Rattlesnake Crotalphine Analgesic Active on Tetrodotoxin-Sensitive Na+ Current in Mouse Dorsal Root Ganglion Neurons


 ,
,  and
and 
Abstract
:1. Introduction
2. Results
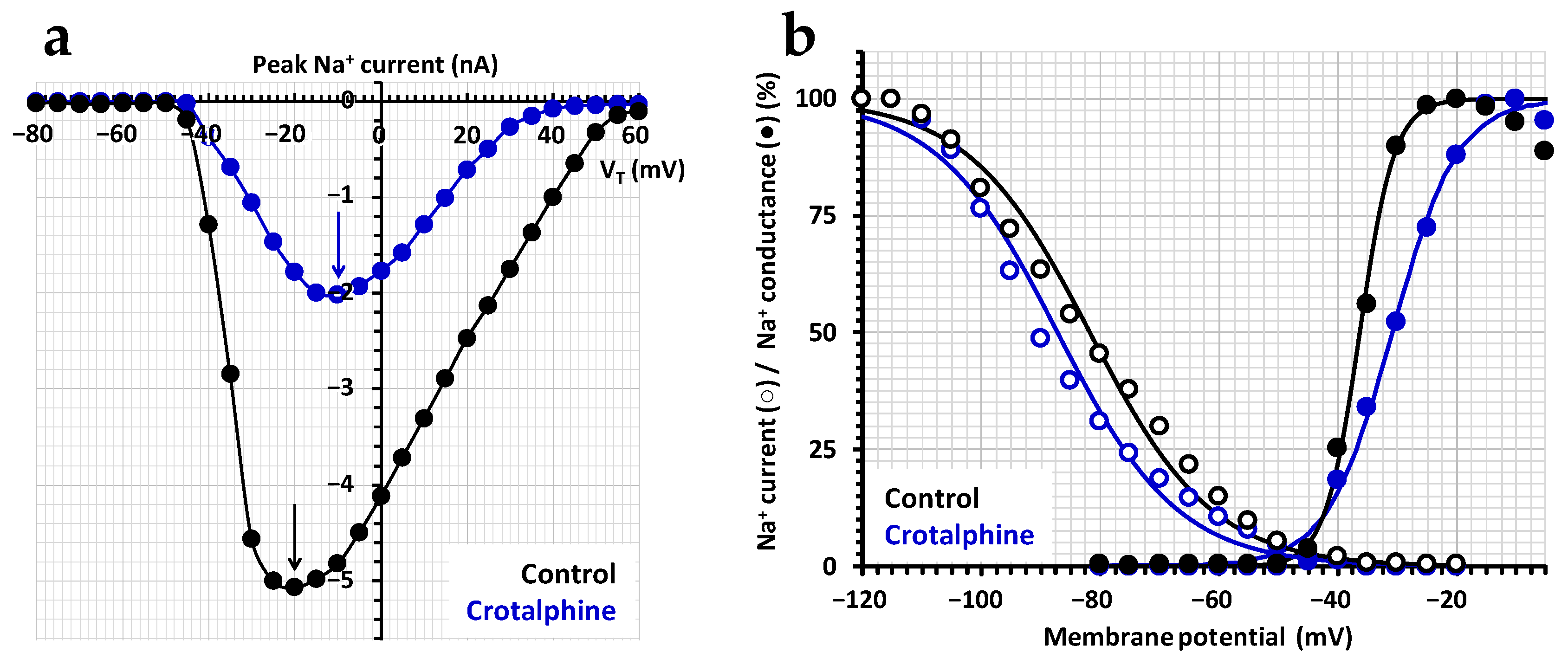
2.1. Effect of Crotalphine on the TTX-Sensitive Na+ Current
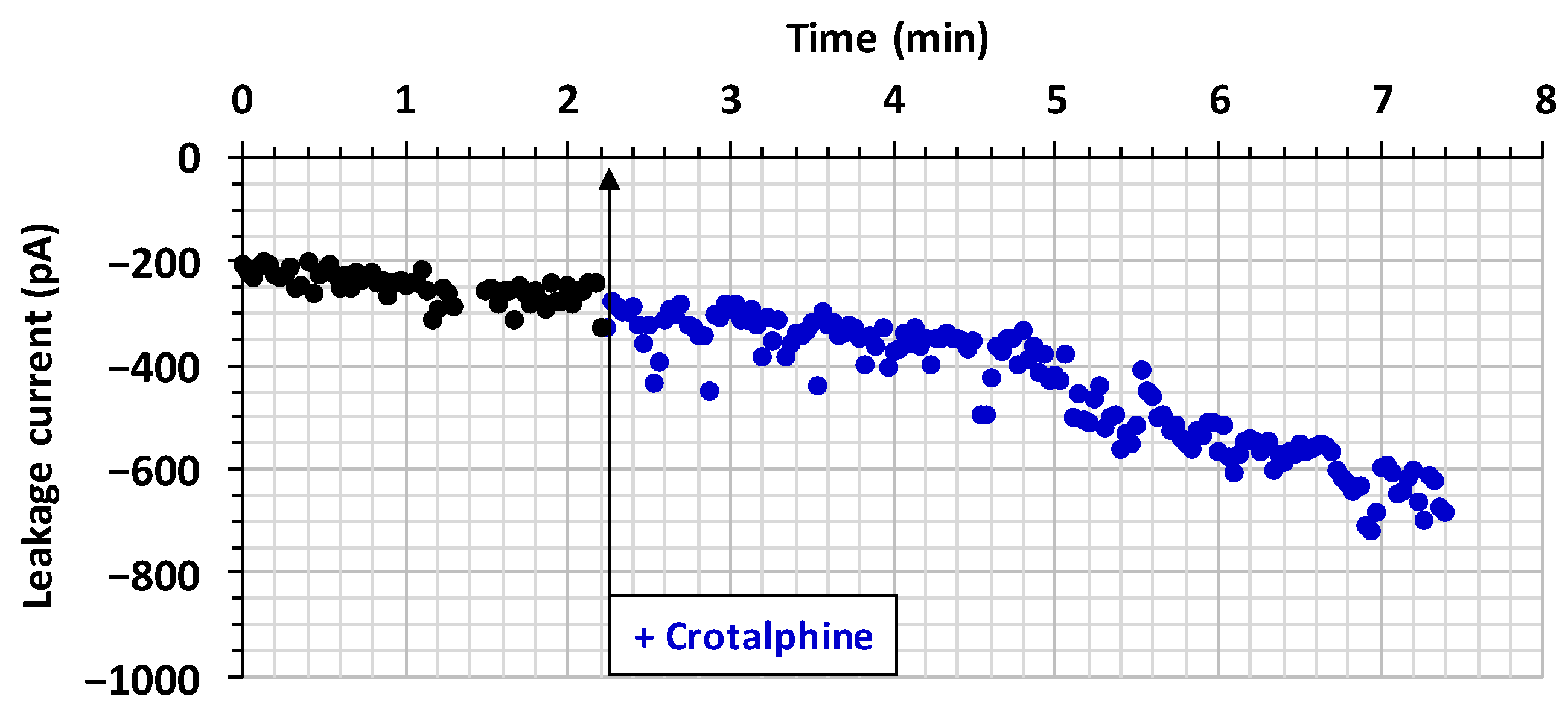
2.2. Effect of Crotalphine on Cell Viability
3. Discussion
3.1. Crotalphine-Induced Blocking of NaV1.7 Channel Subtype
3.2. Crotalphine-Induced Decrease in Cell Viability
4. Conclusions
5. Materials and Methods
5.1. Chemical Synthesis and Folding of Crotalphine
5.2. Toxins
5.3. Cultures of CHO Epithelial Cell Line and Primary Cultures of DRG Neurons
5.4. Electrophysiological Recordings
5.5. Cell Viability Assays
5.6. Data and Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frare, B.T.; Silva Resende, Y.K.; Dornelas, B.C.; Jorge, M.T.; Souza Ricarte, V.A.; Alves, L.M.; Izidoro, L.F.M. Clinical, laboratory, and therapeutic aspects of Crotalus durissus (South American rattlesnake) victims: A literature review. Biomed. Res. Int. 2019, 2019, 1345923. [Google Scholar] [CrossRef] [PubMed]
- Cañas, C.A. Biological and medical aspects related to South American rattlesnake Crotalus durissus (Linnaeus, 1758): A view from Colombia. Toxins 2022, 14, 875. [Google Scholar] [CrossRef] [PubMed]
- Macedo, J.M.; de Lima, A.M.; Kayano, A.M.; Souza, M.F.; da Silva Oliveira, I.; Gomez Garay, A.F.; Rocha, A.M.; Zuliani, J.P.; Soares, A.M. Literature review on Crotalus durissus terrificus toxins: From a perspective of structural biology and therapeutic applications. Curr. Protein Pept. Sci. 2023, 24, 536–550. [Google Scholar] [CrossRef] [PubMed]
- Laurenti, J.N. Specimen Medicum, Exhibens Synopsin Reptilium Emendatam cum Experimentis Circa Venena et Antidota Reptilium Aus-tracorum, Quod Authoritate et Consensu; Johann Thomas von Trattner: Vienna, Austria, 1768; 217p. [Google Scholar]
- Brazil, V. Do emprego da peçonha na terapêutica. Biol. Méd. São Paulo 1934, 1, 7–21. [Google Scholar]
- Giorgi, R.; Berardi, M.M.; Cury, Y. Analgesic effect evoked by low molecular weight substances extracted from Crotalus durissus terrificus venom. Toxicon 1993, 31, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Picolo, G.; Gutierrez, V.P.; Brigatte, P.; Zambelli, V.O.; Camargo, A.C.; Cury, Y. Crotalphine, a novel potent analgesic peptide from the venom of the South American rattlesnake Crotalus durissus terrificus. Peptides 2008, 29, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Giglio, J.R. Analytical studies on crotamine hydrochloride. Anal. Biochem. 1975, 69, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, V.O.; Fernandes, A.C.; Gutierrez, V.P.; Ferreira, J.C.; Parada, C.A.; Mochly-Rosen, D.; Cury, Y. Peripheral sensitization increases opioid receptor expression and activation by crotalphine in rats. PLoS ONE 2014, 9, e90576. [Google Scholar] [CrossRef]
- Gutierrez, V.P.; Konno, K.; Chacur, M.; Sampaio, S.C.; Picolo, G.; Brigatte, P.; Zambelli, V.O.; Cury, Y. Crotalphine induces potent antinociception in neuropathic pain by acting at peripheral opioid receptors. Eur. J. Pharmacol. 2008, 594, 84–92. [Google Scholar] [CrossRef]
- Gutierrez, V.P.; Zambelli, V.O.; Picolo, G.; Chacur, M.; Sampaio, S.C.; Brigatte, P.; Konno, K.; Cury, Y. The peripheral L-arginine-nitric oxide-cyclic GMP pathway and ATP-sensitive K⁺ channels are involved in the antinociceptive effect of crotalphine on neuropathic pain in rats. Behav. Pharmacol. 2012, 23, 14–24. [Google Scholar] [CrossRef]
- Brigatte, P.; Konno, K.; Gutierrez, V.P.; Sampaio, S.C.; Zambelli, V.O.; Picolo, G.; Curi, R.; Cury, Y. Peripheral kappa and delta opioid receptors are involved in the antinociceptive effect of crotalphine in a rat model of cancer pain. Pharmacol. Biochem. Behav. 2013, 109, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Giardini, A.C.; Evangelista, B.G.; Sant’Anna, M.B.; Martins, B.B.; Lancellotti, C.L.P.; Ciena, A.P.; Chacur, M.; Pagano, R.L.; Ribeiro, O.G.; Zambelli, V.O.; et al. Crotalphine attenuates pain and neuroinflammation induced by experimental autoimmune encephalomyelitis in mice. Toxins 2021, 13, 827. [Google Scholar] [CrossRef] [PubMed]
- Machado, F.C.; Zambelli, V.O.; Fernandes, A.C.; Heimann, A.S.; Cury, Y.; Picolo, G. Peripheral interactions between cannabinoid and opioid systems contribute to the antinociceptive effect of crotalphine. Br. J. Pharmacol. 2014, 171, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Hösch, N.G.; Martins, B.B.; Alcantara, Q.A.; Bufalo, M.C.; Neto, B.S.; Chudzinki-Tavassi, A.M.; Santa-Cecilia, F.V.; Cury, Y.; Zambelli, V.O. Wnt signaling is involved in crotalphine-induced analgesia in a rat model of neuropathic pain. Eur. J. Pharmacol. 2023, 959, 176058. [Google Scholar] [CrossRef] [PubMed]
- Bressan, E.; Touska, F.; Vetter, I.; Kistner, K.; Kichko, T.I.; Teixeira, N.B.; Picolo, G.; Cury, Y.; Lewis, R.J.; Fischer, M.J.M.; et al. Crotalphine desensitizes TRPA1 ion channels to alleviate inflammatory hyperalgesia. Pain 2016, 157, 2504–2516. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, T.C.; Benoit, E.; Partiseti, M.; Servent, D. The NaV1.7 channel subtype as an antinociceptive target for spider toxins in adult dorsal root ganglia neurons. Front. Pharmacol. 2018, 9, 1000. [Google Scholar] [CrossRef]
- Vetter, I.; Deuis, J.R.; Mueller, A.; Israel, M.R.; Starobova, H.; Zhang, A.; Rash, L.D.; Mobli, M. NaV1.7 as a pain target—From gene to pharmacology. Pharmacol. Ther. 2017, 172, 73–100. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Mis, M.A.; Estacion, M.; Dib-Hajj, S.D.; Waxman, S.G. NaV1.7 as a pharmacogenomic target for pain: Moving toward precision medicine. Trends Pharmacol. Sci. 2018, 39, 258–275. [Google Scholar] [CrossRef] [PubMed]
- de Lera Ruiz, M.; Kraus, R.L. Voltage-gated sodium channels: Structure, function, pharmacology, and clinical indications. J. Med. Chem. 2015, 58, 7093–7118. [Google Scholar] [CrossRef] [PubMed]
- Alexandrou, A.J.; Brown, A.R.; Chapman, M.L.; Estacion, M.; Turner, J.; Mis, M.A.; Wilbrey, A.; Payne, E.C.; Gutteridge, A.; Cox, P.J.; et al. Subtype-selective small molecule inhibitors reveal a fundamental role for NaV1.7 in nociceptor electrogenesis, axonal conduction and presynaptic release. PLoS ONE 2016, 11, e0152405. [Google Scholar] [CrossRef]
- Flinspach, M.; Xu, Q.; Piekarz, A.D.; Fellows, R.; Hagan, R.; Gibbs, A.; Liu, Y.; Neff, R.A.; Freedman, J.; Eckert, W.A.; et al. Insensitivity to pain induced by a potent selective closed-state NaV1.7 inhibitor. Sci. Rep. 2017, 7, 39662. [Google Scholar] [CrossRef] [PubMed]
- Schmalhofer, W.A.; Calhoun, J.; Burrows, R.; Bailey, T.; Kohler, M.G.; Weinglass, A.B.; Kaczorowski, G.J.; Garcia, M.L.; Koltzenburg, M.; Priest, B.T. ProTx-II, a selective inhibitor of NaV1.7 sodium channels, blocks action potential propagation in nociceptors. Mol. Pharmacol. 2008, 74, 1476–1484. [Google Scholar] [CrossRef]
- Montnach, J.; De Waard, S.; Nicolas, S.; Burel, S.; Osorio, N.; Zoukimian, C.; Mantegazza, M.; Boukaiba, R.; Béroud, R.; Partiseti, M.; et al. Fluorescent- and tagged-protoxin II peptides: Potent markers of the NaV1.7 channel pain target. Br. J. Pharmacol. 2021, 178, 2632–2650. [Google Scholar] [CrossRef]
- Deplazes, E.; Henriques, S.T.; Smith, J.J.; King, G.F.; Craik, D.J.; Mark, A.E.; Schroeder, C.I. Membrane-binding properties of gating modifier and pore-blocking toxins: Membrane interaction is not a prerequisite for modification of channel gating. Biochim. Biophys. Acta 2016, 1858, 872–882. [Google Scholar] [CrossRef]
- Hille, B. Ionic basis of resting and action potentials. In Handbook of Physiology; Kandel, E.R., Ed.; Sec. I “The Nervous System”, Vol. I “Cellular Biology of Neurons”; American Physiological Society: Bethesda, MD, USA, 1977; Part 1, Chapter 4; pp. 99–136. [Google Scholar] [CrossRef]
- Agwa, A.J.; Peigneur, S.; Chow, C.Y.; Lawrence, N.; Craik, D.J.; Tytgat, J.; King, G.F.; Henriques, S.T.; Schroeder, C.I. Gating modifier toxins isolated from spider venom: Modulation of voltage-gated sodium channels and the role of lipid membranes. J. Biol. Chem. 2018, 293, 9041–9052. [Google Scholar] [CrossRef]
- Matavel, A.C.; Ferreira-Alves, D.L.; Beirão, P.S.; Cruz, J.S. Tension generation and increase in voltage-activated Na+ current by crotamine. Eur. J. Pharmacol. 1998, 348, 167–173. [Google Scholar] [CrossRef]
- Nicastro, G.; Franzoni, L.; de Chiara, C.; Mancin, A.C.; Giglio, J.R.; Spisni, A. Solution structure of crotamine, a Na+ channel affecting toxin from Crotalus durissus terrificus venom. Eur. J. Biochem. 2003, 270, 1969–1979. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, C.T.; Carvalho-de-Souza, J.L.; Schiavon, E.; Cassola, A.C.; Wanke, E.; Troncone, L.R. Crotamine inhibits preferentially fast-twitching muscles but is inactive on sodium channels. Toxicon 2007, 50, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Orts, D.J.; Prieto da Silva, A.R.; Oguiura, N.; Boni-Mitake, M.; de Oliveira, E.B.; Zaharenko, A.J.; de Freitas, J.C.; Tytgat, J. Crotamine pharmacology revisited: Novel insights based on the inhibition of KV channels. Mol. Pharmacol. 2012, 82, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Kerkis, I.; Hayashi, M.A.; Prieto da Silva, A.R.; Pereira, A.; De Sá Júnior, P.L.; Zaharenko, A.J.; Rádis-Baptista, G.; Kerkis, A.; Yamane, T. State of the art in the studies on crotamine, a cell penetrating peptide from South American rattlesnake. BioMed. Res. Int. 2014, 2014, 675985. [Google Scholar] [CrossRef]
- Türkaydin, B.; Schewe, M.; Riel, E.B.; Schulz, F.; Biedermann, J.; Baukrowitz, T.; Sun, H. Atomistic mechanism of coupling between cytosolic sensor domain and selectivity filter in TREK K2P channels. Nat. Commun. 2024, 15, 4628. [Google Scholar] [CrossRef] [PubMed]
- García, G.; Martínez-Rojas, V.A.; Murbartián, J. TREK-1 potassium channels participate in acute and long-lasting nociceptive hypersensitivity induced by formalin in rats. Behav. Brain Res. 2021, 413, 113446. [Google Scholar] [CrossRef] [PubMed]
- Borrull, A.; Allard, B.; Wijkhuisen, A.; Herbet, A.; Lamourette, P.; Birouk, W.; Leiber, D.; Tanfin, Z.; Ducancel, F.; Boquet, D.; et al. Rendomab B4, a monoclonal antibody that discriminates the human endothelin B receptor of melanoma cells and inhibits their migration. mAbs 2016, 8, 1371–1385. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, T.C.; Benoit, E.; Kurz, M.; Lucarain, L.; Fouconnier, S.; Combemale, S.; Jaquillard, L.; Schombert, B.; Chambard, J.M.; Boukaiba, R.; et al. From identification to functional characterization of cyriotoxin-1a, an antinociceptive toxin from the spider Cyriopagopus schioedtei. Br. J. Pharmacol. 2019, 176, 1298–1314. [Google Scholar] [CrossRef] [PubMed]
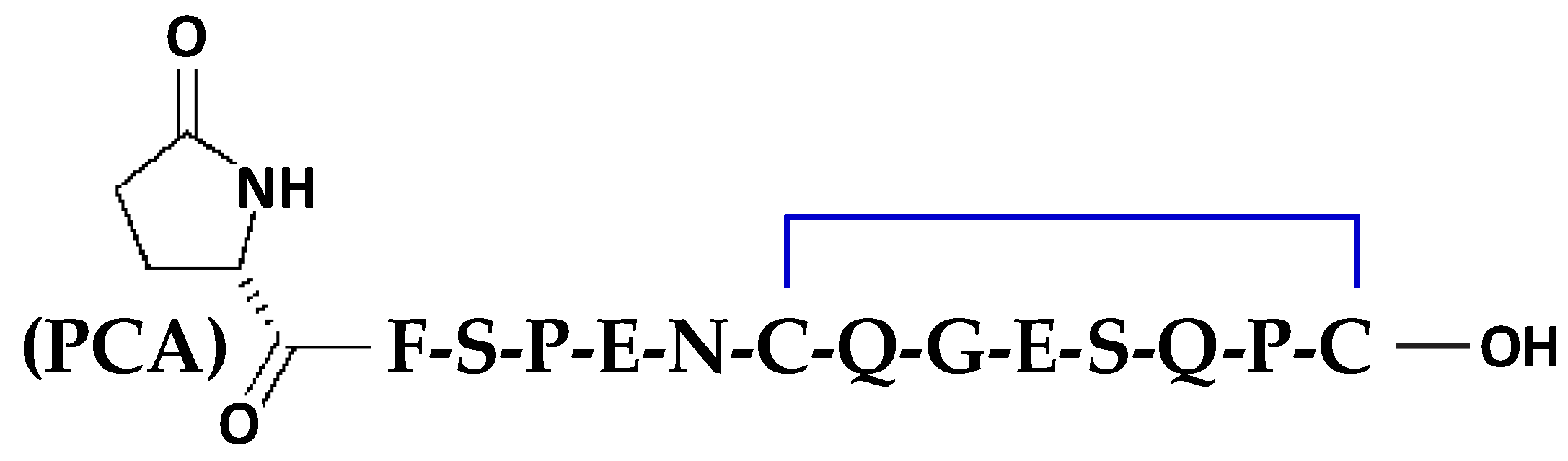




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Activation | Inactivation | |||
|---|---|---|---|---|
| TimeA (ms) | SlopeA (nA/ms) | TimeI (ms) | SlopeI (nA/ms) | |
| Standard physiological solution | 0.66 ± 0.06 | −3.33 ± 0.76 | 6.30 ± 0.67 | 0.34 ± 0.05 |
| Crotalphine (50 µM) | 0.68 ± 0.03 | −3.14 ± 0.33 | 6.67 ± 0.62 | 0.31 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antunes, A.; Robin, P.; Mourier, G.; Béroud, R.; De Waard, M.; Servent, D.; Benoit, E. Rattlesnake Crotalphine Analgesic Active on Tetrodotoxin-Sensitive Na+ Current in Mouse Dorsal Root Ganglion Neurons. Toxins 2024, 16, 359. https://doi.org/10.3390/toxins16080359
Antunes A, Robin P, Mourier G, Béroud R, De Waard M, Servent D, Benoit E. Rattlesnake Crotalphine Analgesic Active on Tetrodotoxin-Sensitive Na+ Current in Mouse Dorsal Root Ganglion Neurons. Toxins. 2024; 16(8):359. https://doi.org/10.3390/toxins16080359
Chicago/Turabian StyleAntunes, Aurélie, Philippe Robin, Gilles Mourier, Rémy Béroud, Michel De Waard, Denis Servent, and Evelyne Benoit. 2024. "Rattlesnake Crotalphine Analgesic Active on Tetrodotoxin-Sensitive Na+ Current in Mouse Dorsal Root Ganglion Neurons" Toxins 16, no. 8: 359. https://doi.org/10.3390/toxins16080359
APA StyleAntunes, A., Robin, P., Mourier, G., Béroud, R., De Waard, M., Servent, D., & Benoit, E. (2024). Rattlesnake Crotalphine Analgesic Active on Tetrodotoxin-Sensitive Na+ Current in Mouse Dorsal Root Ganglion Neurons. Toxins, 16(8), 359. https://doi.org/10.3390/toxins16080359