Aflatoxin Control in Maize by Trametes versicolor


Abstract
:1. Introduction
2. Results and Discussion
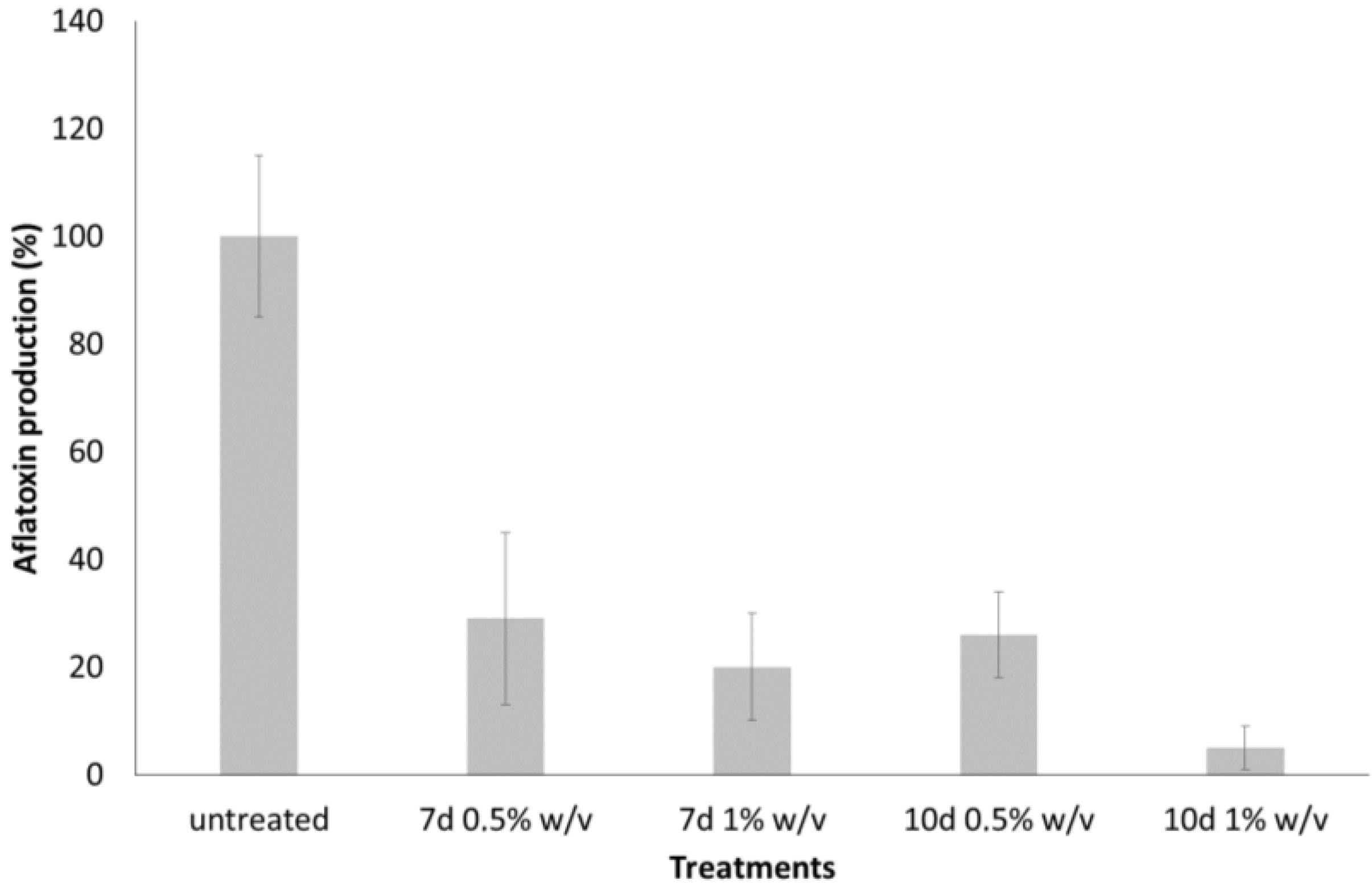
2.1. Inhibition of Aflatoxin B1 in Liquid Culture of A. flavus by LF and LS of T. versicolor


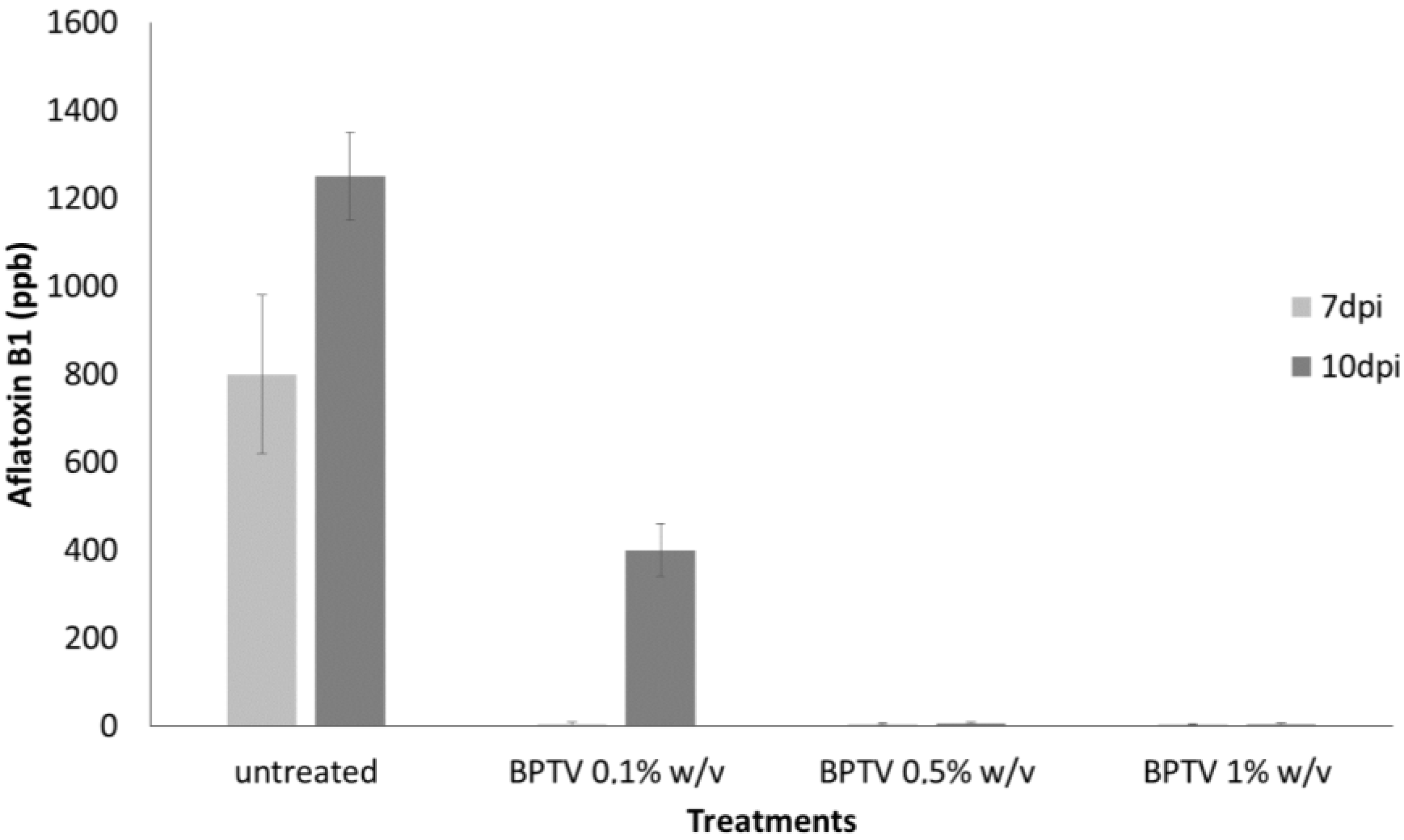
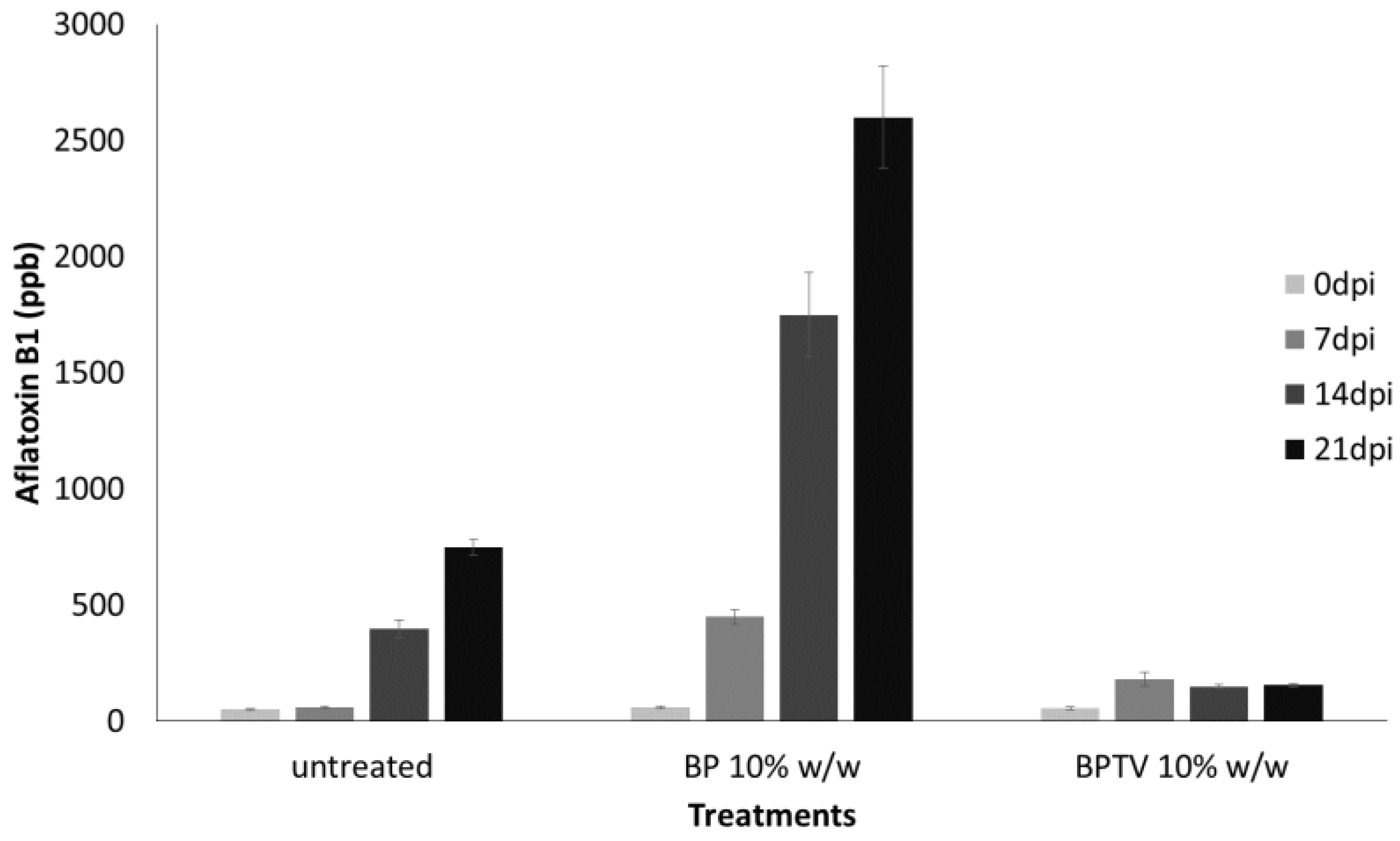
2.2. Inhibition of Aflatoxin B1 in Maize Kernels Contaminated by A. flavus by LF and LS of T. versicolor

2.3. Degradation of AFB1 by Lignin Degrading Enzymes of T. versicolor


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Aflatoxin B1 (ppb) | % Degradation |
|---|---|---|
| untreated | 1528 ± 250 | - |
| 3.5 U LAC | 760 ± 52 | 50 ± 6 |
| 7 U LAC | 458 ± 40 | 70 ± 11 |
| Time (days) | Aflatoxin B1 (ppb) | |||
|---|---|---|---|---|
| Seeds | % degradation | milled seeds | % degradation | |
| 0 | 74 ± 5 | - | 140 ± 4 | - |
| 7 | 53 ± 3 | 30 ± 6 | 80 ± 2 | 44 ± 3 |
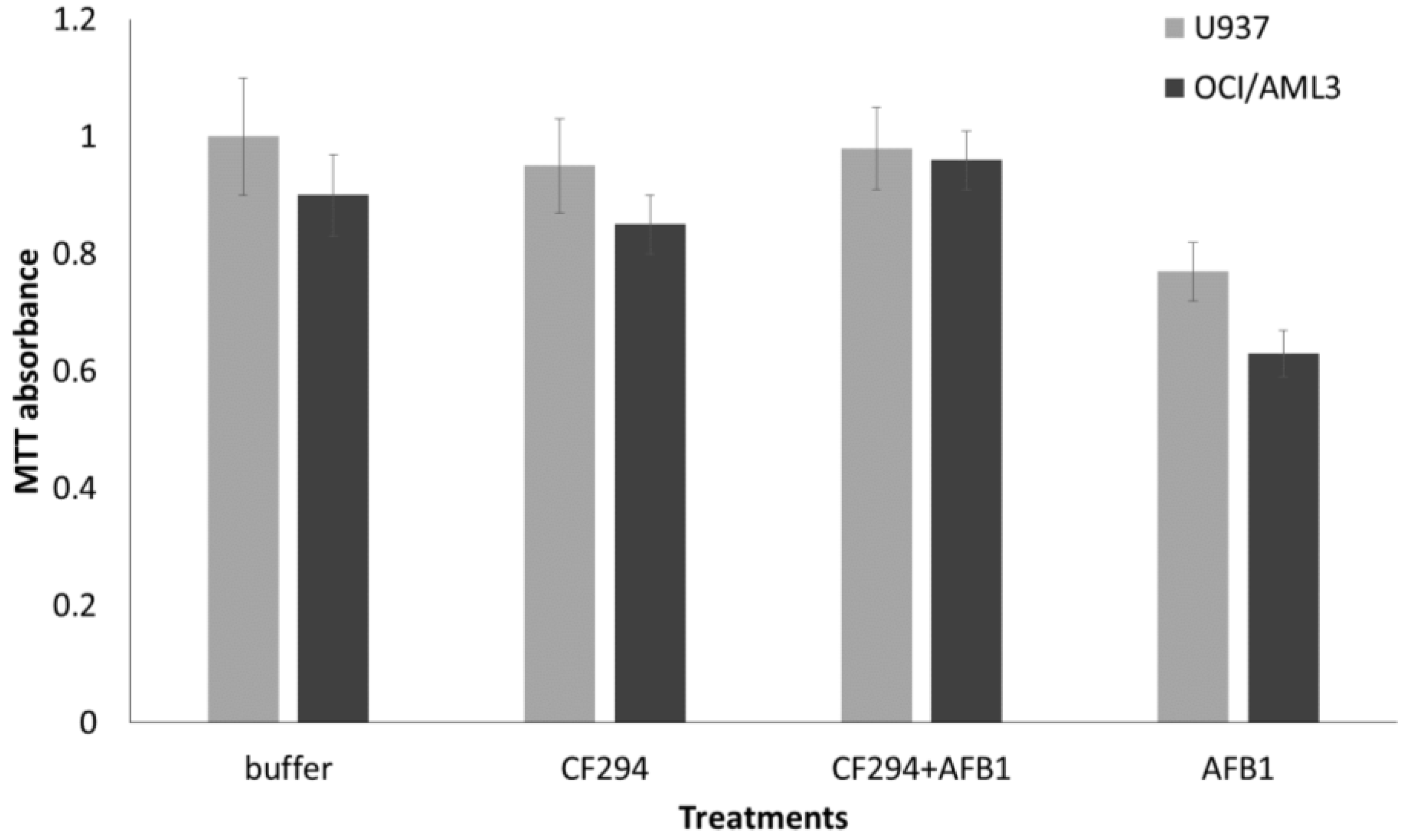
2.4. Human Cell Viability Assays

3. Materials and Methods
3.1. Fungal Strains
3.2. Assays of T. versicolor Culture Filtrates on Aflatoxin Production: “In vitro Experiments”
3.3. Assays of T. versicolor Solid Substrate on Aflatoxin Production: “In vitro Experiments”
3.4. Assays of T. versicolor Culture Filtrates on Aflatoxin Degradation: “In vitro Experiments”
3.5. Experiments on Maize and Seeds
3.6. Assays of T. versicolor Culture Filtrates on Aflatoxin Degradation: “In vivo Experiments”
3.7. Cell Viability Assays
3.8. Analytical Determination
3.9. Laccase Activity
3.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bryden, W.L. Mycotoxins in the food chain: Human health implications. Mutagenesis 2002, 17, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Hell, K.; Mutegi, C. Aflatoxin control and prevention strategies in key crops of Sub-Saharan Africa. Afr. J. Microbiol. Res. 2011, 5, 459–466. [Google Scholar]
- Williams, W.P.; Krakowsky, M.; Windham, G.; Balint-Kurti, P.; Hawkins, L.; Henry, W.B. Identifying maize germplasm with resistance to aflatoxin accumulation. Toxin Rev. 2008, 27, 319–345. [Google Scholar] [CrossRef]
- Luo, M.; Liu, J.; Lee, D.; Scully, B.T.; Guo, B. Monitoring the expression of maize genes in developing kernels under drought stress using oligo-microarray. J. Integr. Plant Biol. 2010, 52, 1059–1074. [Google Scholar] [CrossRef] [PubMed]
- Kelley, R.Y.; Williams, W.P.; Mylroie, J.E.; Boykin, D.L.; Harper, J.W.; Windham, G.L.; Ankala, A.; Shan, X. Identification of maize genes associated with host plant resistance or susceptibility to Aspergillus flavus infection and aflatoxin accumulation. PLoS One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Camardo-Leggieri, M.; Rossi, V.; Giorni, P. AFLA-maize, a mechanistic model for Aspergillus flavus infection and aflatoxin B1 contamination in maize. Comput. Electron. Agric. 2013, 94, 38–46. [Google Scholar] [CrossRef]
- Mauro, A.; Battilani, P.; Callicott, K.A.; Giorni, P.; Pietri, A.; Cotty, P.J. Structure of an Aspergillus flavus population from maize kernels in northern Italy. Int. J. Food Microbiol. 2013, 162, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Boyd, I.L.; Freer-Smith, P.H.; Gilligan, C.A.; Godfray, H.C. The consequence of tree pests and diseases for ecosystem services. Science 2013, 342. [Google Scholar] [CrossRef]
- Matarese, F.; Sarrocco, S.; Gruber, S.; Seidl-Seiboth, V.; Vannacci, G. Biocontrol of Fusarium head blight: Interactions between Trichoderma and mycotoxigenic Fusarium. Microbiology 2012, 158, 98–106. [Google Scholar] [CrossRef]
- Probst, C.; Bandyopadhyay, R.; Cotty, P.J. Diversity of aflatoxin-producing fungi and their impact on food safety in sub-Saharan Africa. Int. J. Food Microbiol. 2014, 174, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Olarte, R.A.; Horn, B.W.; Dorner, J.W.; Monacell, J.T.; Singh, R.; Stone, E.A.; Carbone, I. Effect of sexual recombination on population diversity in aflatoxin production by Aspergillus flavus and evidence for cryptic heterokaryosis. Mol. Ecol. 2012, 21, 1453–1476. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Scala, V.; Fabbri, A.A.; Scarpari, M.; Fanelli, C.; Zjalic, S.; Nobili, C. Methodologies to prevent and control the biosynthesis of different mycotoxins in food and feeds. Acta Hort. (ISHS) 2011, 963, 83–86. [Google Scholar]
- Reverberi, M.; Fabbri, A.A.; Zjalic, S.; Ricelli, A.; Punelli, F.; Fanelli, C. Antioxidant enzymes stimulation in Aspergillus parasiticus by Lentinula edodes inhibits aflatoxin production. Appl. Microbiol. Biotechnol. 2005, 69, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Zjalic, S.; Reverberi, M.; Ricelli, A.; Granito, V.M.; Fanelli, C.; Fabbri, A.A. Trametes versicolor: A possible tool for aflatoxin control. Int. J. Food Microbiol. 2006, 107, 243–249. [Google Scholar] [CrossRef]
- Fanelli, C.; Ricelli, A.; Reverberi, M.; Fabbri, A.A. Aflatoxins and ochratoxins in cereal grains: An open challenge. In Recent Research Development in Crop Science; Pandalai, S.G., Ed.; Research Signpost: Kerala, India, 2004; Volume 1, pp. 295–317. [Google Scholar]
- Tomsovsky, M.; Homolka, L. Laccase and other ligninolytic enzymes activities of selected strains of Trametes spp. From different localities and substrates. Folia Microbiol. 2003, 48, 413–416. [Google Scholar] [CrossRef]
- Urbanelli, S.; Della Rosa, V.; Fanelli, C.; Fabbri, A.A.; Reverberi, M. Genetic diversity and population structure of the Italian fungi belonging to the taxa Pleurotus eryngii (DC.:FR.) Quèl and P. ferulae (DC.:Fr.) Quèl. Heredity 2003, 90, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Punelli, F.; Reverberi, M.; Porretta, D.; Nogarotto, S.; Fabbri, A.A.; Fanelli, C.; Urbanelli, S. Molecular characterization and enzymatic activity of laccases in two Pleurotus spp. with different pathogenic behaviour. Mycol. Res. 2009, 113, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P. Fungal laccase—Occurrence and properties. FEMS Microbiol. Rev. 2006, 30, 215–242. [Google Scholar] [CrossRef] [PubMed]
- Alberts, J.F.; Gelderblom, W.C.A.; Botha, A.; van Zyl, W.H. Degradation of aflatoxin B1 by fungal laccase enzymes. Int. J. Food Microbiol. 2009, 135, 47–52. [Google Scholar] [CrossRef]
- Reverberi, M.; Zjalic, S.; Punelli, F.; Ricelli, A.; Fabbri, A.A.; Fanelli, C. Apyap1 affects aflatoxin biosynthesis during Aspergillus parasiticus growth in maize seeds. Food Addit. Contam. 2007, 24, 1070–1075. [Google Scholar] [CrossRef] [PubMed]
- Xavier, A.M.R.B.; Tavares, A.P.M.; Ferreira, R.; Amado, F. Trametes versicolor growth and laccase induction with by-products of pulp and paper industry. Electron. J. Biotechnol. 2007, 10, 444–451. [Google Scholar] [CrossRef]
- Harkin, J.M.; Obst, J.R. Syringaldazine as effective reagent for detecting laccase and peroxidase in fungi. Experientia 1973, 29, 381–386. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scarpari, M.; Bello, C.; Pietricola, C.; Zaccaria, M.; Bertocchi, L.; Angelucci, A.; Ricciardi, M.R.; Scala, V.; Parroni, A.; Fabbri, A.A.; et al. Aflatoxin Control in Maize by Trametes versicolor. Toxins 2014, 6, 3426-3437. https://doi.org/10.3390/toxins6123426
Scarpari M, Bello C, Pietricola C, Zaccaria M, Bertocchi L, Angelucci A, Ricciardi MR, Scala V, Parroni A, Fabbri AA, et al. Aflatoxin Control in Maize by Trametes versicolor. Toxins. 2014; 6(12):3426-3437. https://doi.org/10.3390/toxins6123426
Chicago/Turabian StyleScarpari, Marzia, Cristiano Bello, Chiara Pietricola, Marco Zaccaria, Luigi Bertocchi, Alessandra Angelucci, Maria Rosaria Ricciardi, Valeria Scala, Alessia Parroni, Anna A. Fabbri, and et al. 2014. "Aflatoxin Control in Maize by Trametes versicolor" Toxins 6, no. 12: 3426-3437. https://doi.org/10.3390/toxins6123426
APA StyleScarpari, M., Bello, C., Pietricola, C., Zaccaria, M., Bertocchi, L., Angelucci, A., Ricciardi, M. R., Scala, V., Parroni, A., Fabbri, A. A., Reverberi, M., Zjalic, S., & Fanelli, C. (2014). Aflatoxin Control in Maize by Trametes versicolor. Toxins, 6(12), 3426-3437. https://doi.org/10.3390/toxins6123426