In-vitro Neurotoxicity of Two Malaysian Krait Species (Bungarus candidus and Bungarus fasciatus) Venoms: Neutralization by Monovalent and Polyvalent Antivenoms from Thailand

 and
and 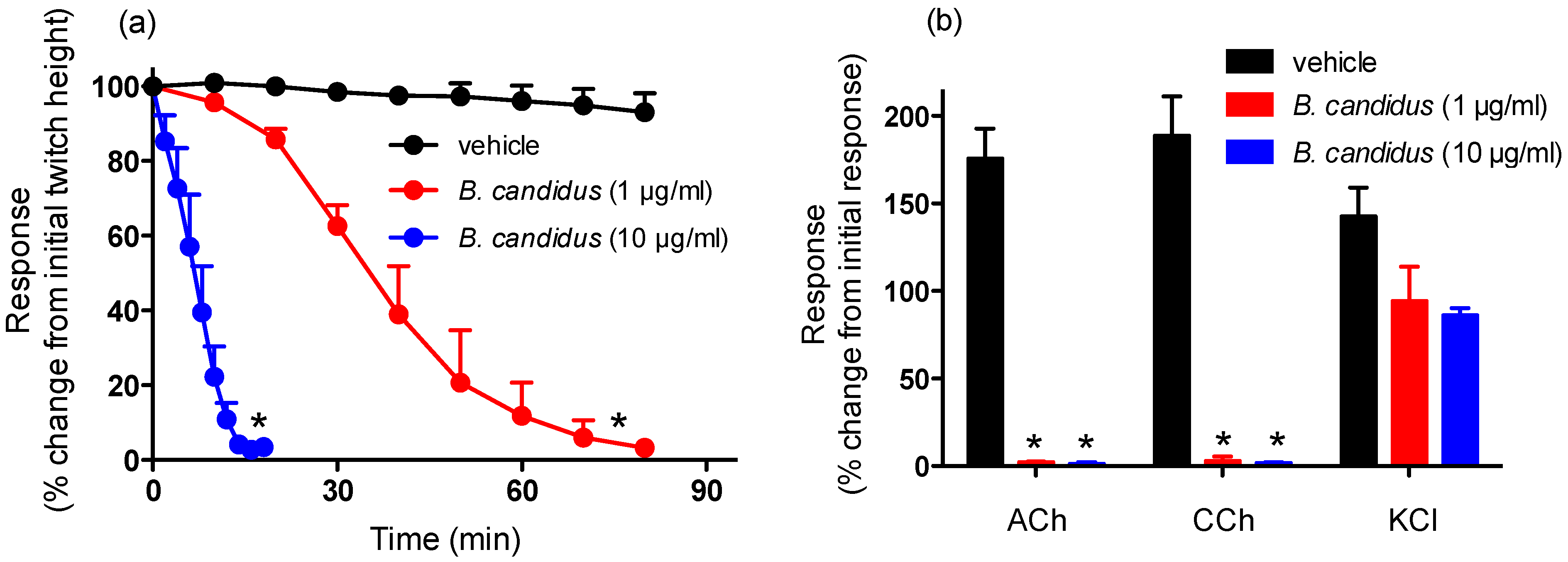{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
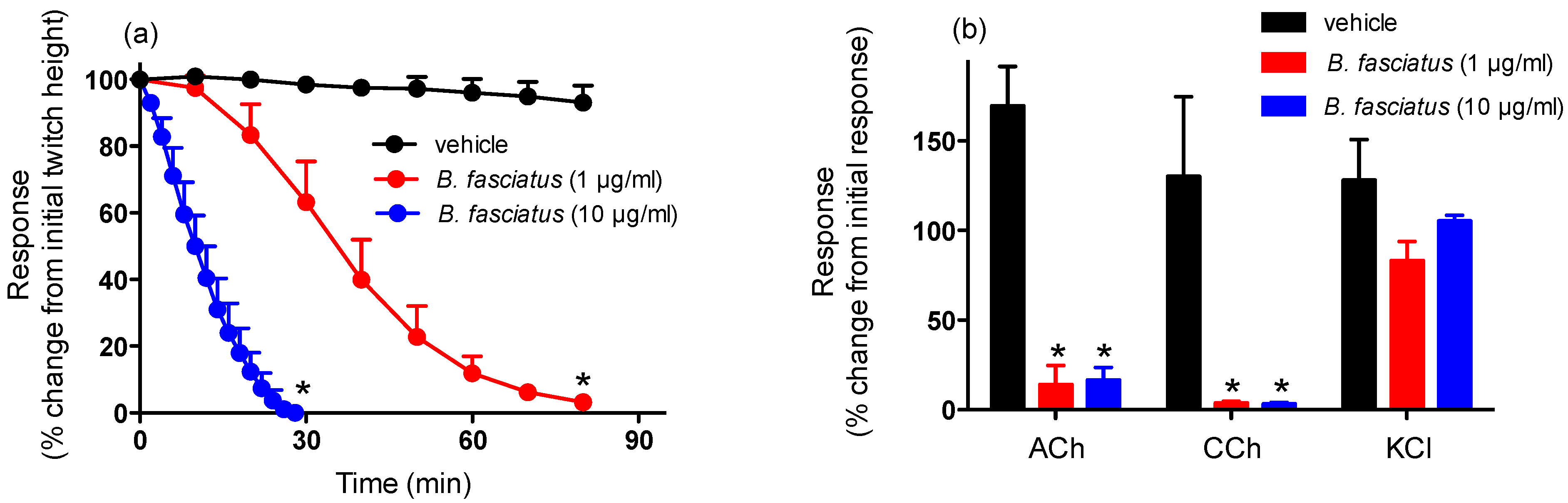
2.1. Chick Biventer Cercivis Nerve-Muscle Preparation


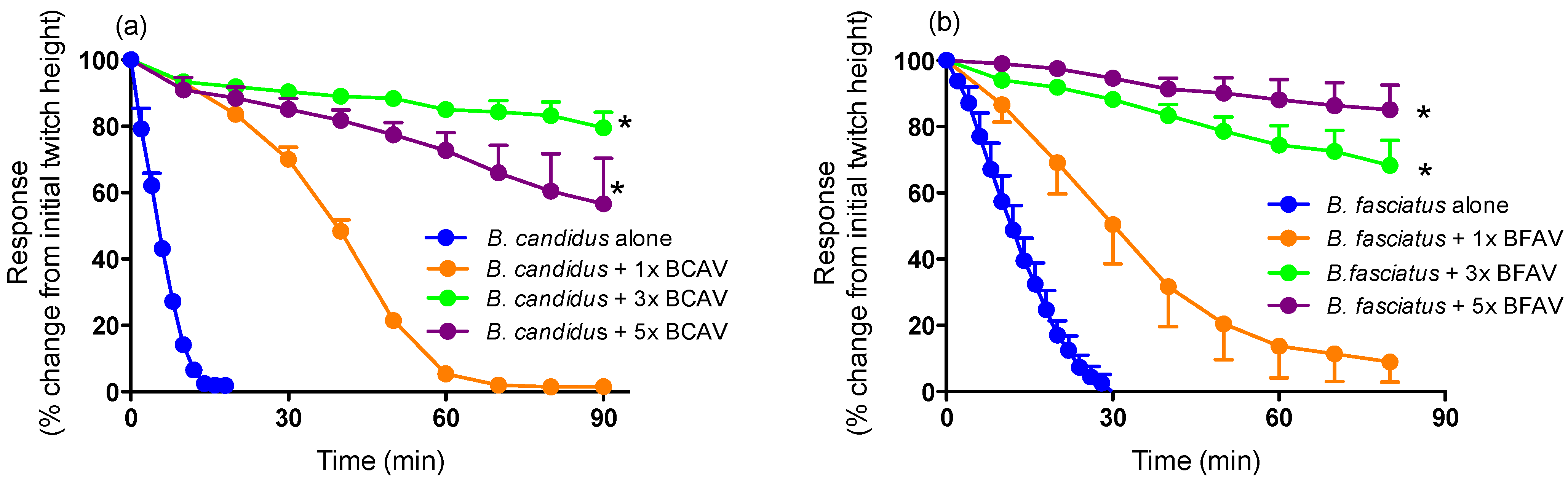
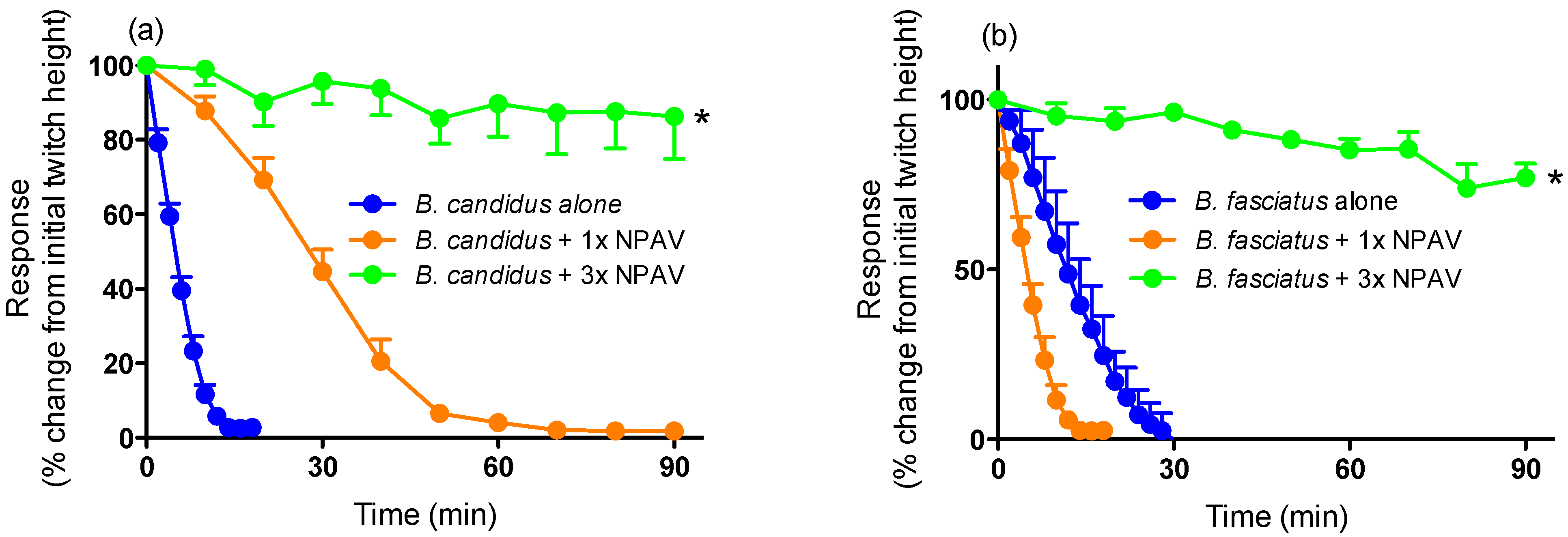
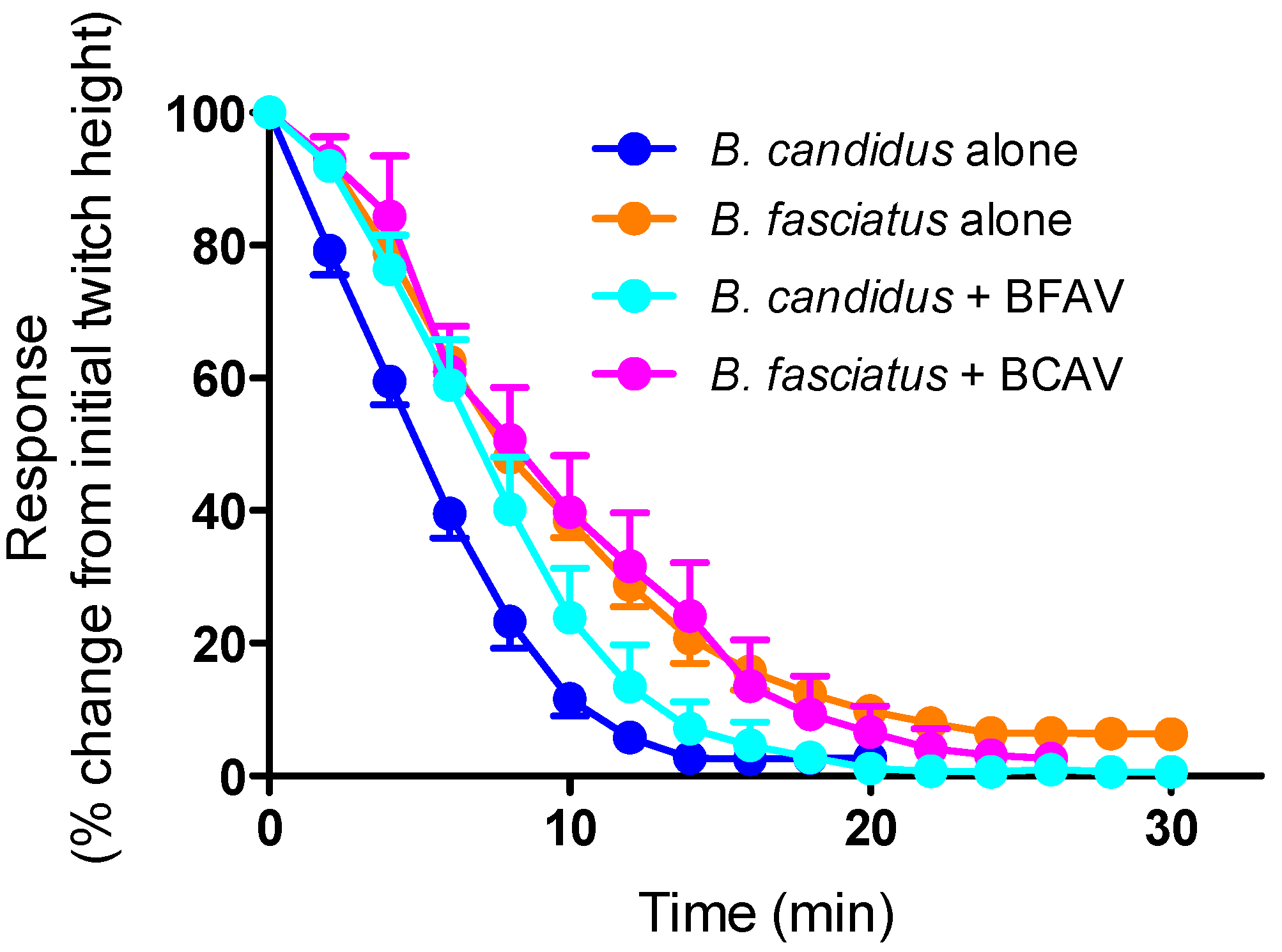
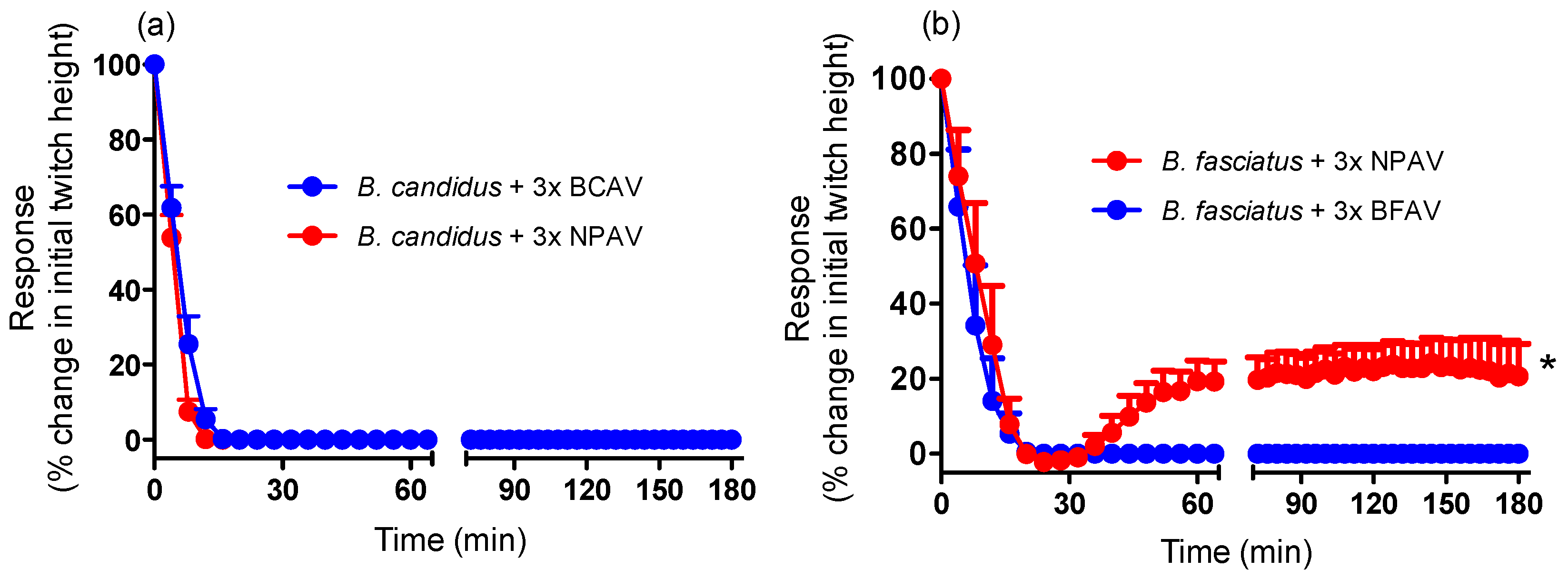
2.2. Antivenom Studies




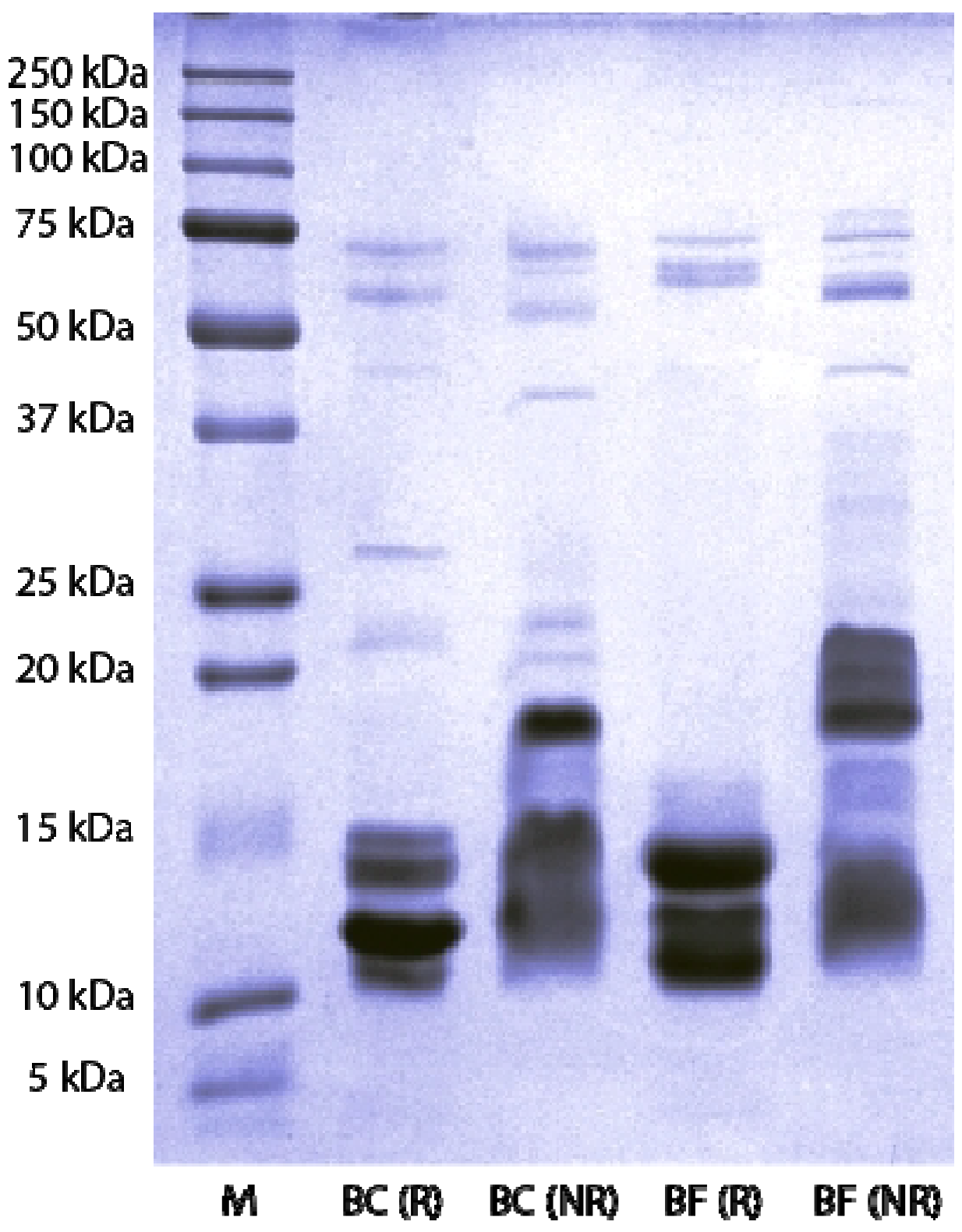
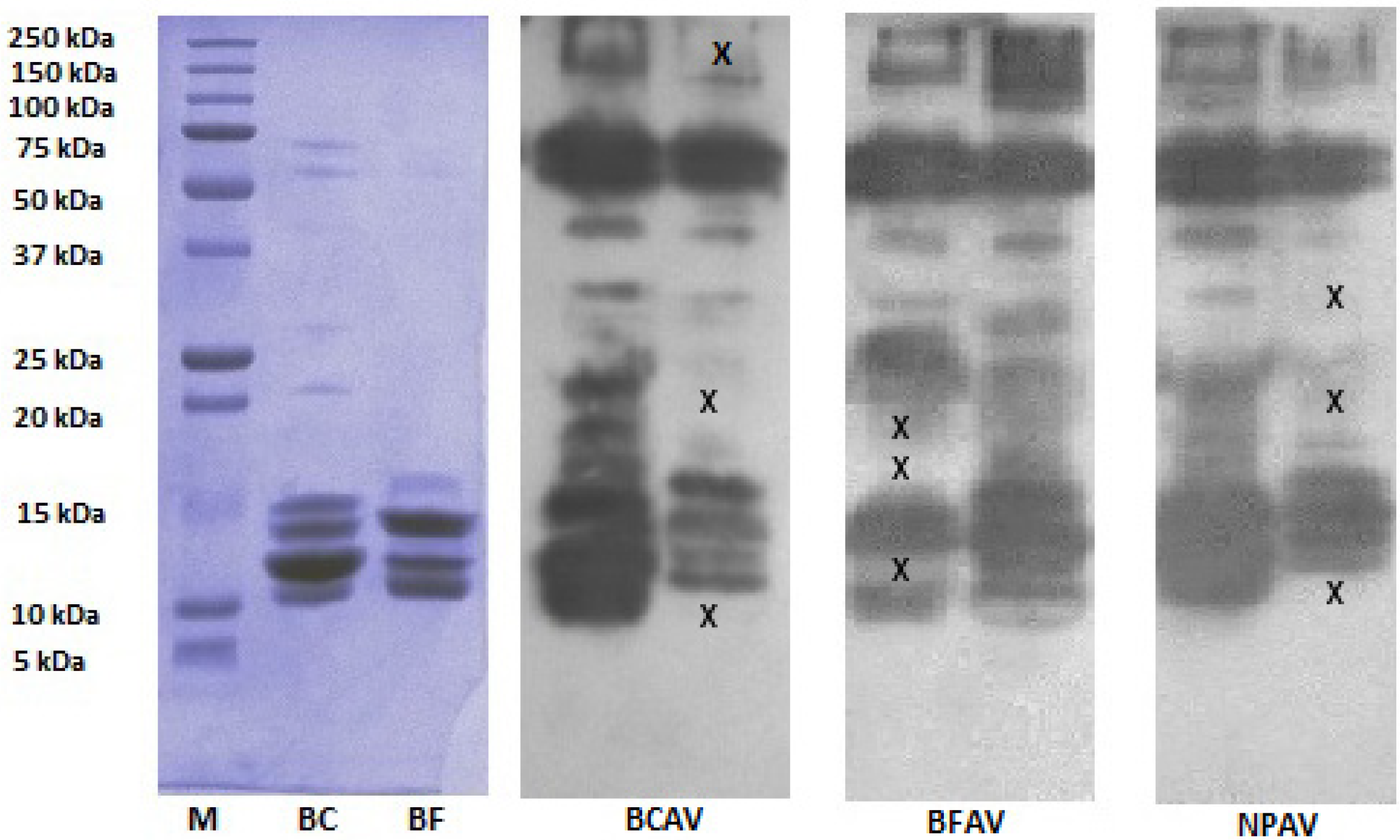
2.3. SDS-PAGE and Western Blot


3. Discussion
4. Experimental Section
4.1. Venoms and Antivenoms
4.2. Chemicals and Drugs
4.3. Protein Quantification by Bicinchoninic Acid Assay (BCA)
4.4. Indirectly-Stimulated Chick Biventer Cervicis Nerve-Muscle Preparation (CBCM)
4.5. Sodium Dodecyl Sulphate Polyacylamide Gel Electrophoresis (SDS-PAGE)
4.6. Western-Blot
4.7. Data Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Chanhome, L.; Cox, M.J.; Wilde, H.; Jintakoon, P.; Chaiyabutr, N.; Sitprija, V. Venomous snakebite in Thailand. I: Medically important snakes. Mil. Med. 1998, 163, 310–317. [Google Scholar]
- Kuch, U. Range extension and first record for Indonesia of the Kinabalu krait, Bungarus flaviceps baluensis LOVERIDGE, 1938. Herpatozoa 2002, 14, 149–151. [Google Scholar]
- Hojer, J.; Tran Hung, H.; Warrell, D. Life-threatening hyponatremia after krait bite envenoming—A new syndrome. Clin. Toxicol. 2010, 48, 956–957. [Google Scholar] [CrossRef]
- Laothong, C.; Sitprija, V. Decreased parasympathetic activities in Malayan krait (Bungarus candidus) envenoming. Toxicon 2001, 39, 1353–1357. [Google Scholar] [CrossRef]
- Pochanugool, C.; Wilde, H.; Jitapunkul, S.; Limthongkul, S. Spontaneous recovery from severe neurotoxic envenoming by a Malayan krait Bungarus candidus (Linnaeus) in Thailand. Wilderness Environ. Med. 1997, 8, 223–225. [Google Scholar] [CrossRef]
- Pe, T.; Myint, T.; Htut, A.; Htut, T.; Myint, A.A.; Aung, N.N. Envenoming by Chinese krait (Bungarus multicinctus) and banded krait (B. fasciatus) in Myanmar. Trans. R. Soc. Trop. Med. Hyg. 1997, 91, 686–688. [Google Scholar] [CrossRef]
- Tsai, I.H.; Tsai, H.Y.; Saha, A.; Gomes, A. Sequences, geographic variations and molecular phylogeny of venom phospholipases and threefinger toxins of eastern India Bungarus fasciatus and kinetic analyses of its Pro31 phospholipases A2. FEBS J. 2007, 274, 512–525. [Google Scholar] [CrossRef]
- Ariaratnam, C.A.; Sheriff, M.R.; Theakston, R.D.G.; Warrell, D.A. Distinctive epidemiologic and clinical features of common krait (Bungarus caeruleus) bites in sri lanka. Am. J. Trop. Med. Hyg. 2008, 79, 458–462. [Google Scholar]
- Zulkifli, A.; Hashim, M.; Khairul Anuar, A. Snake bites in Kelantan, Peninsular Malaysia. Trop. Biomed. 1995, 12, 1–4. [Google Scholar]
- Chew, K.; Khor, H.; Ahmad, R.; Rahman, N. A five-year retrospective review of snakebite patients admitted to a tertiary university hospital in Malaysia. Int. J. Emerg. Med. 2011, 4, 41. [Google Scholar] [CrossRef]
- Kanchanapongkul, J. Neurotoxic envenoming following bites by the Malayan krait (Bungarus candidus). J. Med. Assoc. Thail. 2002, 85, 945–948. [Google Scholar]
- Kuch, U.; Molles, B.E.; Omori-Satoh, T.; Chanhome, L.; Samejima, Y.; Mebs, D. Identification of alpha-bungarotoxin (A31) as the major postsynaptic neurotoxin, and complete nucleotide identity of a genomic DNA of Bungarus candidus from Java with exons of the Bungarus multicinctus alpha-bungarotoxin (A31) gene. Toxicon 2003, 42, 381–390. [Google Scholar] [CrossRef]
- Khow, O.; Chanhome, L.; Omori-Satoh, T.; Ogawa, Y.; Yanoshita, R.; Samejima, Y.; Kuch, U.; Mebs, D.; Sitprija, V. Isolation, toxicity and amino terminal sequences of three major neurotoxins in the venom of Malayan krait (Bungarus candidus) from Thailand. J. Biochem. 2003, 134, 799–804. [Google Scholar] [CrossRef]
- Tan, N.H.; Ponnudurai, G. A comparative study of the biological properties of krait (genus Bungarus) venoms. Comp. Biochem. Physiol. C. 1990, 95, 105–109. [Google Scholar] [CrossRef]
- Yanoshita, R.; Ogawa, Y.; Murayama, N.; Omori-Satoh, T.; Saguchi, K.; Higuchi, S.; Khow, O.; Chanhome, L.; Samejima, Y.; Sitprija, V. Molecular cloning of the major lethal toxins from two kraits (Bungarus flaviceps and Bungarus candidus). Toxicon 2006, 47, 416–424. [Google Scholar] [CrossRef]
- Awasthi, R.; Narang, S.; Chowdhury, P.P. Cerebellar ataxia following snake bite. JAPI 2010, 58, 389–390. [Google Scholar]
- Chieh-Fan, C.; Tzeng-Jih, L.; Wen-Chi, H.; Hua-Wei, Y. Appropriate antivenom doses for six types of envenomations caused by snakes in taiwan. J. Venom Anim. Toxins 2009, 15, 479–490. [Google Scholar]
- Leeprasert, W.; Kaojarern, S. Specific antivenom for Bungarus candidus. J. Med. Assoc. Thail. 2007, 90, 1467–1476. [Google Scholar]
- Leong, P.K.; Sim, S.M.; Fung, S.Y.; Sumana, K.; Sitprija, V.; Tan, N.H. Cross neutralization of Afro-Asian cobra and Asian krait venoms by a Thai polyvalent snake antivenom (Neuro Polyvalent Snake Antivenom). PLoS Negl. Trop. Dis. 2012, 6, e1672. [Google Scholar] [CrossRef]
- Chanhome, L.; Wongtongkam, N.; Khow, O.; Pakmanee, N.; Omori-Satoh, T.; Sitprija, V. Genus specific neutralization of Bungarus snake venoms by Thai Red Cross banded krait antivenom. J. Nat. Toxins 1999, 8, 135–140. [Google Scholar]
- Ali, S.A.; Yang, D.C.; Jackson, T.N.; Undheim, E.A.; Koludarov, I.; Wood, K.; Jones, A.; Hodgson, W.C.; McCarthy, S.; Ruder, T.; et al. Venom proteomic characterization and relative antivenom neutralization of two medically important Pakistani elapid snakes (Bungarus sindanus and Naja naja). J. Proteomics 2013, 89, 15–23. [Google Scholar] [CrossRef]
- Chotwiwatthanakun, C.; Pratanaphon, R.; Akesowan, S.; Sriprapat, S.; Ratanabanangkoon, K. Production of potent polyvalent antivenom against three elapid venoms using a low dose, low volume, multi-site immunization protocol. Toxicon 2001, 39, 1487–1494. [Google Scholar] [CrossRef]
- Fry, B.G.; Wickramaratna, J.C.; Jones, A.; Alewood, P.F.; Hodgson, W.C. Species and regional variations in the effectiveness of antivenom against the in vitro neurotoxicity of death adder (Acanthophis) venoms. Toxicol. Appl. Pharm. 2001, 175, 140–148. [Google Scholar] [CrossRef]
- Fry, B.G.; Winkel, K.D.; Wickramaratna, J.C.; Hodgson, W.C.; Wüster, W. Effectiveness of snake antivenom: Species and regional venom variation and its clinical impact. Toxin Rev. 2003, 22, 23–34. [Google Scholar] [CrossRef]
- Shashidharamurthy, R.; Jagadeesha, D.; Girish, K.; Kemparaju, K. Variation in biochemical and pharmacological properties of Indian cobra (Naja naja naja) venom due to geographical distribution. Mol. Cell. Biochem. 2002, 229, 93–101. [Google Scholar] [CrossRef]
- Skejić, J.; Hodgson, W.C. Population divergence in venom bioactivities of elapid snake Pseudonaja textilis: Role of procoagulant proteins in rapid rodent prey incapacitation. PLoS ONE 2013, 8, e63988. [Google Scholar]
- Chang, H.C.; Tsai, T.S.; Tsai, I.H. Functional proteomic approach to discover geographic variations of king cobra venoms from Southeast Asia and China. J. Proteomics 2013, 89, 141–153. [Google Scholar] [CrossRef]
- Hodgson, W.C.; Wickramaratna, J.C. In vitro neuromuscular activity of snake venoms. Clin. Exp. Pharmacol. Physiol. 2002, 29, 807–814. [Google Scholar] [CrossRef]
- Karsani, S.A.; Othman, I. Isolation, complete amino acid sequence and characterization of a previously unreported post-synaptic neurotoxin—AlphaN3, from the venom of Bungarus candidus. Biochem. Biophys. Res. Commun. 2009, 389, 343–348. [Google Scholar] [CrossRef]
- Bon, C.; Saliou, B. Ceruleotoxin: Identification in the venom of Bungarus fasciatus, molecular properties and importance of phospholipase A2 activity for neurotoxicity. Toxicon 1983, 21, 681–698. [Google Scholar] [CrossRef]
- Malih, I.; Ahmad Rusmili, M.R.; Tee, T.Y.; Saile, R.; Ghalim, N.; Othman, I. Proteomic analysis of Moroccan cobra Naja haje legionis venom using tandem mass spectrometry. J. Proteomics 2014, 96, 240–252. [Google Scholar] [CrossRef]
- Chanhome, L.; Khow, O.; Puempunpanich, S.; Sitprija, V.; Chaiyabutr, N. Biological characteristics of the Bungarus candidus venom due to geographical variation. J. Cell Anim. Biol. 2009, 3, 93–100. [Google Scholar]
- Chanhome, L.; Puempunpanich, S.; Omori-Satoh, T.; Chaiyabutr, N.; Sitprija, V. A pilot experiment for production of malayan krait antivenom: Immunization of rabbits with Bungarus candidus venom. J. Nat. Toxins 2002, 11, 353–356. [Google Scholar]
- Prasarnpun, S.; Walsh, J.; Awad, S.; Harris, J. Envenoming bites by kraits: The biological basis of treatment-resistant neuromuscular paralysis. Brain 2005, 128, 2987–2996. [Google Scholar] [CrossRef]
- Kornhauser, R.; Isbister, G.K.; O’Leary, M.A.; Mirtschin, P.; Dunstan, N.; Hodgson, W.C. Cross-neutralisation of the neurotoxic effects of Egyptian cobra venom with commercial tiger snake antivenom. Basic Clin. Pharmacol. Toxicol. 2013, 112, 138–143. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Lomonte, B.; Leon, G.; Alape-Giron, A.; Flores-Diaz, M.; Sanz, L.; Angulo, Y.; Calvete, J.J. Snake venomics and antivenomics: Proteomic tools in the design and control of antivenoms for the treatment of snakebite envenoming. J. Proteomics 2009, 72, 165–182. [Google Scholar] [CrossRef]
- Kini, R.M.; Doley, R. Structure, function and evolution of three-finger toxins: Mini proteins with multiple targets. Toxicon 2010, 56, 855–867. [Google Scholar] [CrossRef]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rusmili, M.R.A.; Yee, T.T.; Mustafa, M.R.; Othman, I.; Hodgson, W.C. In-vitro Neurotoxicity of Two Malaysian Krait Species (Bungarus candidus and Bungarus fasciatus) Venoms: Neutralization by Monovalent and Polyvalent Antivenoms from Thailand. Toxins 2014, 6, 1036-1048. https://doi.org/10.3390/toxins6031036
Rusmili MRA, Yee TT, Mustafa MR, Othman I, Hodgson WC. In-vitro Neurotoxicity of Two Malaysian Krait Species (Bungarus candidus and Bungarus fasciatus) Venoms: Neutralization by Monovalent and Polyvalent Antivenoms from Thailand. Toxins. 2014; 6(3):1036-1048. https://doi.org/10.3390/toxins6031036
Chicago/Turabian StyleRusmili, Muhamad Rusdi Ahmad, Tee Ting Yee, Mohd Rais Mustafa, Iekhsan Othman, and Wayne C. Hodgson. 2014. "In-vitro Neurotoxicity of Two Malaysian Krait Species (Bungarus candidus and Bungarus fasciatus) Venoms: Neutralization by Monovalent and Polyvalent Antivenoms from Thailand" Toxins 6, no. 3: 1036-1048. https://doi.org/10.3390/toxins6031036
APA StyleRusmili, M. R. A., Yee, T. T., Mustafa, M. R., Othman, I., & Hodgson, W. C. (2014). In-vitro Neurotoxicity of Two Malaysian Krait Species (Bungarus candidus and Bungarus fasciatus) Venoms: Neutralization by Monovalent and Polyvalent Antivenoms from Thailand. Toxins, 6(3), 1036-1048. https://doi.org/10.3390/toxins6031036