A Label Free Colorimetric Assay for the Detection of Active Botulinum Neurotoxin Type A by SNAP-25 Conjugated Colloidal Gold

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Coating of Colloidal Gold
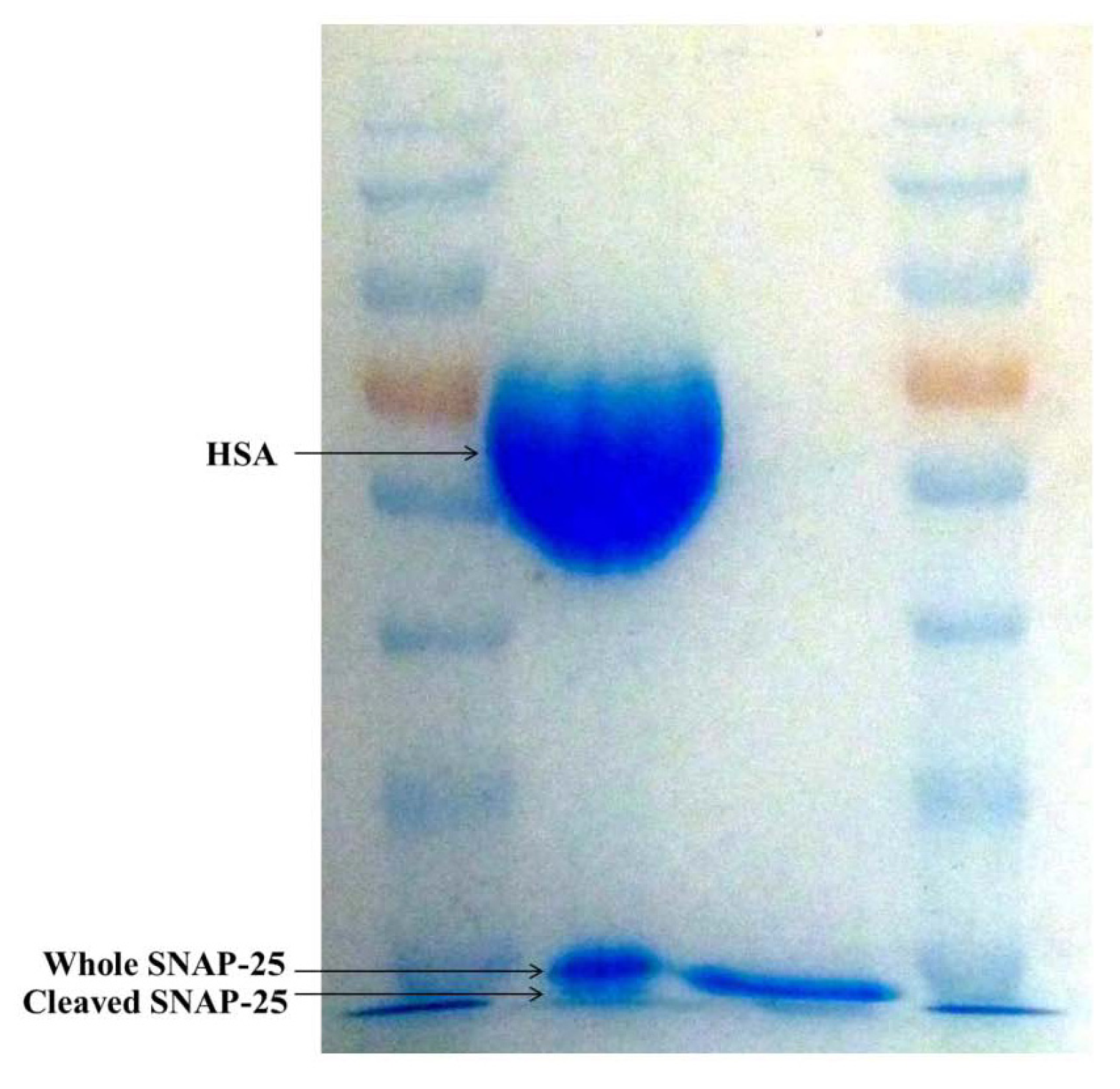
2.2. SDS-PAGE

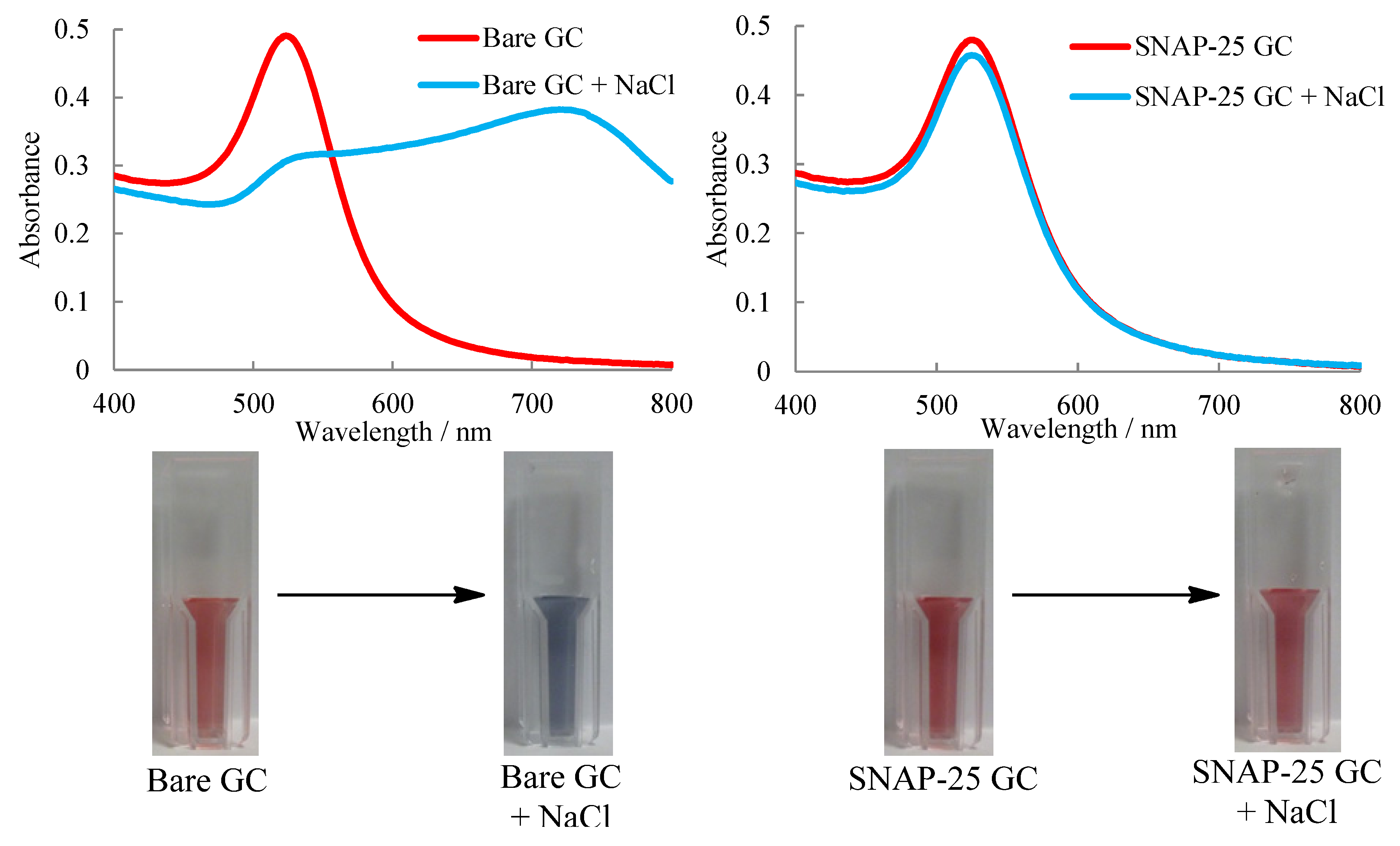
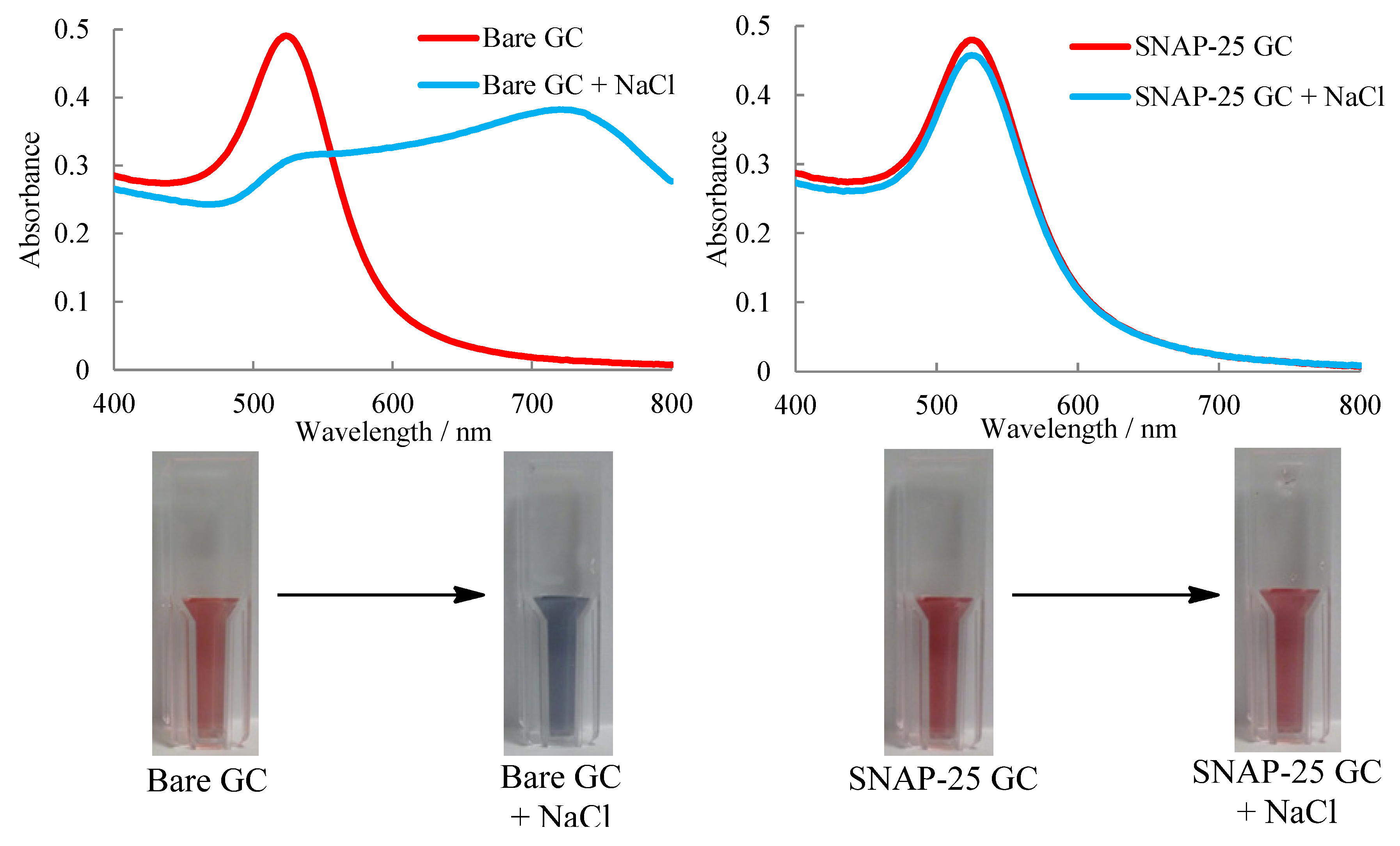
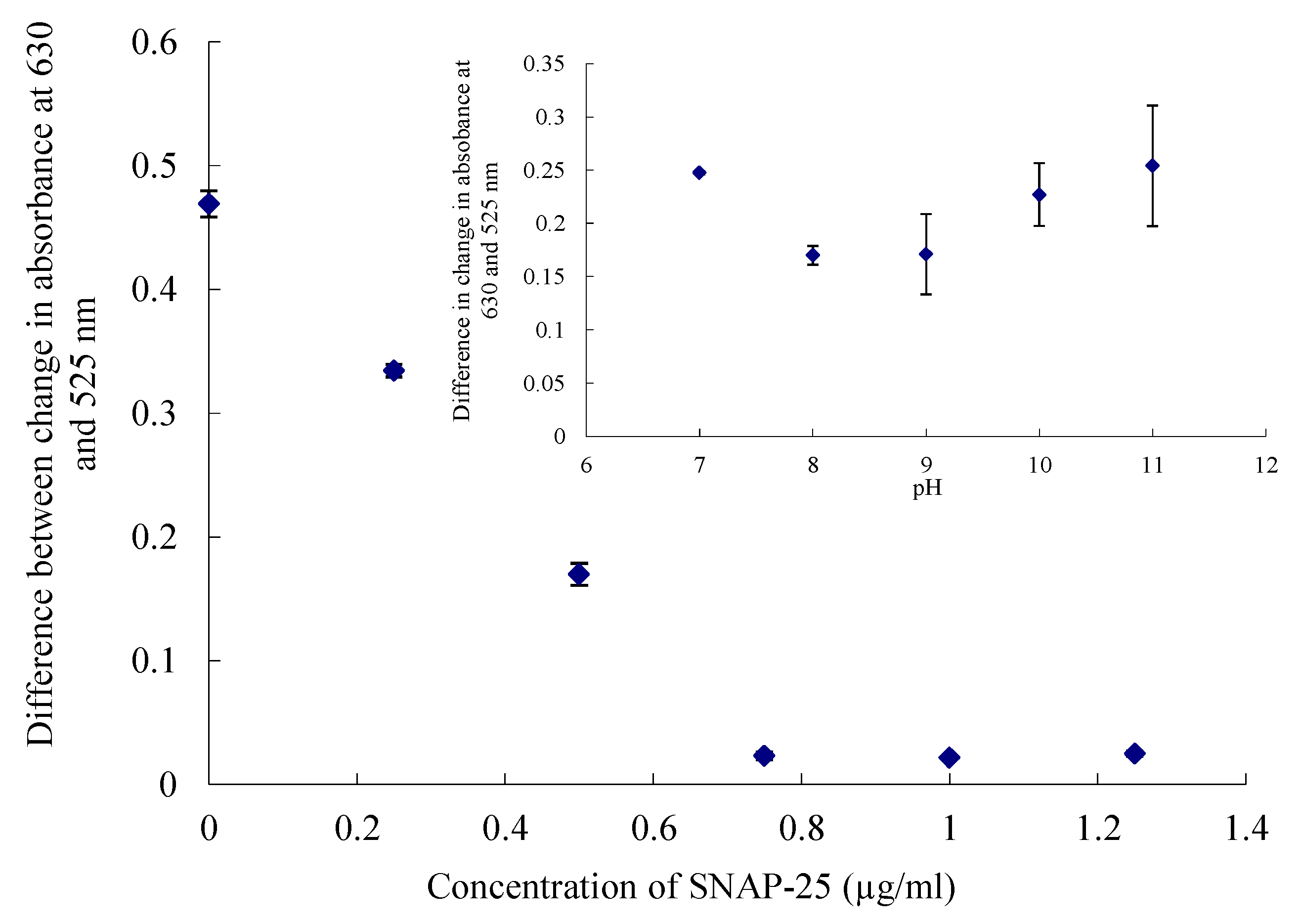
2.3. UV-Visible Spectroscopy Assay

2.4. Microplate Assay
3. Methods and Materials
3.1. Coating Colloidal Gold
3.2. Pre-treatment of Samples
3.3. SDS-PAGE
3.4. UV-Visible Spectroscopy Assay
3.5. Microplate Assay
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Pellett, S.; Tepp, W.H.; Toth, S.I.; Johnson, E.A. Comparison of the primary rat spinal cord cell (RSC) assay and the mouse bioassay for botulinum neurotoxin type A potency determination. J. Pharmacol. Toxicol. Method. 2010, 61, 304–310. [Google Scholar] [CrossRef]
- Rasooly, R.; Do, P.M. Development of an in vitro activity assay as an alternative to the mouse bioassay for Clostridium botulinum neurotoxin type A. Appl. Environ. Microbiol. 2008, 74, 4309–4313. [Google Scholar] [CrossRef]
- Pires-Alves, M.; Ho, M.; Aberle, K.K.; Janda, K.D.; Wilson, B.A. Tandem fluorescent proteins as enhanced FRET-based substrates for botulinum neurotoxin activity. Toxicon 2009, 53, 392–399. [Google Scholar] [CrossRef]
- Moorthy, J.; Mensing, G.A.; Kim, D.; Mohanty, S.; Eddington, D.T.; Tepp, W.H.; Johnson, E.A.; Beebe, D.J. Microfluidic tectonics platform: A colorimetric, disposable botulinum toxin enzyme-linked immunosorbent assay system. Electrophoresis 2004, 25, 1705–1713. [Google Scholar] [CrossRef]
- Ching, K.H.; Lin, A.; McGarvey, J.A.; Stanker, L.H.; Hnasko, R. Rapid and selective detection of botulinum neurotoxin serotype-A and -B with a single immunochromatographic test strip. J. Immunol. Method. 2012, 380, 23–29. [Google Scholar] [CrossRef]
- Katona, P. Botulinum toxin: Therapeutic agent to cosmetic enhancement to lethal biothreat. Anaerobe 2012, 18, 240–243. [Google Scholar] [CrossRef]
- Kukreja, R.V.; Singh, B.R. Comparative role of neurotoxin-associated proteins in the structural stability and endopeptidase activity of botulinum neurotoxin complex types A and E. Biochemistry 2007, 46, 14316–14324. [Google Scholar] [CrossRef]
- Montal, M. Botulinum neurotoxin: A marvel of protein design. Annu. Rev. Biochem. 2010, 79, 591–617. [Google Scholar] [CrossRef]
- Chen, S. Clinical uses of botulinum neurotoxins: Current indications, limitations and future developments. Toxins 2012, 4, 913–939. [Google Scholar] [CrossRef]
- Dasary, S.S.R.; Singh, A.K.; Senapati, D.; Yu, H.; Ray, P.C. Gold nanoparticle based label-free SERS probe for ultrasensitive and selective detection of trinitrotoluene. J. Am. Chem. Soc. 2009, 131, 13806–13812. [Google Scholar]
- Bajaj, A.; Miranda, O.R.; Kim, I.-B.; Phillips, R.L.; Jerry, D.J.; Bunz, U.H.F.; Rotello, V.M. Detection and differentiation of normal, cancerous, and metastatic cells using nanoparticle-polymer sensor arrays. Proc. Nat. Acad. Sci. USA 2009, 106, 10912–10916. [Google Scholar]
- Xu, H.; Mao, X.; Zeng, Q.; Wang, S.; Kawde, A.-N.; Liu, G. Aptamer-functionalized gold nanoparticles as probes in a dry-reagent strip biosensor for protein analysis. Anal. Chem. 2009, 81, 669–675. [Google Scholar] [CrossRef]
- Zhao, W.; Brook, M.A.; Li, Y. Design of gold nanoparticle-based colorimetric biosensing assays. Chembiochem 2008, 9, 2363–2371. [Google Scholar] [CrossRef]
- Li, L.; Li, B.; Qi, Y.; Jin, Y. Label-free aptamer-based colorimetric detection of mercury ions in aqueous media using unmodified gold nanoparticles as colorimetric probe. Anal. Bioanal. Chem. 2009, 393, 2051–2057. [Google Scholar] [CrossRef]
- Wang, G.; Sun, W. Optical limiting of gold nanoparticle aggregates induced by electrolytes. J. Phys. Chem. B 2006, 110, 20901–20905. [Google Scholar] [CrossRef]
- Dominguez-Medina, S.; Blankenburg, J.; Olson, J.; Landes, C.F.; Link, S. Adsorption of a protein monolayer via hydrophobic interactions prevents nanoparticle aggregation under harsh environmental conditions. ACS Sustain. Chem. Eng. 2013, 1, 833–842. [Google Scholar]
- Chen, S.; Barbieri, J.T. Association of botulinum neurotoxin serotype a light chain with plasma membrane-bound SNAP-25. J. Biol. Chem. 2011, 286, 15067–15072. [Google Scholar] [CrossRef]
- Greaves, J.; Chamberlain, L.H. Differential palmitoylation regulates intracellular patterning of SNAP25. J. Cell Sci. 2011, 124, 1351–1360. [Google Scholar] [CrossRef]
- Breidenbach, M.A.; Brunger, A.T. Substrate recognition strategy for botulinum neurotoxin serotype A. Nature 2004, 432, 925–929. [Google Scholar] [CrossRef]
- Gwenin, C.D.; Kalaji, M.; Williams, P.A.; Jones, R.M. The orientationally controlled assembly of genetically modified enzymes in an amperometric biosensor. Biosens. Bioelectron. 2007, 22, 2869–2875. [Google Scholar] [CrossRef]
- Chen, S.; Barbieri, J.T. Unique substrate recognition by botulinum neurotoxins serotypes A and E. J. Biol. Chem. 2006, 281, 10906–10911. [Google Scholar] [CrossRef]
- Zhang, F.; Skoda, M.W.A.; Jacobs, R.M.J.; Dressen, D.G.; Martin, R.A.; Martin, C.M.; Clark, G.F.; Lamkemeyer, T.; Schreiber, F. Gold nanoparticles decorated with oligo(ethylene glycol) thiols: Enhanced hofmeister effects in colloid-protein mixtures. J. Phys. Chem. C 2009, 113, 4839–4847. [Google Scholar] [CrossRef]
- Cai, S.O.; Sarkar, H.K.; Singh, B.R. Enhancement of the endopeptidase activity of botulinum neurotoxin by its associated proteins and dithiothreitol. Biochemistry 1999, 38, 6903–6910. [Google Scholar] [CrossRef]
- Chen, S.; Kim, J.-J.P.; Barbieri, J.T. Mechanism of substrate recognition by botulinum neurotoxin serotype A. J. Biol. Chem. 2007, 282, 9621–9627. [Google Scholar] [CrossRef]
- Armbruster, D.A.; Pry, T. Limit of blank, limit of detection and limit of quantitation. Clin. Biochem. Rev. 2008, 29, S49–S52. [Google Scholar]
- Hunt, T.; Rupp, D.; Shimizu, G.; Tam, K.; Weidler, J.; Xie, J. Characterization of SNARE cleavage products generated by formulated botulinum neurotoxin type-a drug products. Toxins 2010, 2, 2198–2212. [Google Scholar]
- Davis, L.E. Botulism. Curr. Treat Opt. Neurol. 2003, 5, 23–31. [Google Scholar] [CrossRef]
- Liu, Y.Y.B.; Rigsby, P.; Sesardic, D.; Marks, J.D.; Jones, R.G.A. A functional dual-coated (FDC) microtiter plate method to replace the botulinum toxin LD50 test. Anal. Biochem. 2012, 425, 28–35. [Google Scholar] [CrossRef]
- Dunning, F.M.; Ruge, D.R.; Piazza, T.M.; Stanker, L.H.; Zeytin, F.N.; Tucker, W.C. Detection of botulinum neurotoxin serotype A, B, and F proteolytic activity in complex matrices with picomolar to femtomolar sensitivity. Appl. Environ. Microbiol. 2012, 78, 7687–7697. [Google Scholar] [CrossRef]
- Moser, A.C.; Hage, D.S. Immunoaffinity chromatography: An introduction to applications and recent developments. Bioanalysis 2010, 2, 769–790. [Google Scholar] [CrossRef]
- Lebeda, F.J.; Cer, R.Z.; Mudunuri, U.; Stephens, R.; Singh, B.R.; Adler, M. The zinc-dependent protease activity of the botulinum neurotoxins. Toxins 2010, 2, 978–997. [Google Scholar]
- Strong, A.E.; Moore, B.D. Self-assembling monolayers of helical oligopeptides on gold with applications in molecular electronics. J. Mater. Chem. 1999, 9, 1097–1105. [Google Scholar] [CrossRef]
- Jones, R.G.A.; Liu, Y.; Halls, C.; Thorpe, S.J.; Longstaff, C.; Matejtschuk, P.; Sesardic, D. Release of proteolytic activity following reduction in therapeutic human serum albumin containing products: Detection with a new neoepitope endopeptidase immunoassay. J. Pharmaceut.Biomed. Anal. 2011, 54, 74–80. [Google Scholar] [CrossRef]
- Gwenin, C.D.; Halliwell, J. Detection of Botulinum Toxin. UK patent application No. 1310090.4, May 2013. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Halliwell, J.; Gwenin, C. A Label Free Colorimetric Assay for the Detection of Active Botulinum Neurotoxin Type A by SNAP-25 Conjugated Colloidal Gold. Toxins 2013, 5, 1381-1391. https://doi.org/10.3390/toxins5081381
Halliwell J, Gwenin C. A Label Free Colorimetric Assay for the Detection of Active Botulinum Neurotoxin Type A by SNAP-25 Conjugated Colloidal Gold. Toxins. 2013; 5(8):1381-1391. https://doi.org/10.3390/toxins5081381
Chicago/Turabian StyleHalliwell, Jennifer, and Christopher Gwenin. 2013. "A Label Free Colorimetric Assay for the Detection of Active Botulinum Neurotoxin Type A by SNAP-25 Conjugated Colloidal Gold" Toxins 5, no. 8: 1381-1391. https://doi.org/10.3390/toxins5081381
APA StyleHalliwell, J., & Gwenin, C. (2013). A Label Free Colorimetric Assay for the Detection of Active Botulinum Neurotoxin Type A by SNAP-25 Conjugated Colloidal Gold. Toxins, 5(8), 1381-1391. https://doi.org/10.3390/toxins5081381