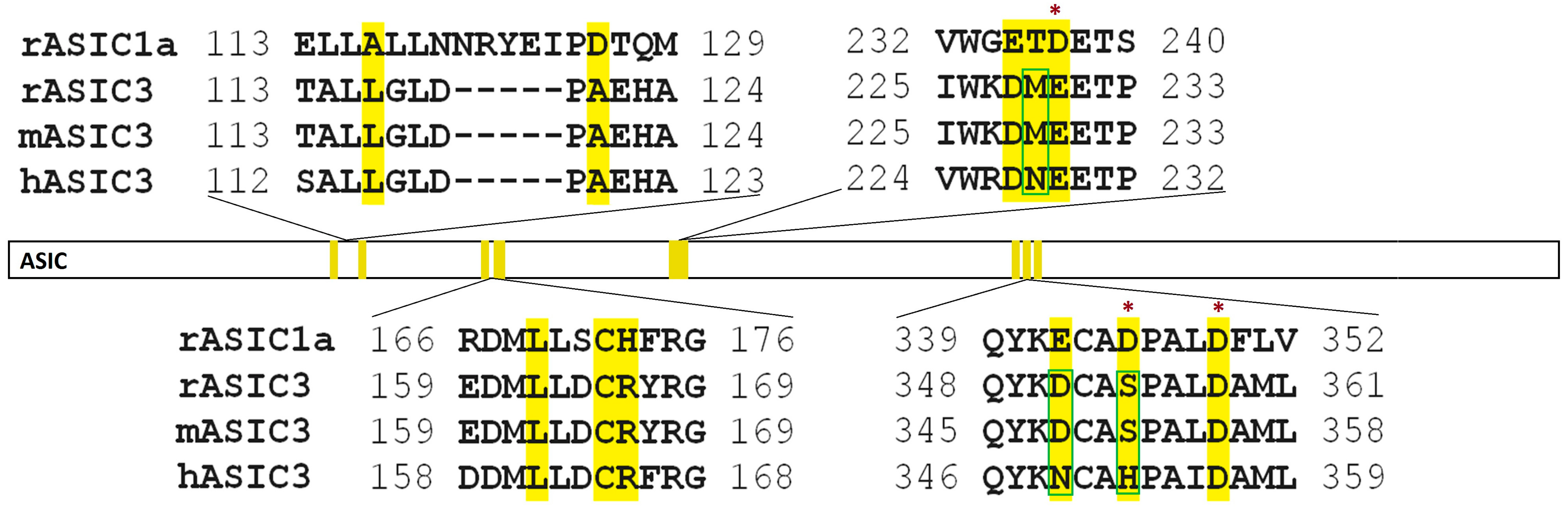
2.2. Electrophysiological Effects of Peptides on ASIC1a and ASIC3 Channels
2.3. Animal Experiments
2.3.1. Acute Toxicity
2.3.2. Behavioral Tests
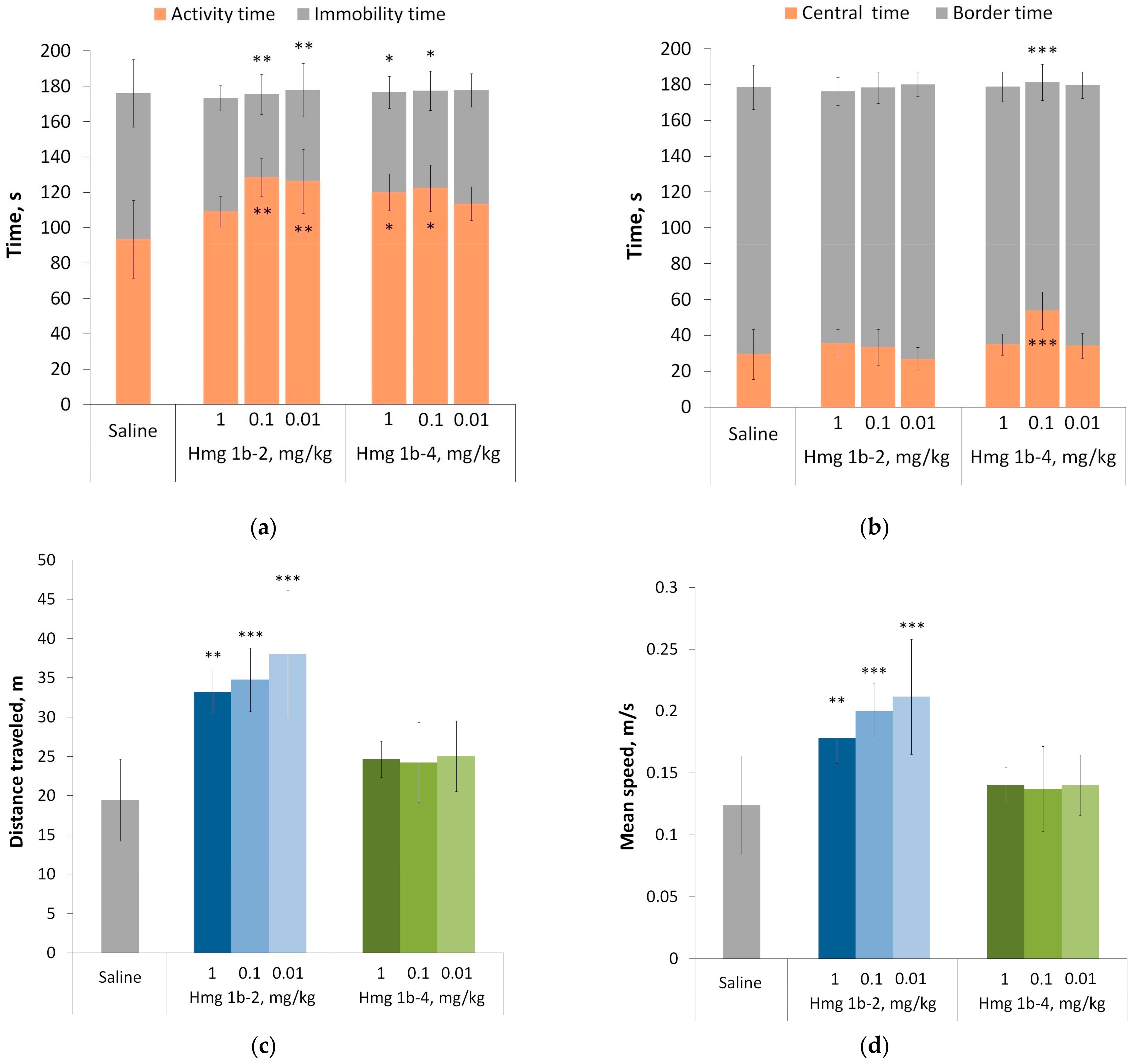
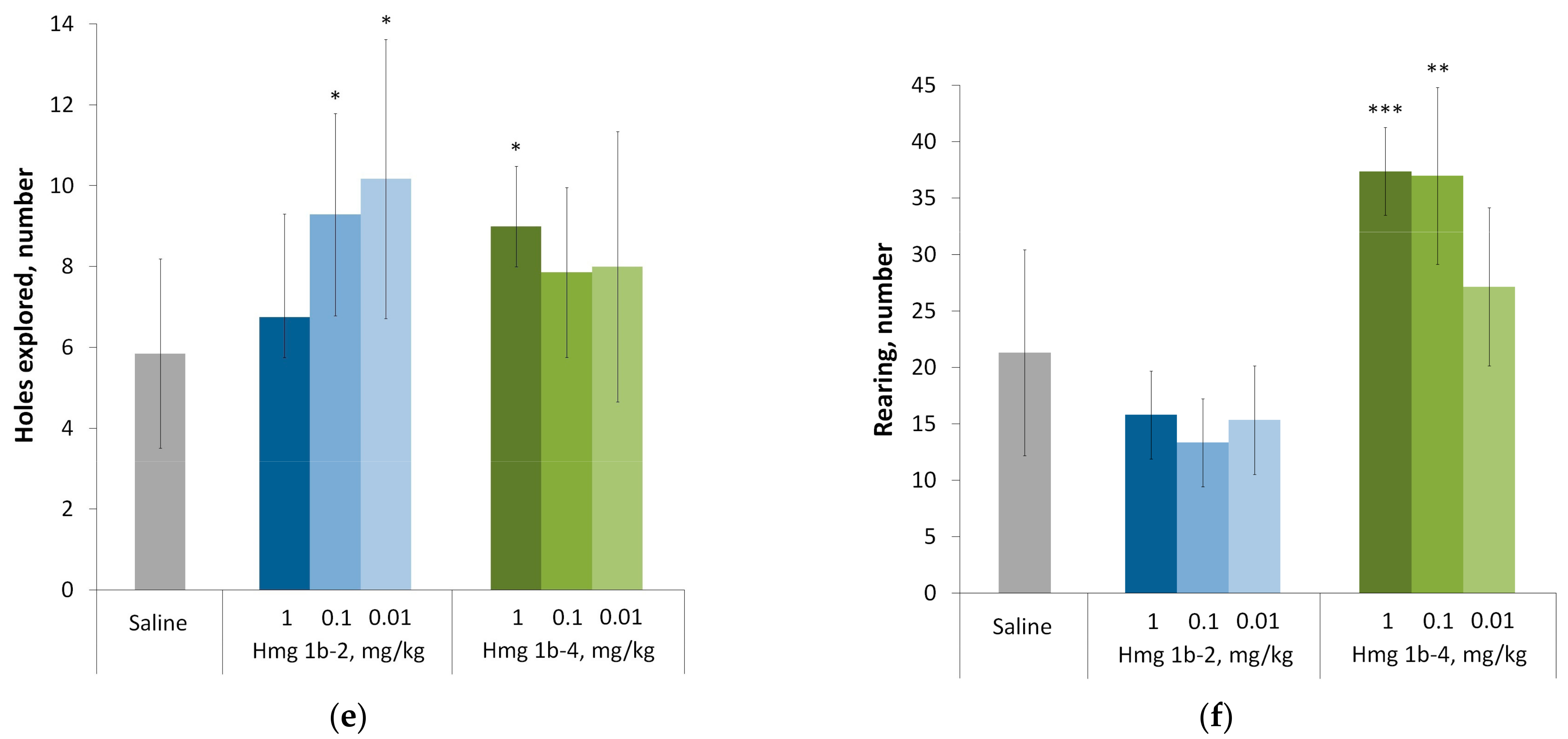
Open Field Test
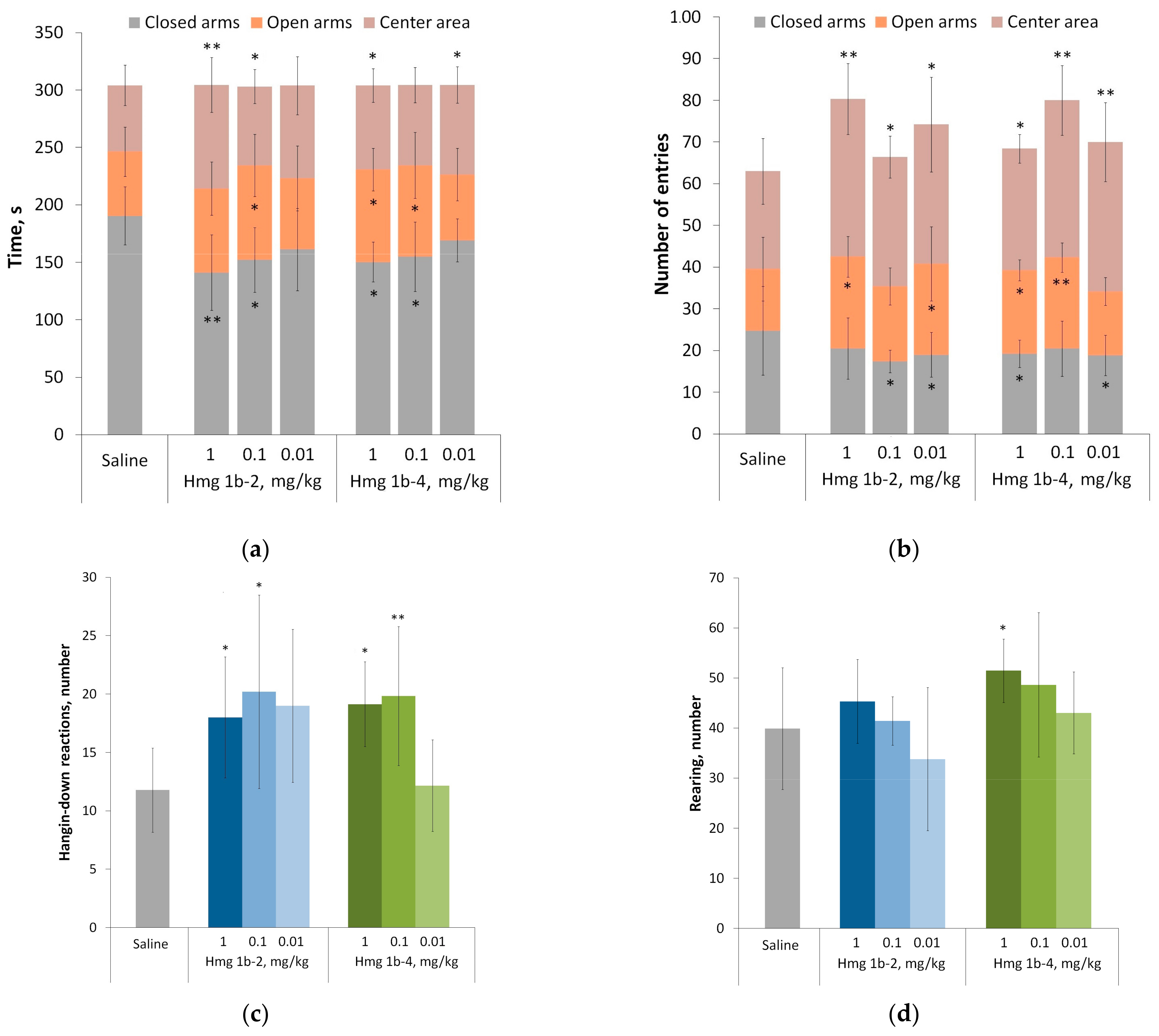
Elevated Plus Maze Test
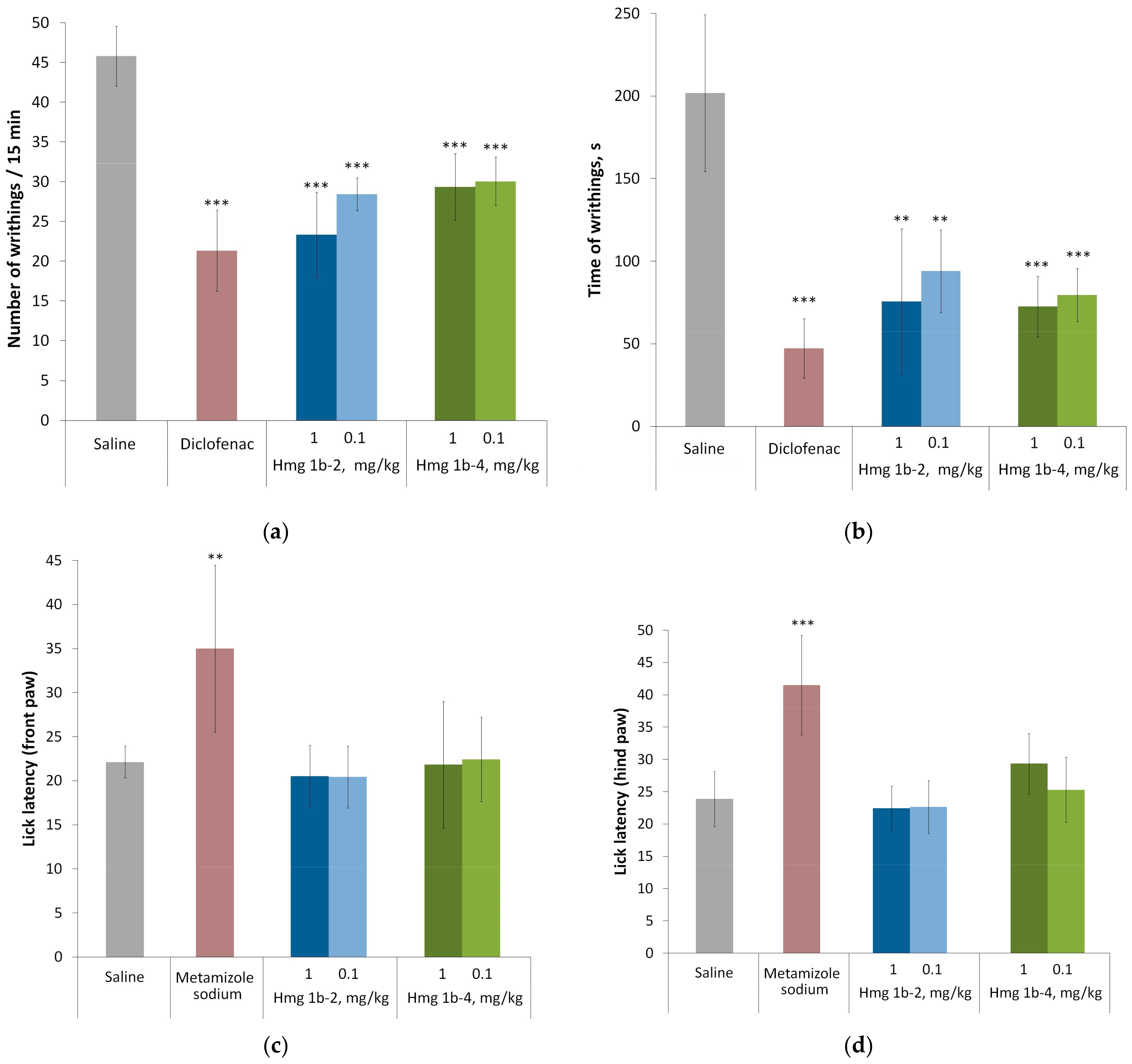
2.3.3. Analgesic Activity
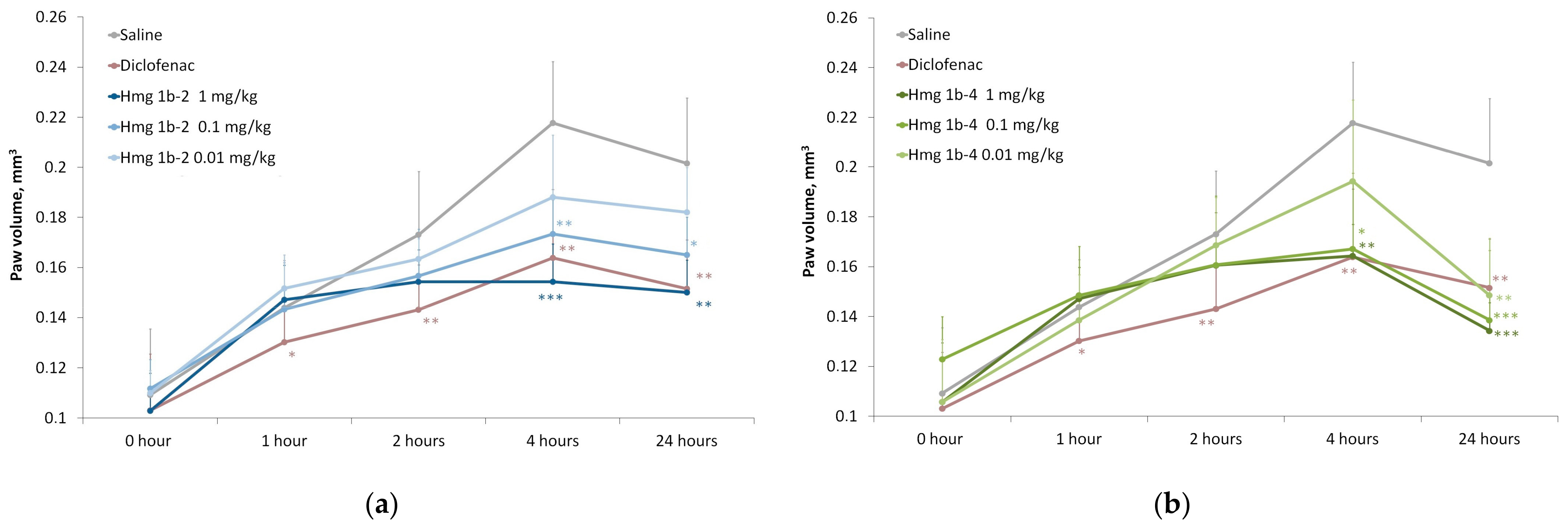
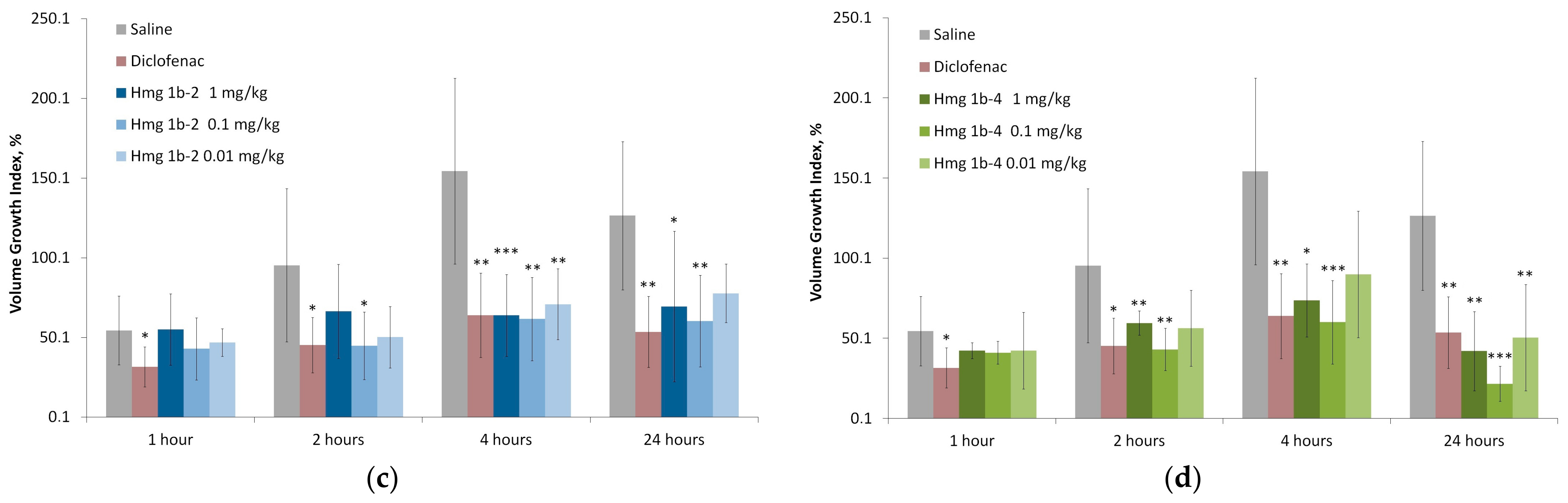
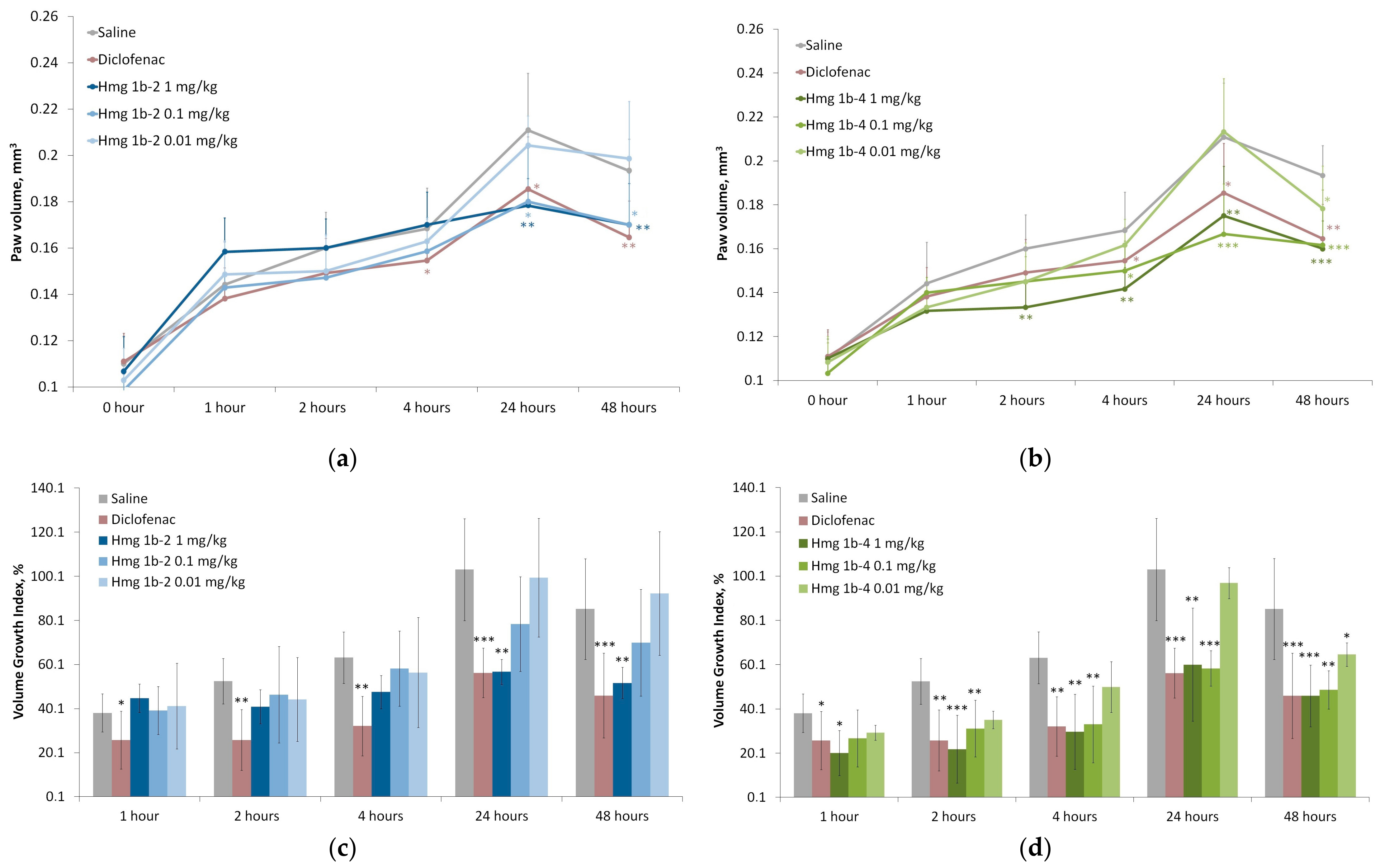
2.3.4. Anti-Inflammatory Activity
Carrageenan-Induced Paw Edema
Complete Freund’s Adjuvant-Induced Acute Inflammation
3. Discussion
4. Materials and Methods
4.1. Obtaining Peptides Hmg 1b-2 and Hmg1b-4
4.2. Electrophysiology
4.3. Animal Studies
4.3.1. Acute Toxicity
4.3.2. Open Field Test
4.3.3. Elevated Plus Maze Test
4.3.4. Acetic-Acid-Induced Writhings
4.3.5. Hot Plate Test
4.3.6. Carrageenan-Induced Paw Edema
4.3.7. Complete Freund’s-Adjuvant-Induced Acute Inflammation
4.4. Statistic Calculation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Panigrahy, D.; Gilligan, M.M.; Serhan, C.N.; Kashfi, K. Resolution of inflammation: An organizing principle in biology and medicine. Pharmacol. Ther. 2021, 227, 107879. [Google Scholar] [CrossRef] [PubMed]
- Marchand, F.; Perretti, M.; McMahon, S.B. Role of the immune system in chronic pain. Nat. Rev. Neurosci. 2005, 6, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Linley, J.E.; Rose, K.; Ooi, L.; Gamper, N. Understanding inflammatory pain: Ion channels contributing to acute and chronic nociception. Pflugers Arch. Eur. J. Physiol. 2010, 459, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, M.; Wallace, H. Ion channels in inflammation. Pflugers Arch. Eur. J. Physiol. 2011, 461, 401–421. [Google Scholar] [CrossRef]
- Babini, E.; Paukert, M.; Geisler, H.-S.; Gründer, S. Alternative splicing and interaction with di- and polyvalent cations control the dynamic range of Acid-sensing Ion Channel 1 (ASIC1). J. Biol. Chem. 2002, 277, 41597–41603. [Google Scholar] [CrossRef]
- Duan, B.; Wang, Y.-Z.; Yang, T.; Chu, X.-P.; Yu, Y.; Huang, Y.; Cao, H.; Hansen, J.; Simon, R.P.; Zhu, M.X.; et al. Extracellular spermine exacerbates ischemic neuronal injury through sensitization of ASIC1a channels to extracellular acidosis. J. Neurosci. 2011, 31, 2101–2112. [Google Scholar] [CrossRef]
- Tikhonov, D.B.; Magazanik, L.G.; Nagaeva, E.I. Ligands of Acid-Sensing Ion Channel 1a: Mechanisms of action and binding sites. Acta Nat. 2019, 11, 4–13. [Google Scholar] [CrossRef]
- Wiemuth, D.; Assmann, M.; Gründer, S. The bile acid-sensitive ion channel (BASIC), the ignored cousin of ASICs and ENaC. Channels 2014, 8, 29–34. [Google Scholar] [CrossRef]
- Pattison, L.A.; Callejo, G.; Smith, E.S.J. Evolution of acid nociception: Ion channels and receptors for detecting acid. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190291. [Google Scholar] [CrossRef]
- Ortega-Ramírez, A.; Vega, R.; Soto, E. Acid-Sensing Ion Channels as potential therapeutic targets in neurodegeneration and neuroinflammation. Mediat. Inflamm. 2017, 2017, 3728096. [Google Scholar] [CrossRef]
- Wu, J.; Xu, Y.; Jiang, Y.; Xu, J.; Hu, Y.; Zha, X. ASIC subunit ratio and differential surface trafficking in the brain. Mol. Brain 2016, 9, 4. [Google Scholar] [CrossRef]
- Soto, E.; Ortega-Ramírez, A.; Vega, R. Protons as messengers of intercellular communication in the nervous system. Front. Cell. Neurosci. 2018, 12, 342. [Google Scholar] [CrossRef] [PubMed]
- Santos, P.L.; Guimaraes, A.G.; Barreto, R.S.; Serafini, M.R.; Quintans, J.S.; Quintans-Junior, L.J. A Review of recent patents on the ASICs as a key drug target. Recent Pat. Biotechnol. 2015, 9, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Arun, T.; Tomassini, V.; Sbardella, E.; de Ruiter, M.B.; Matthews, L.; Leite, M.I.; Gelineau-Morel, R.; Cavey, A.; Vergo, S.; Craner, M.; et al. Targeting ASIC1 in primary progressive multiple sclerosis: Evidence of neuroprotection with amiloride. Brain 2013, 136, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Bohlen, C.J.; Chesler, A.T.; Sharif-Naeini, R.; Medzihradszky, K.F.; Zhou, S.; King, D.; Sánchez, E.E.; Burlingame, A.L.; Basbaum, A.I.; Julius, D. A heteromeric Texas coral snake toxin targets acid-sensing ion channels to produce pain. Nature 2011, 479, 410–414. [Google Scholar] [CrossRef]
- Reimers, C.; Lee, C.-H.; Kalbacher, H.; Tian, Y.; Hung, C.-H.; Schmidt, A.; Prokop, L.; Kauferstein, S.; Mebs, D.; Chen, C.-C.; et al. Identification of a cono-RFamide from the venom of Conus textile that targets ASIC3 and enhances muscle pain. Proc. Natl. Acad. Sci. USA 2017, 114, E3507–E3515. [Google Scholar] [CrossRef] [PubMed]
- Chassagnon, I.R.; McCarthy, C.A.; Chin, Y.K.-Y.; Pineda, S.S.; Keramidas, A.; Mobli, M.; Pham, V.; De Silva, T.M.; Lynch, J.W.; Widdop, R.E.; et al. Potent neuroprotection after stroke afforded by a double-knot spider-venom peptide that inhibits acid-sensing ion channel 1a. Proc. Natl. Acad. Sci. USA 2017, 114, 3750–3755. [Google Scholar] [CrossRef]
- Dibas, J.; Al-Saad, H.; Dibas, A. Basics on the use of acid-sensing ion channels’ inhibitors as therapeutics. Neural Regen. Res. 2019, 14, 395. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, C.A.; Rash, L.D.; Chassagnon, I.R.; King, G.F.; Widdop, R.E. PcTx1 affords neuroprotection in a conscious model of stroke in hypertensive rats via selective inhibition of ASIC1a. Neuropharmacology 2015, 99, 650–657. [Google Scholar] [CrossRef]
- Verkest, C.; Salinas, M.; Diochot, S.; Deval, E.; Lingueglia, E.; Baron, A. Mechanisms of action of the peptide toxins targeting human and rodent acid-sensing ion channels and relevance to their in vivo analgesic effects. Toxins 2022, 14, 709. [Google Scholar] [CrossRef]
- Escoubas, P.; De Weille, J.R.; Lecoq, A.; Diochot, S.; Waldmann, R.; Champigny, G.; Moinier, D.; Ménez, A.; Lazdunski, M. Isolation of a tarantula toxin specific for a class of proton-gated Na+ channels. J. Biol. Chem. 2000, 275, 25116–25121. [Google Scholar] [CrossRef]
- Liu, Y.; Hagan, R.; Schoellerman, J. Dual actions of Psalmotoxin at ASIC1a and ASIC2a heteromeric channels (ASIC1a/2a). Sci. Rep. 2018, 8, 7179. [Google Scholar] [CrossRef] [PubMed]
- Mazzuca, M.; Heurteaux, C.; Alloui, A.; Diochot, S.; Baron, A.; Voilley, N.; Blondeau, N.; Escoubas, P.; Gélot, A.; Cupo, A.; et al. A tarantula peptide against pain via ASIC1a channels and opioid mechanisms. Nat. Neurosci. 2007, 10, 943–945. [Google Scholar] [CrossRef] [PubMed]
- Diochot, S.; Baron, A.; Salinas, M.; Douguet, D.; Scarzello, S.; Dabert-Gay, A.-S.; Debayle, D.; Friend, V.; Alloui, A.; Lazdunski, M.; et al. Black mamba venom peptides target acid-sensing ion channels to abolish pain. Nature 2012, 490, 552–555. [Google Scholar] [CrossRef]
- Diochot, S.; Alloui, A.; Rodrigues, P.; Dauvois, M.; Friend, V.; Aissouni, Y.; Eschalier, A.; Lingueglia, E.; Baron, A. Analgesic effects of mambalgin peptide inhibitors of acid-sensing ion channels in inflammatory and neuropathic pain. Pain 2016, 157, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Frazão, B.; Vasconcelos, V.; Antunes, A. Sea Anemone (Cnidaria, Anthozoa, Actiniaria) Toxins: An Overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Menezes, C.; Thakur, N.L. Sea anemone venom: Ecological interactions and bioactive potential. Toxicon 2022, 208, 31–46. [Google Scholar] [CrossRef]
- Fu, J.; Liao, Y.; Jin, A.-H.; Gao, B. Discovery of novel peptide neurotoxins from sea anemone species. Front. Biosci. 2021, 26, 1256. [Google Scholar] [CrossRef]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea anemone toxins: A structural overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef]
- Ramírez-Carreto, S.; Miranda-Zaragoza, B.; Rodríguez-Almazán, C. Actinoporins: From the structure and function to the generation of biotechnological and therapeutic tools. Biomolecules 2020, 10, 539. [Google Scholar] [CrossRef]
- Moran, Y.; Gordon, D.; Gurevitz, M. Sea anemone toxins affecting voltage-gated sodium channels—Molecular and evolutionary features. Toxicon 2009, 54, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Osmakov, D.I.; Kozlov, S.A.; Andreev, Y.A.; Koshelev, S.G.; Sanamyan, N.P.; Sanamyan, K.E.; Dyachenko, I.A.; Bondarenko, D.A.; Murashev, A.N.; Mineev, K.S.; et al. Sea anemone peptide with uncommon β-hairpin structure inhibits acid-sensing ion channel 3 (ASIC3) and reveals analgesic activity. J. Biol. Chem. 2013, 288, 23116–23127. [Google Scholar] [CrossRef] [PubMed]
- Osmakov, D.I.; Koshelev, S.G.; Andreev, Y.A.; Dyachenko, I.A.; Bondarenko, D.A.; Murashev, A.N.; Grishin, E.V.; Kozlov, S.A. Conversed mutagenesis of an inactive peptide to ASIC3 inhibitor for active sites determination. Toxicon 2016, 116, 11–16. [Google Scholar] [CrossRef]
- Andreev, Y.; Osmakov, D.; Koshelev, S.; Maleeva, E.; Logashina, Y.; Palikov, V.; Palikova, Y.; Dyachenko, I.; Kozlov, S. Analgesic activity of acid-sensing ion channel 3 (ASIC3) Inhibitors: Sea anemones peptides Ugr9-1 and APETx2 versus low molecular weight compounds. Mar. Drugs 2018, 16, 500. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, J.; Spencer, R.H.; Garsky, V.M.; Liang, A.; Leitl, M.D.; Cato, M.J.; Cook, S.P.; Kane, S.; Urban, M.O. Reversal of acid-induced and inflammatory pain by the selective ASIC3 inhibitor, APETx2. Br. J. Pharmacol. 2010, 161, 950–960. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.P.; Saez, N.J.; Cristofori-Armstrong, B.; Anangi, R.; King, G.F.; Smith, M.T.; Rash, L.D. Inhibition of acid-sensing ion channels by diminazene and APETx2 evoke partial and highly variable antihyperalgesia in a rat model of inflammatory pain. Br. J. Pharmacol. 2018, 175, 2204–2218. [Google Scholar] [CrossRef] [PubMed]
- Deval, E.; Noel, J.; Gasull, X.; Delaunay, A.; Alloui, A.; Friend, V.; Eschalier, A.; Lazdunski, M.; Lingueglia, E. Acid-sensing ion channels in postoperative pain. J. Neurosci. 2011, 31, 6059–6066. [Google Scholar] [CrossRef] [PubMed]
- Karsan, N.; Gonzales, E.B.; Dussor, G. Targeted acid-sensing ion channel therapies for migraine. Neurotherapeutics 2018, 15, 402–414. [Google Scholar] [CrossRef]
- Kalina, R.S.; Kasheverov, I.E.; Koshelev, S.G.; Sintsova, O.V.; Peigneur, S.; Pinheiro-Junior, E.L.; Popov, R.S.; Chausova, V.E.; Monastyrnaya, M.M.; Dmitrenok, P.S.; et al. Nicotinic acetylcholine receptors are novel targets of APETx-like Toxins from the sea anemone Heteractis magnifica. Toxins 2022, 14, 697. [Google Scholar] [CrossRef]
- Kalina, R.; Gladkikh, I.; Dmitrenok, P.; Chernikov, O.; Koshelev, S.; Kvetkina, A.; Kozlov, S.; Kozlovskaya, E.; Monastyrnaya, M. New APETx-like peptides from sea anemone Heteractis crispa modulate ASIC1a channels. Peptides 2018, 104, 41–49. [Google Scholar] [CrossRef]
- Kalina, R.S.; Koshelev, S.G.; Zelepuga, E.A.; Kim, N.Y.; Kozlov, S.A.; Kozlovskaya, E.P.; Monastyrnaya, M.M.; Gladkikh, I.N. APETx-like peptides from the sea anemone Heteractis crispa, diverse in their effect on ASIC1a and ASIC3 ion channels. Toxins 2020, 12, 266. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro-Junior, E.L.; Kalina, R.; Gladkikh, I.; Leychenko, E.; Tytgat, J.; Peigneur, S. A Tale of Toxin Promiscuity: The versatile pharmacological effects of Hcr 1b-2 sea anemone peptide on voltage-gated ion channels. Mar. Drugs 2022, 20, 147. [Google Scholar] [CrossRef] [PubMed]
- Logashina, Y.A.; Korolkova, Y.V.; Maleeva, E.E.; Osmakov, D.I.; Kozlov, S.A.; Andreev, Y.A. Refolding of disulfide containing peptides in fusion with thioredoxin. Mendeleev Commun. 2020, 30, 214–216. [Google Scholar] [CrossRef]
- Osmakov, D.I.; Korolkova, Y.V.; Lubova, K.I.; Maleeva, E.E.; Andreev, Y.A.; Kozlov, S.A. The role of the C-terminal intracellular domain in acid-sensing ion channel 3 functioning. J. Evol. Biochem. Physiol. 2021, 57, 413–423. [Google Scholar] [CrossRef]
- Voilley, N.; de Weille, J.; Mamet, J.; Lazdunski, M. Nonsteroid anti-inflammatory drugs inhibit both the activity and the inflammation-induced expression of acid-sensing ion channels in nociceptors. J. Neurosci. 2001, 21, 8026–8033. [Google Scholar] [CrossRef]
- Jasiecka, A.; Maślanka, T.; Jaroszewski, J.J. Pharmacological characteristics of metamizole. Pol. J. Vet. Sci. 2014, 17, 207–214. [Google Scholar] [CrossRef]
- Yen, Y.-T.; Tu, P.-H.; Chen, C.-J.; Lin, Y.-W.; Hsieh, S.-T.; Chen, C.-C. Role of acid-sensing ion channel 3 in sub-acute-phase inflammation. Mol. Pain 2009, 5, 1. [Google Scholar] [CrossRef]
- Klipp, R.C.; Cullinan, M.M.; Bankston, J.R. Insights into the molecular mechanisms underlying the inhibition of acid-sensing ion channel 3 gating by stomatin. J. Gen. Physiol. 2020, 152, e201912471. [Google Scholar] [CrossRef]
- Cristofori-armstrong, B.; Saez, N.J.; Chassagnon, I.R.; King, G.F.; Rash, L.D. The modulation of acid-sensing ion channel 1 by PcTx1 is pH-, subtype- and species-dependent: Importance of interactions at the channel subunit interface and potential for engineering selective analogues. Biochem. Pharmacol. 2019, 163, 381–390. [Google Scholar] [CrossRef]
- Wulff, H.; Castle, N.A.; Pardo, L.A. Voltage-gated potassium channels as therapeutic targets. Nat. Rev. Drug Discov. 2009, 8, 982–1001. [Google Scholar] [CrossRef]
- Coryell, M.W.; Wunsch, A.M.; Haenfler, J.M.; Allen, J.E.; Schnizler, M.; Ziemann, A.E.; Cook, M.N.; Dunning, J.P.; Price, M.P.; Rainier, J.D.; et al. Acid-Sensing Ion Channel-1a in the amygdala, a novel therapeutic target in depression-related behavior. J. Neurosci. 2009, 29, 5381–5388. [Google Scholar] [CrossRef] [PubMed]
- Taugher, R.J.; Lu, Y.; Fan, R.; Ghobbeh, A.; Kreple, C.J.; Faraci, F.M.; Wemmie, J.A. ASIC1A in neurons is critical for fear-related behaviors. Genes Brain Behav. 2017, 16, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Pidoplichko, V.I.; Aroniadou-Anderjaska, V.; Prager, E.M.; Figueiredo, T.H.; Almeida-Suhett, C.P.; Miller, S.L.; Braga, M.F.M. ASIC1a activation enhances inhibition in the basolateral amygdala and reduces anxiety. J. Neurosci. 2014, 34, 3130–3141. [Google Scholar] [CrossRef] [PubMed]
- Storozhuk, M.; Cherninskyi, A.; Maximyuk, O.; Isaev, D.; Krishtal, O. Acid-Sensing Ion Channels: Focus on physiological and some pathological roles in the brain. Curr. Neuropharmacol. 2021, 19, 1570–1589. [Google Scholar] [CrossRef] [PubMed]
- Ritzel, R.M.; He, J.; Li, Y.; Cao, T.; Khan, N.; Shim, B.; Sabirzhanov, B.; Aubrecht, T.; Stoica, B.A.; Faden, A.I.; et al. Proton extrusion during oxidative burst in microglia exacerbates pathological acidosis following traumatic brain injury. Glia 2021, 69, 746–764. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-H.; Jones, S.; Talman, W.T. Cellular localization of acid-sensing ion channel 1 in rat nucleus tractus solitarii. Cell. Mol. Neurobiol. 2018, 38, 219–232. [Google Scholar] [CrossRef]
- Stein, C.; Gaveriaux-Ruff, C. Opioids and pain. In The Oxford Handbook of the Neurobiology of Pain; Oxford University Press: Oxford, UK, 2018; pp. 728–769. [Google Scholar]
- Chesler, M. Regulation and modulation of pH in the brain. Physiol. Rev. 2003, 83, 1183–1221. [Google Scholar] [CrossRef]
- Pakulska, W.; Czarnecka, E. Influence of mianserin on the antinociceptive effect of morphine, metamizol and indomethacin in mice. Pharmacol. Res. 2002, 46, 415–423. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gladkikh, I.N.; Klimovich, A.A.; Kalina, R.S.; Kozhevnikova, Y.V.; Khasanov, T.A.; Osmakov, D.I.; Koshelev, S.G.; Monastyrnaya, M.M.; Andreev, Y.A.; Leychenko, E.V.; et al. Anxiolytic, Analgesic and Anti-Inflammatory Effects of Peptides Hmg 1b-2 and Hmg 1b-4 from the Sea Anemone Heteractis magnifica. Toxins 2023, 15, 341. https://doi.org/10.3390/toxins15050341
Gladkikh IN, Klimovich AA, Kalina RS, Kozhevnikova YV, Khasanov TA, Osmakov DI, Koshelev SG, Monastyrnaya MM, Andreev YA, Leychenko EV, et al. Anxiolytic, Analgesic and Anti-Inflammatory Effects of Peptides Hmg 1b-2 and Hmg 1b-4 from the Sea Anemone Heteractis magnifica. Toxins. 2023; 15(5):341. https://doi.org/10.3390/toxins15050341
Chicago/Turabian StyleGladkikh, Irina N., Anna A. Klimovich, Rimma S. Kalina, Yulia V. Kozhevnikova, Timur A. Khasanov, Dmitry I. Osmakov, Sergey G. Koshelev, Margarita M. Monastyrnaya, Yaroslav A. Andreev, Elena V. Leychenko, and et al. 2023. "Anxiolytic, Analgesic and Anti-Inflammatory Effects of Peptides Hmg 1b-2 and Hmg 1b-4 from the Sea Anemone Heteractis magnifica" Toxins 15, no. 5: 341. https://doi.org/10.3390/toxins15050341
APA StyleGladkikh, I. N., Klimovich, A. A., Kalina, R. S., Kozhevnikova, Y. V., Khasanov, T. A., Osmakov, D. I., Koshelev, S. G., Monastyrnaya, M. M., Andreev, Y. A., Leychenko, E. V., & Kozlov, S. A. (2023). Anxiolytic, Analgesic and Anti-Inflammatory Effects of Peptides Hmg 1b-2 and Hmg 1b-4 from the Sea Anemone Heteractis magnifica. Toxins, 15(5), 341. https://doi.org/10.3390/toxins15050341