2.5. Biochemical Body and Organ Composition
Investigation on whole body composition revealed that the fat content in whole body homogenates of fish treated with the medium- and high-dose DON diet was increased compared to control fish (
Table 8), whereas in fish treated with the low dose DON diet, only a tendency for increased fat content was found (
p = 0.065). Still, a significant relationship between the toxin concentrations in the experimental feeds and lipid content was found (Spearman correlation coefficient, 0.498, and significance,
p = 0.013).
Table 7.
Lactate dehydrogenase (LDH; mU mg protein−1) activity in tissue homogenates after DON feeding and a two-week recovery phase; n = 6; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
Table 7.
Lactate dehydrogenase (LDH; mU mg protein−1) activity in tissue homogenates after DON feeding and a two-week recovery phase; n = 6; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
| LDH Activity | Basal Feed | Low DON | Medium DON | High DON |
|---|
| DON-treated: | | | | |
| head kidney | 31.58 ± 4.77 a | 71.55 ± 3.63 b | 61.22 ± 5.11 b | 51.89 ± 6.30 b |
| trunk kidney | 34.34 ± 5.18 a | 77.79 ± 3.95 b | 66.23 ± 4.55 b,c | 53.16 ± 6.84 a,c |
| spleen | 2.64 ± 0.25 | 2.97 ± 0.47 | 2.90 ± 0.34 | 2.92 ± 0.43 |
| liver | 275.23 ± 61.86 | 370.96 ± 66.37 | 176.58 ± 40.47 | 264.67 ± 56.73 |
| white muscle | 38.28 ± 13.68 a | 17.51 ± 5.77 a,b | 20.86 ± 7.39 a,b | 3.12 ± 1.81 b |
| Recovery: | | | | |
| head kidney | 51.21 ± 4.97 a | 52.54 ± 6.65 a | 57.12 ± 9.96 a | 13.34 ± 2.77 b |
| trunk kidney | 54.58 ± 6.08 a | 65.33 ± 6.10 a | 60.70 ± 13.15a | 19.89 ± 4.50 b |
| spleen | 10.88 ± 0.99 | 9.74 ± 0.90 | 7.92 ± 0.59 | 12.96 ± 2.86 |
| liver | 496.02 ± 118.64 | 669.79 ± 113.86 | 510.05 ± 106.11 | 344.31 ± 89.97 |
| white muscle | 17.73 ± 4.85 | 12.35 ± 4.57 | 22.29 ± 3.90 | 25.91 ± 3.92 |
After two additional weeks of recovery, all DON-treated fish showed lower lipid levels than the control fish (
Table 9). These results were paralleled by significant differences in the energy content of the whole body homogenates. After recovery, the ash contents of the whole body homogenates were also found to be different in fish fed the high-dose DON diet compared to control fish (
Table 9).
In the recovered fish, the ash content correlated with the lipid content negatively (Spearman correlation coefficient, −0.568; significance, p = 0.002). Moreover, a significant relationship between the toxin concentrations in the experimental feeds and the lipid and ash content was found (Spearman correlation coefficient, −0.792 and 0.568, and significance, p < 0.000 and p = 0.007, respectively) in fish after the recovery phase. This led to the investigation of similar parameters in samples of liver tissue and white dorsal musculature to allow a possible explanation for these observations.
Impairment of the intestinal nutrient uptake by DON was reported for mammalian systems [
25,
26,
27]. Since the liver is an important metabolic organ that processes nutrients from feed, one would expect effects on the nutritional status of liver tissue if such impairment of intestinal nutrient uptake would also be present in the experimental carp.
Therefore, the biochemical composition of liver tissue of carp was analysed. Dry matter values of liver tissue were comparable to the values reported for other cyprinid species, such as freshwater major carp,
Catla catla [
28]. No significant differences in the dry matter of DON-treated fish compared to control fish were observed after four weeks of experiments, but an additional two weeks of recovery led to significantly different dry matter incorporations between fish fed the low-dose DON feed and the control fish (
Table 10). Liver total lipid contents are higher than in the whole body homogenates. This seems feasible, since liver in carp is known to be important for the storage of lipids. Moreover, in fish treated with the highest concentration of DON, the lipid content in liver tissue was significantly increased after the recovery phase compared to control fish. This is in contrast to the whole body lipid levels of DON-treated fish after the recovery phase and indicates that liver damage was accompanied by hepatic lipid accumulation, which also influenced the lipid balance of the entire body.
Table 8.
The whole body composition of experimental fish two weeks after DON feeding (recovery phase); n = 6; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
Table 8.
The whole body composition of experimental fish two weeks after DON feeding (recovery phase); n = 6; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
| Whole Body Composition | Basal Feed | Low DON | Medium DON | High DON |
|---|
| Crude lipid content (% dry matter) | 24.83 ± 1.85 a | 29.33 ± 1.36 a,b | 32.17 ± 0.95 b | 30.33 ± 0.95 b |
| Crude ash (% dry matter) | 8.56 ± 1.22 | 8.72 ± 0.19 | 8.03 ± 1.01 | 9.37 ± 0.34 |
| Energy content (MJ kg−1 dry matter) | 24.32 ± 0.47 a | 25.27 ± 0.34 a,b | 25.78 ± 0.25 b | 25.25 ± 0.31 a,b |
Table 9.
The whole body composition of experimental fish two weeks after DON feeding (recovery phase); n = 6; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
Table 9.
The whole body composition of experimental fish two weeks after DON feeding (recovery phase); n = 6; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
| Whole Body Composition | Basal Feed | Low DON | Medium DON | High DON |
|---|
| Crude lipid content (% dry matter) | 33.17 ± 0.75 a | 30.25 ± 0.48 b | 29.60 ± 1.08 b | 25.00 ± 2.71 b |
| Crude ash (% dry matter) | 8.23 ± 0.35 a | 8.46 ± 0.34 a,b | 9.37 ± 0.34 a,b | 9.98 ± 0.53 b |
| Energy content (MJ kg−1 dry matter) | 25.63 ± 0.12 a | 25.40 ± 0.19 a,b | 25.02 ± 0.28 a,b | 24.64 ± 0.23 b |
Table 10.
The composition of liver samples after four weeks of DON feeding and after the recovery phase of two weeks; n = 6; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
Table 10.
The composition of liver samples after four weeks of DON feeding and after the recovery phase of two weeks; n = 6; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
| Liver Composition | Basal Feed | Low DON | Medium DON | High DON |
|---|
| DON-treated: | | | | |
| Dry matter (% wet matter) | 25.9 ± 0.8 | 22.9 ± 2.2 | 25.6 ± 0.8 | 27.0 ± 1.4 |
| Total lipids (% wet matter) | 15.0 ± 1.3 | 14.5 ± 1.2 | 16.2 ± 1.7 | 17.3 ± 1.7 |
| Free glucose (mM g−1 wet matter) | 1.16 ± 0.27 | 0.87 ± 0.14 | 1.19 ± 0.08 | 0.91 ± 0.19 |
| Lactate (mM g−1 wet matter) | 1.75 ± 0.48 | 3.05 ± 1.29 | 3.77 ± 2.03 | 3.56 ± 1.87 |
| AST (U mg protein−1) | 90.3 ± 18.2 | 60.2 ± 20.5 | 81.7 ± 8.2 | 89.1 ± 16.6 |
| ALT (U mg protein−1) | 1.43 ± 0.14 a | 2.61 ± 0.98 a,b | 2.95 ± 0.51 b | 2.73 ± 0.49 a,b |
| Ascorbate (μM g−1 wet matter) | 81.7 ± 15.7 | 63.5 ± 8.3 | 71.5 ± 14.8 | 54.3 ± 8.8 |
| Recovery: | | | | |
| Dry matter (% wet matter) | 25.1 ± 0.5 a | 28.9 ± 1.4 a | 27.5 ± 1.8 a,b | 25.8 ± 1.7 a,b |
| Total lipids (% wet matter) | 17.7 ± 1.2 a | 19.2 ± 1.3 a,b | 18.7 ± 0.8 a,b | 23.6 ± 4.1 b |
| Free glucose (mM g−1 wet matter) | 0.73 ± 0.10 | 0.96 ± 0.17 | 0.91 ± 0.12 | 0.99 ± 0.21 |
| Lactate (mM g−1 wet matter) | 1.36 ± 0.35 a | 3.74 ± 1.13 a,b | 6.48 ± 2.02 b | 2.85 ± 0.80 a,b |
| AST (U mg protein−1) | 98.5 ± 22.1 | 96.5± 27.5 | 81.7 ± 8.2 | 99.7 ± 10.8 |
| ALT (U mg protein−1) | 2.47 ± 0.23 | 2.88 ± 0.22 | 2.79 ± 0.11 | 3.03 ± 0.26 |
| Ascorbate (μM g−1 wet matter) | 51.8 ± 6.5 | 58.8 ± 7.0 | 64.9 ± 7.5 | 62.2 ± 7.6 |
Liver glucose levels were not significantly different between treatment groups. It was suggested that amino acids are a superior energy sources to glucose for carp [
29]. It has also been shown for carps under starvation that the conversion of lipid to glycogen in liver tissue was accompanied by an increase in blood glucose levels [
30]. No changes of blood glucose (
Table 11) and lipid (
Table 10) in liver tissue were observed in DON-treated fish, which suggests that blood homeostasis was not subjected to a fasting-like status, due to an impairment of nutrient uptake in the intestine.
Liver glucose, lactate and ascorbate concentrations were not influenced by DON feeding and did not correlate with toxin concentrations in the experimental feeds. Although ascorbic acid has been shown to prevent the hemolytic action of DON on rat erythrocytes to some extent [
22], ascorbate levels in carp remained unchanged, which indicated that ascorbate alone is not sufficient to prevent damage due to DON exposure. Liver alanine aminotransferase (ALT) activity was increased by treatment with the medium-dose DON diet for four weeks compared to control fish, while the other DON-treated groups showed no significant difference of this enzyme activity compared to the control group, probably due to higher individual variation (coefficient of variance (
CV) for control fish of 24.6
versus CVs of 92.2, 42.3 and 51.0 for fish treated with the low-, medium- and high-dose, respectively, while the
CVs for all fish in the recovery phase ranged from 9.5 to 23.2). ALT activity also correlated with the mycotoxin concentrations in feed (Spearman correlation coefficient, −0.409; significance,
p = 0.047). This indicates damage to liver tissue, as has already been observed in carp under chemical stress [
31]. In contrast, aspartate aminotransferase (AST) activity was not significantly influenced by DON feeding, which indicates that the rate of amino acid transformations via transamination is not influenced.
A considerable amount of the fish consists of white musculature, which shows low levels of myoglobin and is mostly used for burst swimming [
32]. Swimming performance is known to lead to the utilization of nutrients from blood circulation and white and red musculature [
33]. Several factors further influence the chemical body composition. For example, the genetic background of carp determined the dry matter content of the fillet, as has been shown for different crossbreds of common carp, accounting for 19% to 28% [
34,
35,
36]. With respect to common mirror carps, a value for dry matter of 22.0% was observed [
36]. The dry matter of the carp used in our study corresponds to this value and was only influenced by feeding the low-dose DON diet (
Table 12).
As expected, the lipid content in carp muscle was found to be low in the present study, and it was not influenced by DON feeding. Different crossbreds of common carp showed lipid contents in the fillet of up to 9.9%, with mirror carps showing 2.4% lipid in fillet [
36]. Low muscle lipid contents ranging from 0.5% to 2.6% have also been noted by another study [
34]. However, slightly higher values, ranging from 2.9% to 5.2%, have been reported elsewhere [
35,
37]. Differences in the chemical composition of carp musculature can be due to the influence of rearing conditions [
38,
39], the influence of age [
35,
36,
39] and differences in the composition of the diet [
37,
39,
40,
41], and these factors should be considered when the values for the present study are compared to other studies. For example, in the study of Steffens and Wirth [
40], the addition of 10% different lipid sources in the diet led to 2.2 to 2.5% lipid in dorsal muscle of carp, which corresponds to the values in the present study.
The significant difference in the dry matter of white muscle of fish fed the low-dose diet compared to the control fish cannot be explained at the moment. Reductions of dry matter in the musculature can be caused by the depletion of tissue nutrient contents, which are compensated for by increasing the water content [
29]. However, even after calculation of the dry weight lipid contents in white muscle, the samples do not show a significant influence of DON. Thus, the reason for this observation remains obscure.
Muscle glucose shows no differences of the DON treatment of fish. The glucose concentration in red musculature has been reported to be rather independent of blood glucose levels [
42], but the aerobic glucose utilization was still assumed to be relying by approximately 30% on glucose in circulation. In contrast, white musculature in fish is known to largely depend on anaerobic glycogenolysis for energy liberation [
43]. Lactate in carp white musculature ranged from four to 9 mM [
44], even after exhaustive exercise. The lactate levels in muscle tissue in the present study were a bit lower and were not influenced by DON feeding. This might be due to the fact that even after severe hypoxic stress, lactate did not accumulate in white muscle, but was probably transferred out of the tissue [
45].
Ascorbate levels in white muscle samples strongly depend on the supply via the diet and have been reported to be low in fish musculature [
46]. This corresponds to the present study, although higher values have been reported for carp previously [
47]. This study also reported an influence of the mycotoxin, sterigmatocystin, on ascorbate levels in the white musculature of carp, which was not observed after DON feeding of carp in the present study. Similar to the ascorbate levels in liver tissue, the lack of influence of DON on their concentration in muscle further indicates that ascorbate as an endogenous antioxidant does not prevent the detrimental effects of DON on carp.
2.6. Serum Parameter
Our study demonstrates a large variation of biochemical serum parameters, which are known to be affected by many endogenous and exogenous factors, such as age, health condition, nutrition or stress, including chemical stress [
48,
49].
Glucose levels in the experimental fish in the present study were comparable to values for unstressed carp reported previously [
48,
49]. Glucose concentrations in serum remained unchanged in fish treated with DON for four weeks (
Table 11). A direct influence of DON on glucose metabolism has been shown in the human epithelial intestinal cell line, HT-29-D4, and in jejunum of chicken; however, cholesterol metabolism remained unchanged in this cell line [
26,
27]. No evidence for a similar impairment of glucose from the diet could be observed in the present study, although a possible decrease in blood glucose might also have been compensated for in DON-treated fish. Similar to other fish, carp are known to maintain blood glucose levels, even after prolonged starvation [
29].
Serum lactate levels in control fish are comparable to values in unstressed fish in other studies [
49,
50]. However, lactate values in DON-treated carp were considerably higher. This was probably caused by an activation of gluconeogenesis to maintain levels of circulation glucose. Consequently, elevated serum glucose levels can be caused [
49], which has also been observed in fish recovering for the treatment with the medium- and high-dose diet compared to the control fish (
Table 11). Thus, it can be assumed that DON affects anaerobic metabolism in carp muscle.
Table 11.
The serum parameter in DON-treated fish and in fish after a recovery of two weeks; n = 6 per group; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
Table 11.
The serum parameter in DON-treated fish and in fish after a recovery of two weeks; n = 6 per group; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
| Serum Parameters | Basal Feed | Low DON | Medium DON | High DON |
|---|
| DON-treated: | | | | |
| Free glucose (μM mL−1) | 2.22 ± 0.22 | 2.37 ± 0.19 | 2.12 ± 0.09 | 2.53 ± 0.27 |
| Lactate (μM mL−1) | 8.04 ± 2.87 a | 21.85 ± 5.78 a,b | 20.50 ± 6.70 a,b | 29.96 ± 8.44 b |
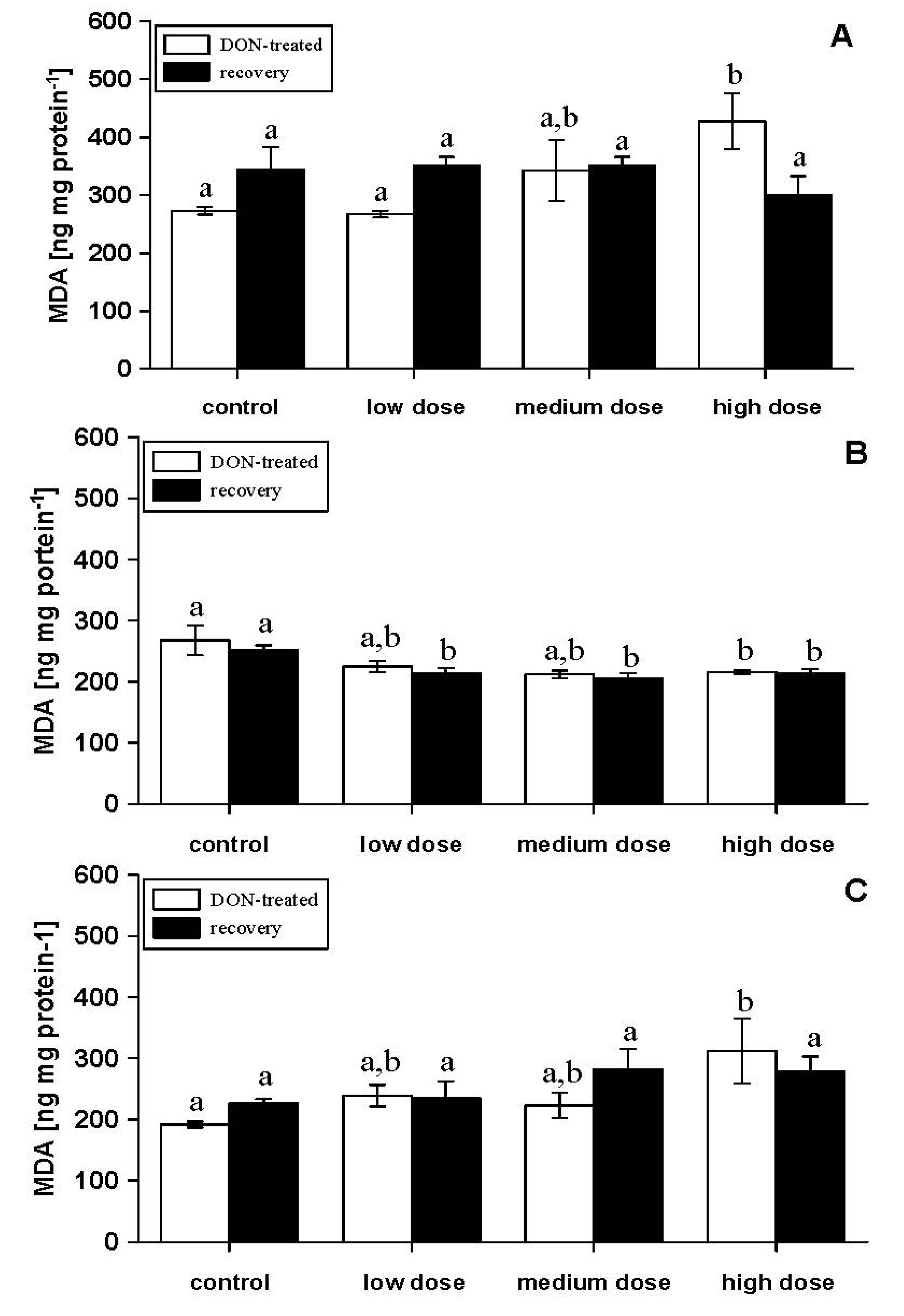
| LDH (mU mg protein−1) | 15.7 ± 1.0 a | 19.8 ± 1.8 a,b | 18.6 ± 2.5 a,b | 23.6 ± 2.1 b |
| SDH (mU mg protein−1) | 11.2 ± 4.4 a,b | 30.3 ± 4.8 a | 23.1 ± 8.2 a,b | 8.5 ± 2.1 b |
| AST (U mg protein−1) | 7.1 ± 0.9 a | 5.1 ± 0.2 a,b | 4.8 ± 0.7 b | 10.6 ± 3.8 a,c |
| ALT (U mg protein−1) | 0.7 ± 0.2 a | 0.7 ± 0.3 a | 1.2 ± 0.6 a | 0.2 ± 0.0 b |
| Total protein (mg mL−1) | 23.05 ± 0.73 | 23.32 ± 0.64 | 26.22 ± 1.26 | 23.41 ± 0.78 |
| Albumin (mg mL−1) | 19.85 ± 1.49 a | 15.14 ± 2.03 a,b | 5.42 ± 2.40 b | 6.92 ± 3.66 b |
| Recovery: | | | | |
| Free glucose (μM mL−1) | 1.66 ± 0.12 a | 2.78 ± 0.28 b | 2.84 ± 0.33 b | 2.77 ± 0.25 b |
| Lactate (μM mL−1) | 4.12 ± 1.08 a | 34.87 ± 12.74 b | 39.70 ± 8.35 b | 19.83 ± 3.70 b |
| LDH (mU mg protein−1) | 14.8 ± 1.4 a | 18.8 ± 1.0 b | 17.4 ± 0.9 a,b | 21.9 ± 1.6 b |
| SDH (mU mg protein−1) | 32.4 ± 8.8 | 16.1 ± 3.5 | 20.3 ± 5.4 | 34.5 ± 11.7 |
| AST (U mg protein−1) | 6.4 ± 0.8 | 5.1 ± 0.2 | 4.8 ± 0.7 | 7.9 ± 2.6 |
| ALT (U mg protein−1) | 0.4 ± 0.1 a | 0.6 ± 0.1 a | 0.7 ± 0.2 a,b | 1.1 ± 0.2 b |
| Total protein (mg mL−1) | 21.89 ± 0.73 | 22.04 ± 0.39 | 21.47 ± 1.86 | 29.86 ± 0.57 |
| Albumin (mg mL−1) | 15.32 ± 1.50 | 17.22 ± 1.74 | 16.94 ± 3.75 | 22.45 ± 2.40 |
Table 12.
The composition of samples from dorsal white musculature after four weeks of DON feeding and after the recovery phase of two weeks; n = 6; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
Table 12.
The composition of samples from dorsal white musculature after four weeks of DON feeding and after the recovery phase of two weeks; n = 6; mean ± SEM; means with the same letter (a and/or b) are not significantly different from each other (significance tested with Mann-Whitney U-tests, p < 0.05).
| Composition of White Muscle | Basal Feed | Low DON | Medium DON | High DON |
|---|
| DON-treated: | | | | |
| Dry matter (% wet matter) | 22.4 ± 2.0 a | 19.0 ± 0.7 b | 20.9 ± 0.6 a,b | 20.5 ± 0.4 a,b |
| Total lipids (% wet matter) | 2.4 ± 0.4 | 2.2 ± 0.2 | 2.4 ± 0.1 | 2.1 ± 0.2 |
| Free glucose (μM g−1 wet matter) | 2.93 ± 0.39 | 3.30 ± 0.22 | 2.95 ± 0.29 | 2.76 ± 0.29 |
| Lactate (mM g−1 wet matter) | 3.54 ± 0.37 | 4.08 ± 0.29 | 4.56 ± 0.30 | 3.79 ± 0.21 |
| Ascorbate (μM g−1 wet matter) | 10.2 ± 0.5 | 10.0 ± 0.8 | 10.3 ± 0.5 | 9.5 ± 0.6 |
| Recovery: | | | | |
| Dry matter (% wet matter) | 19.6 ± 0.9 | 19.7 ± 0.3 | 21.2 ± 0.6 | 19.7 ± 0.5 |
| total lipids (% wet matter) | 2.2 ± 0.3 | 2.5 ± 0.2 | 2.6 ± 0.2 | 2.9 ± 0.6 |
| Free glucose (μM g−1 wet matter) | 3.33 ± 0.34 | 3.12 ± 0.18 | 3.89 ± 0.40 | 3.32 ± 0.48 |
| Lactate (mM g−1 wet matter) | 3.03 ± 0.11 | 3.41 ± 0.48 | 2.95 ± 0.22 | 2.94 ± 0.31 |
| Ascorbate (μM g−1 wet matter) | 9.7 ± 0.3 | 9.3 ± 0.5 | 9.1 ± 0.7 | 10.2 ± 0.7 |
Total protein content in all carp used in the present study was slightly lower in most cases compared to previously reported values in other studies [
33,
49]. From this, it may be assumed that in general, the nutritional status of the fish was sufficient, since serum total protein levels are known to reflect the nutritional condition of carp [
29]. The albumin concentration in the control group was comparable to previously reported values [
33]. Although the total protein content of serum remained unchanged by DON feeding, albumin concentrations were significantly reduced in fish fed the medium dose and high dose diets for four weeks. This means that the ratio of albumin to total proteins in control fish of more than 80% is reduced in DON-treated fish to 66%, 21% and 31% in fish treated with the low-dose, medium-dose and high-dose diet, respectively. Which adaptations led to the maintenance of total protein levels, although albumin levels were reduced by DON treatment, remains unknown and should be investigated in future studies. Nevertheless, the effect on serum albumin levels was certainly caused by liver impairment that occurred upon DON treatment. That trichothecenes, including DON, are ribotoxic, targeting the 60S ribosomal subunit, and consequently, impairing protein synthesis and transcription, which leads to apoptosis, has already been reported for leukocytes and other actively proliferating eukaryotic cells of higher vertebrates [
51].
In contrast to their activity in liver tissue (
Table 10), the activities of AST and ALT in serum were significantly reduced by DON feeding in the medium-dose group and the high-dose group, respectively, compared to control fish (
Table 11), and only ALT activity was increased in fish recovering from receiving the high-dose DON diet. This indicates that the rate of amino acid transformation via transamination is slowed down by DON. Lactate dehydrogenase (LDH) activity in serum was found to be increased in fish fed the high-dose feed for four weeks and after two weeks of recovery compared to control fish. Increased LDH in serum of carp, indicating membrane leakage in tissues, has also been shown after exposure to toxic concentrations of pesticides [
52]. This parallels the increased lactate levels in DON-treated fish. SDH activity was increased in these fish compared to fish fed the low-dose DON diet. Significant increases of AST, ALT and LDH activities have been observed in carp that have been exposed to handling stress, regular exercise or toxic substances, such as microcystins and cyanide [
33,
53,
54,
55,
56]. Although correlations between the toxin concentrations in feed and AST activities in serum were not significant in the present study during the feeding period, our results showed a significant correlation of ALT, LDH or SDH with the toxin concentrations in the experimental diets (Spearman correlation coefficient, −0.527, 0.466 and 0.475; significance,
p = 0.012, 0.022 and 0.025, respectively). Moreover, after the same time period, albumin or lactate concentrations in serum correlated with the toxin concentration in the experimental feeds (Spearman correlation coefficient, −0.678 and 0.453; significance,
p = 0.000 and 0.034, respectively). Similar results have also been obtained for the recovery phase for the correlation of ALT and LDH with the toxin concentrations previously applied (Spearman correlation coefficient, 0.433 and 0.555; significance,
p = 0.034 and 0.005, respectively). Furthermore, glucose or lactate concentrations in serum correlated with the toxin concentration in the experimental feeds (Spearman correlation coefficient, 0.592 and 0.487; significance,
p = 0.002 and 0.021, respectively). Thus, the activity of serum ALT and LDH together with lactate concentrations seems to be a sensitive indicator of the fish responses to DON. The symptoms of carp exposed to DON resemble the situation of freshwater snakehead fish,
Channa punctatus, that were treated with sublethal concentrations of a carbamate pesticide, leading to increases of LDH and decreased SDH activity in several organs and hyperglycaemia and hyperlactaemia, which suggested that anaerobic metabolism was favored [
57].



{kind=link}
{kind=link}
{kind=link}


