
Nephroprotective Role of Resveratrol and Ursolic Acid in Aristolochic Acid Intoxicated Zebrafish

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
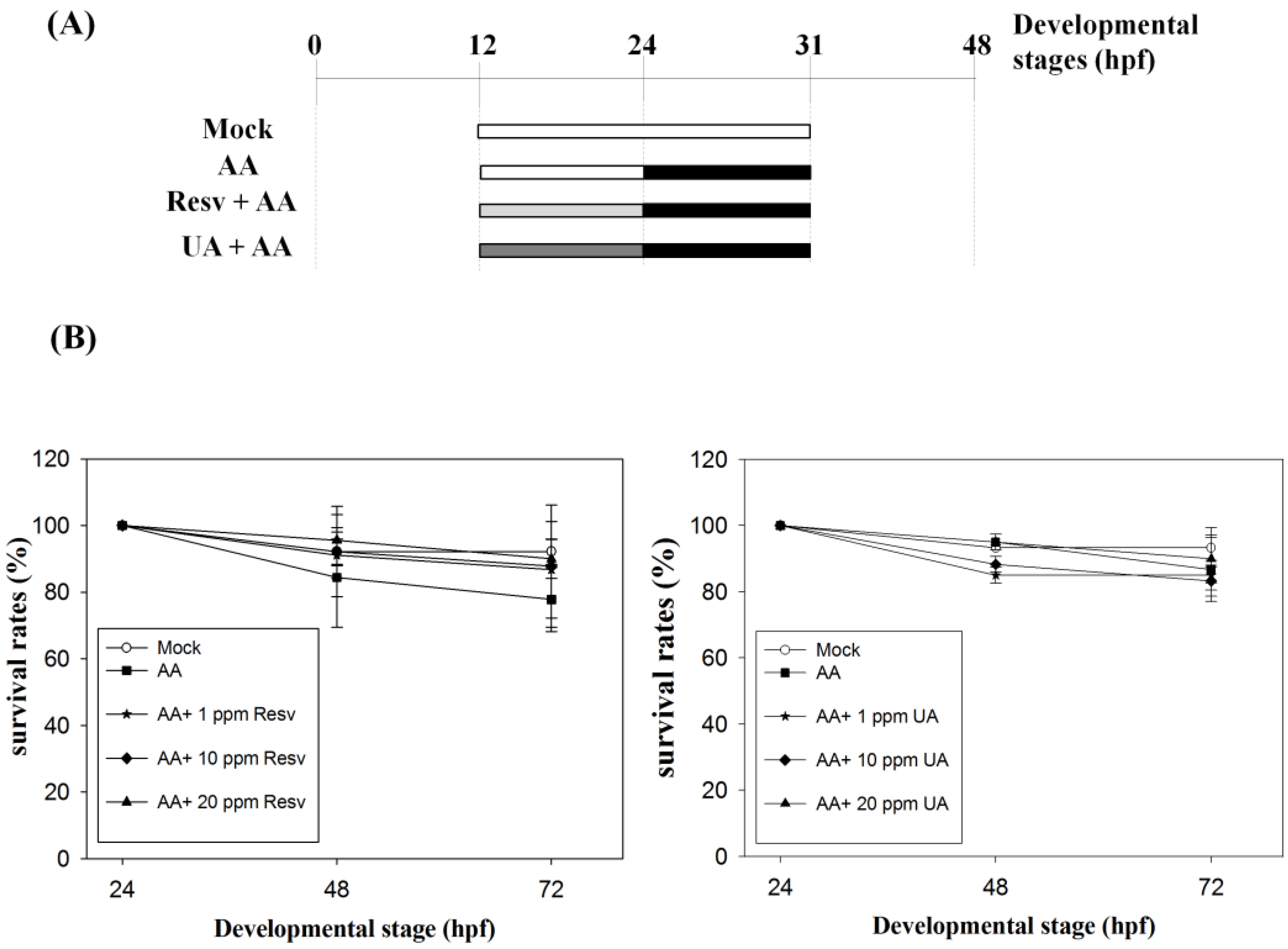
2.1. Resv/UA Has No Evident Effects for Enhancing Survival Rates of AA-Intoxicated Zebrafish Embryos

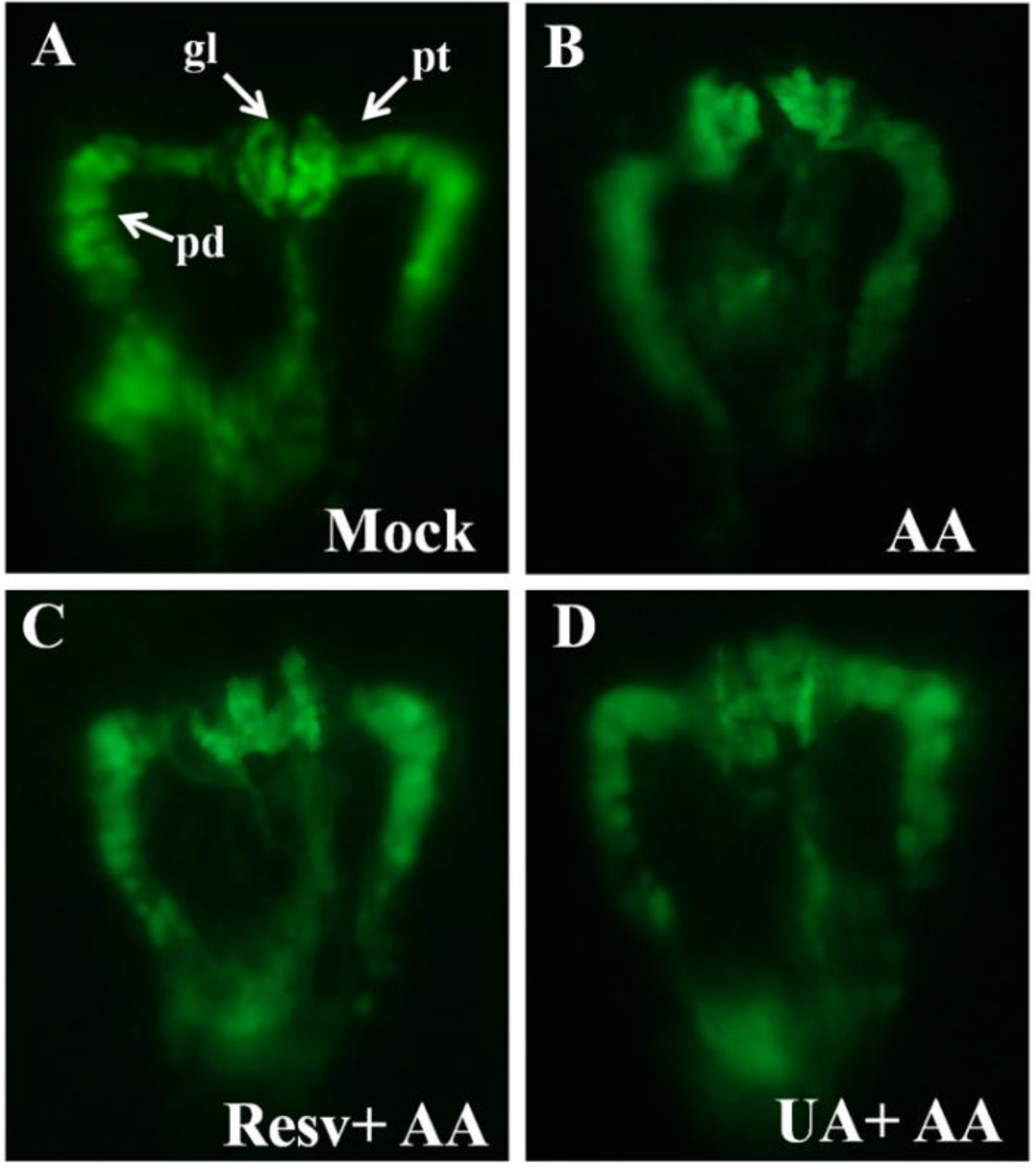
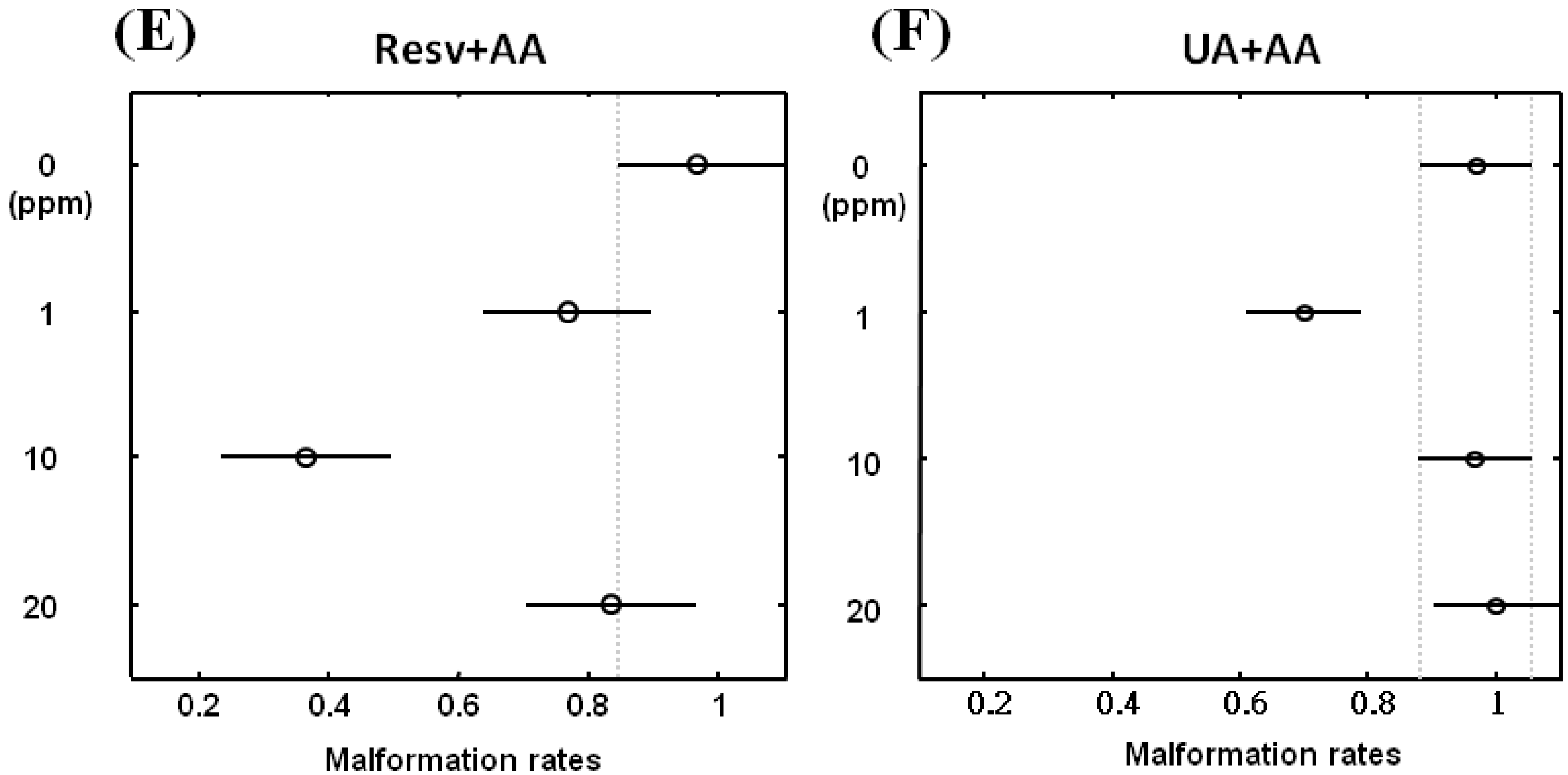
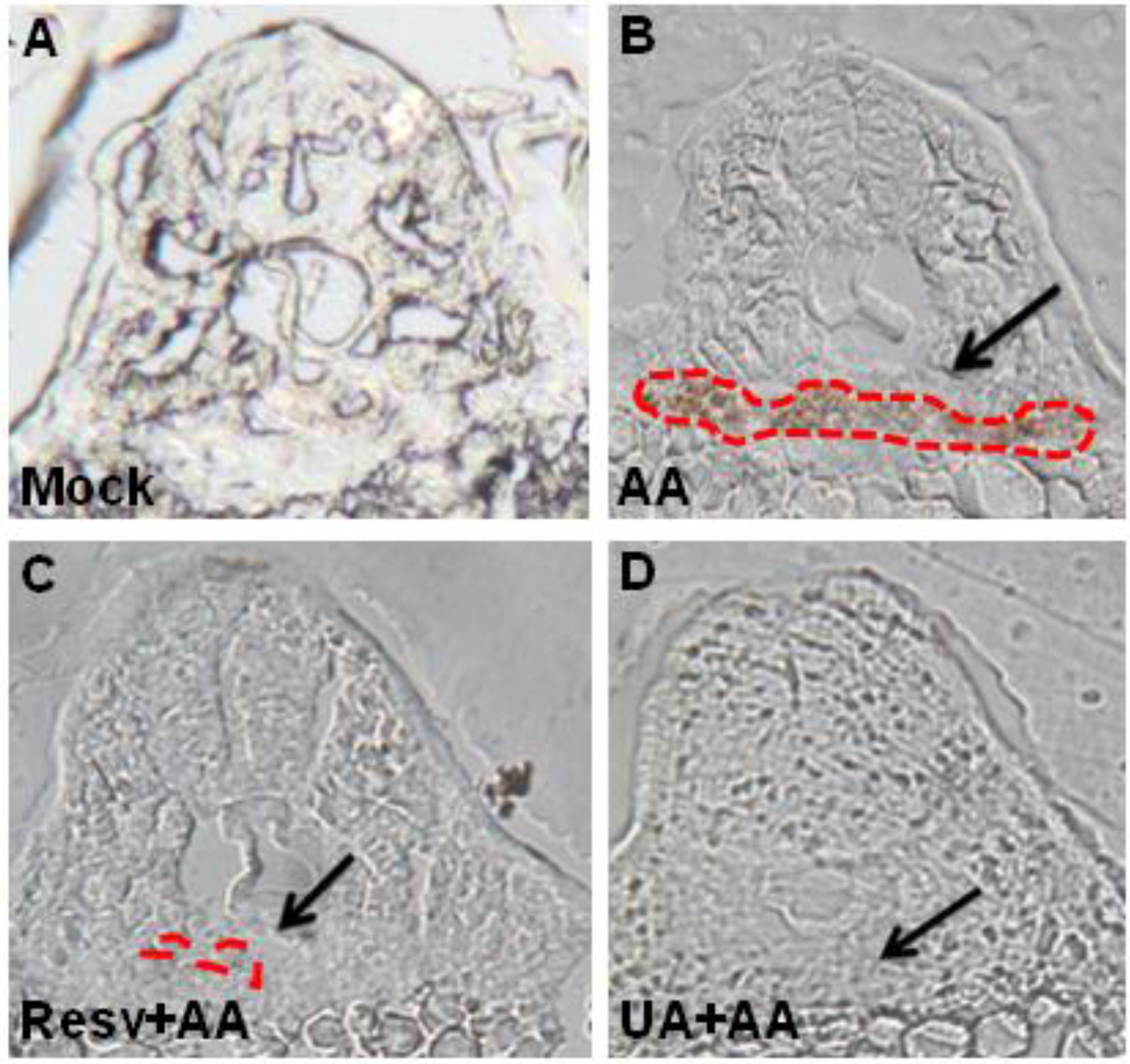
2.2. Resv and UA Can Attenuate Aristolochic Acid-Induced Kidney Malformation


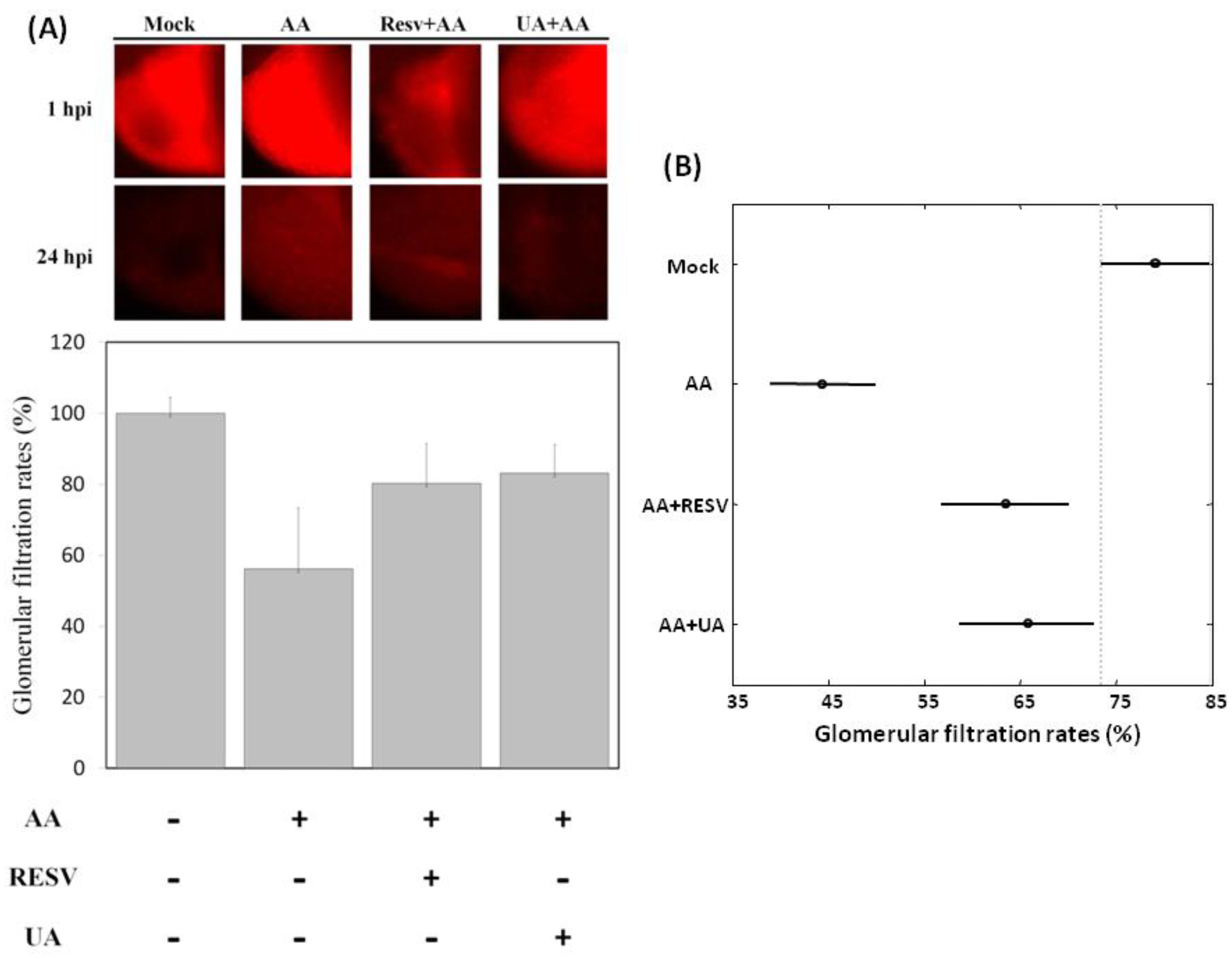
2.3. Resv and UA Treatment Attenuate AA-Induced Renal Failure

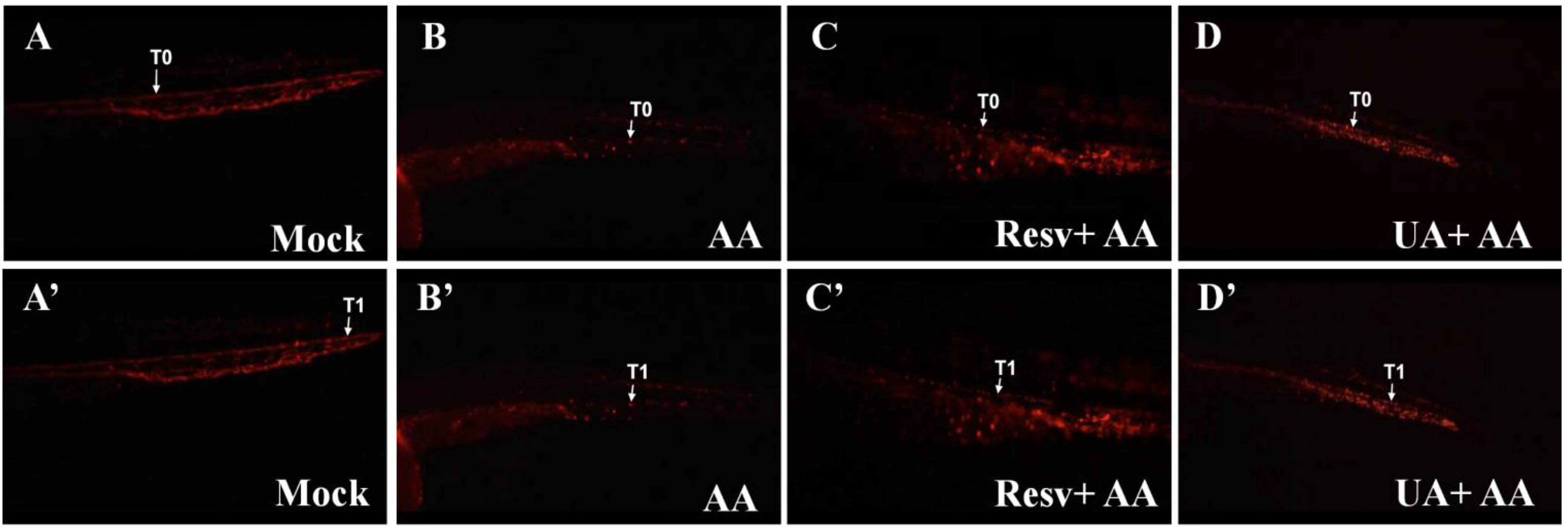
2.4. Resv and UA Improve the Accumulation of Erythrocytes and Restoration of Blood Circulation


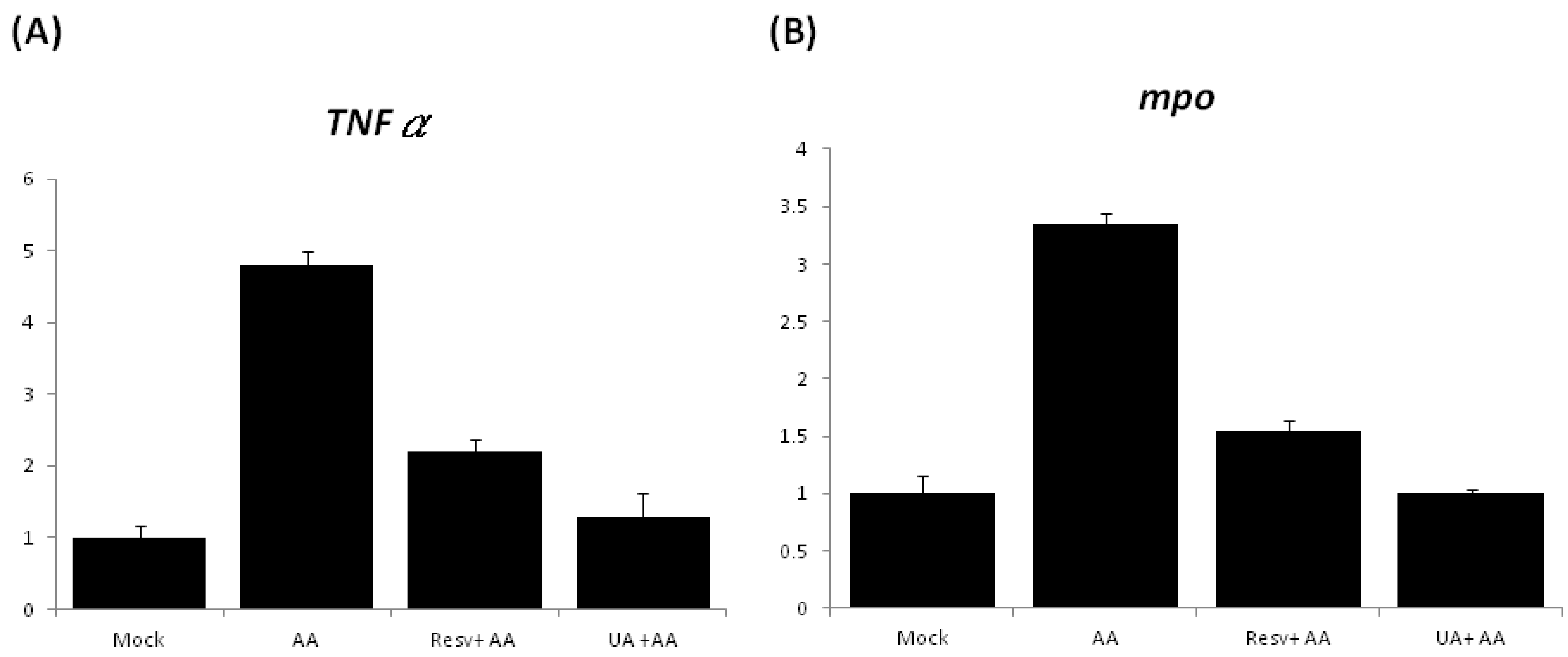
2.5. Treatment with Either Resv or UA Suppresses the Generation of ROS and the Inflammatory Response


3. Experimental Section
3.1. Fish Maintenance and Embryo Collection
3.2. Chemical Exposure and Survival Rate Analysis
3.3. Detection of ROS and Data Analysis
3.4. Zebrafish Renal Function Assay
3.5. Red Blood Cell Staining
3.6. Histology and Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
3.7. Images
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Murphy, T.; Robinson, S. Renal failure and its treatment. Anaesthesia Intensiv. Care Med. 2006, 7, 7. [Google Scholar] [CrossRef]
- Naughton, C.A. Drug-induced nephrotoxicity. Am. Fam. Physician 2008, 78, 743–750. [Google Scholar] [PubMed]
- Perazella, M.A. Renal vulnerability to drug toxicity. Clin. J. Am. Soc. Nephrol. 2009, 4, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, Y.; Kinoshita, K.; Yano, T.; Asato, K.; Shiga, T.; Hino, S.; Niki, K.; Nagare, Y.; Kishimoto, K.; Shimazu, H.; et al. Signaling through the interleukin-18 receptor α attenuates inflammation in cisplatin-induced acute kidney injury. Kidney Int. 2012, 82, 892–902. [Google Scholar] [CrossRef] [PubMed]
- Arlt, V.M.; Ferluga, D.; Stiborova, M.; Pfohl-Leszkowicz, A.; Vukelic, M.; Ceovic, S.; Schmeiser, H.H.; Cosyns, J.P. Is aristolochic acid a risk factor for Balkan endemic nephropathy-associated urothelial cancer? Int. J. Cancer 2002, 101, 500–502. [Google Scholar] [CrossRef] [PubMed]
- Debelle, F.D.; Vanherweghem, J.L.; Nortier, J.L. Aristolochic acid nephropathy: A worldwide problem. Kidney Int. 2008, 74, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Gökmen, R.; Cosyns, J.P.; Arlt, V.M.; Stiborová, M.; Phillips, D.H.; Schmeiser, H.H.; Simmonds, M.S.J.; Cook, H.T.; Vanherweghem, J.L.; Nortier, J.L.; et al. The epidemiology, diagnosis, and management of aristolochic acid nephropathy: A narrative review. Ann. Intern. Med. 2013, 158, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Grollman, A.P.; Jelaković, B. Role of environmental toxins in endemic (Balkan) nephropathy. J. Am. Soc. Nephrol. 2007, 18, 2817–2823. [Google Scholar] [CrossRef] [PubMed]
- Grollman, A.P. Aristolochic acid nephropathy: Harbinber of a global iatrogenic disease. Environ. Mol. Mutagen. 2013, 54, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Depierreux, M.; van Damme, B.; Vanden Houte, K.; Vanherweghem, J.L. Pathologic aspects of a newly described nephropathy related to the prolonged use of Chinese herbs. Am. J. Kidney Dis. 1994, 24, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Pozdzik, A.A.; Salmon, I.J.; Husson, C.P.; Decaestecker, C.; Rogier, E.; Bourgeade, M.F.; Deschodt-Lanckman, M.M.; Vanherweghem, J.L.; Nortier, J.L. Patterns of interstitial inflammation during the evolution of renal injury in experimental aristolochic acid nephropathy. Nephrol. Dial. Transplant. 2008, 23, 2480–2491. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.J.; Chen, Y.H. Developmental nephrotoxicity of aristolochic acid in a zebrafish model. Toxicol. Appl. Pharmacol. 2012, 261, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Csaki, C.; Nebrich, S.; Mobasheri, A. Resveratrol suppresses interleukin-1β-induced inflammatory signaling and apoptosis in human articular chondrocytes: Potential for use as a novel nutraceutical for the treatment of osteoarthritis. Biochem. Pharmacol. 2008, 76, 1426–1439. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Murakami, A.; Ohigashi, H. Ursolic acid: An anti- and pro-inflammatory triterpenoid. Mol. Nutr. Food Res. 2008, 52, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, D.; Wang, S.; Ha, S.; Li, P. Oleanolic acid in decreasing hyperglycemia. Chin. Pharmaceut. J. 1994, 29, 725–726. [Google Scholar]
- Singh, G.B.; Singh, S.; Bani, S.; Gupta, B.D.; Banerjee, S.K. Anti-inflammatory activity of oleanolic acid in rats and mice. J. Pharm. Pharmacol. 1992, 44, 456–458. [Google Scholar] [CrossRef] [PubMed]
- Recio, M.C.; Giner, R.M.; Manez, S.; Gueho, J.; Julien, H.R.; Hostettmann, K.; Rios, J.L. Investigations on the steroidal anti-inflammatory activity of triterpenoids from Diospyros leucomelas. Planta. Med. 1995, 61, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Castro, R.; Lamas, J.; Morais, P.; Sanmartín, M.L.; Orallo, F.; Leiro, J. Resveratrol modulates innate and inflammatory responses in fish leucocytes. Vet. Immunol. Immunopathol. 2008, 126, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Švajger, U.; Jeras, M. Anti-inflammatory effects of resveratrol and its potential use in therapy of immune-mediated diseases. Int. Rev. Immunol. 2012, 31, 202–222. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Hernández, M.; Córdova, C.; Nieto, M.L. Natural triterpenes modulate immune-inflammatory markers of experimental autoimmune encephalomyelitis: Therapeutic implications for multiple sclerosis. Br. J. Pharmacol. 2012, 166, 1708–1723. [Google Scholar] [CrossRef] [PubMed]
- Spitsbergen, J.M.; Kent, M.L. The state of the art of the zebrafish model for toxicology and toxicologic pathology research- advantages and current limitations. Toxicol. Pathol. 2003, 31, 62–87. [Google Scholar] [PubMed]
- Stern, H.M.; Zon, L.I. Cancer genetics and drug discovery in the zebrafish. Nature Rev. 2003, 3, 1–7. [Google Scholar]
- Chen, Y.H.; Yang, Z.S.; Wen, C.C.; Chang, Y.S.; Wang, P.C.; Hsiao, C.A.; Shih, T.L. Evaluation of the structure-activity relationship of flavonoids as antioxidants and toxicants of zebrafish larvae. Food Chem. 2012, 134, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Wang, W.H.; Wang, Y.H.; Lin, Z.Y.; Wen, C.C.; Chern, C.Y. Evaluation of anti-inflammatory effect of chalcone and chalcone analogues in a zebrafish model. Molecules 2013, 18, 2052–2060. [Google Scholar] [CrossRef] [PubMed]
- Perner, B.; Englert, C.; Bollig, F. The Wilms tumor genes wt1a and wt1b control different steps during formation of the zebrafish pronephros. Dev. Biol. 2007, 309, 7–96. [Google Scholar] [CrossRef]
- Westhoff, J.H.; Giselbrecht, S.; Schmidts, M.; Schindler, S.; Beales, P.L.; Tönshoff, B.; Liebel, U.; Gehrig, J. Development of an automated imaging pipeline for the analysis of the zebrafish larval kidney. PLoS One 2013, 8, e82137. [Google Scholar] [CrossRef] [PubMed]
- Traver, D.; Paw, B.H.; Poss, K.D.; Penberthy, W.T.; Lin, S.; Zon, L.I. Transplantation and in vivo imaging of multilineage engraftment in zebrafish bloodless mutants. Nat. Immunol. 2003, 4, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Chen, P.C.; Huang, C.W.; Yu, J. Aristolochic acid induces heart failure in zebrafish embryos that is mediated by inflammation. Toxicol. Sci. 2007, 100, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Yaqoob, N.; Holotta, M.; Prem, C.; Kopp, R.; Schwerte, T. Ontogenetic development of erythropoiesis can be studied non-invasively in GATA-1: DsRed transgenic zebrafish. Comp. Biochem. Physiol. A 2009, 154, 270–278. [Google Scholar] [CrossRef]
- Kimmel, C.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development in the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, D.M.; Park, K.M.; Cilenti, L.; Zervos, A.S.; Drummond, I.; Bonventre, J.V. Acute renal failure in zebrafish: A novel system to study a complex disease. Am. J. Physiol. Renal Physiol. 2005, 288, F923–F929. [Google Scholar] [CrossRef] [PubMed]
- Detrich, H.W., 3rd; Kieran, M.W.; Chan, F.Y.; Barone, L.M.; Yee, K.; Rundstadler, J.A.; Pratt, S.; Ransom, D.; Zon, L.I. Intraembryonic hematopoietic cell migration during vertebrate development. Proc. Natl. Acad. Sci. USA 1995, 92, 10713–10717. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Cheng, C.C.; Lee, W.J.; Chiou, M.L.; Pai, C.W.; Wen, C.C.; Chen, W.L.; Chen, Y.H. A novel phenotype-based approach for systematically screening antiproliferation metallodrugs. Chem.-Biol. Interact. 2009, 182, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H.; Chang, M.Y.; Hsu, C.H.; Chen, Y.H. Essential roles of basic helix-loop-helix transcription factors, Capsulin and Musculin, during craniofacial myogenesis of zebrafish. Cell. Mol. Life Sci. 2011, 68, 4065–4078. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.H.; Liao, Y.F.; Chang, C.Y.; Tsai, J.N.; Wang, Y.H.; Cheng, C.C.; Wen, C.C.; Chen, Y.H. Caffeine treatment disturbs the angiogenesis of zebrafish larvae. Drug Chem. Toxicol. 2012, 35, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Schrier, R.W.; Wang, W.; Poole, B.; Mitra, A. Acute renal failure: defi nitions, diagnosis, pathogenesis, and therapy. J. Clin. Invest. 2004, 114, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Kinsey, G.R.; Okusa, M.D. Role of leukocytes in the pathogenesis of acute kidney injury. Crit. Care 2012, 16, 214. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Y.-J.; Sun, C.-Y.; Wen, C.-C.; Chen, Y.-H. Nephroprotective Role of Resveratrol and Ursolic Acid in Aristolochic Acid Intoxicated Zebrafish. Toxins 2015, 7, 97-109. https://doi.org/10.3390/toxins7010097
Ding Y-J, Sun C-Y, Wen C-C, Chen Y-H. Nephroprotective Role of Resveratrol and Ursolic Acid in Aristolochic Acid Intoxicated Zebrafish. Toxins. 2015; 7(1):97-109. https://doi.org/10.3390/toxins7010097
Chicago/Turabian StyleDing, Yu-Ju, Chiao-Yin Sun, Chi-Chung Wen, and Yau-Hung Chen. 2015. "Nephroprotective Role of Resveratrol and Ursolic Acid in Aristolochic Acid Intoxicated Zebrafish" Toxins 7, no. 1: 97-109. https://doi.org/10.3390/toxins7010097
APA StyleDing, Y.-J., Sun, C.-Y., Wen, C.-C., & Chen, Y.-H. (2015). Nephroprotective Role of Resveratrol and Ursolic Acid in Aristolochic Acid Intoxicated Zebrafish. Toxins, 7(1), 97-109. https://doi.org/10.3390/toxins7010097