
Centipede Venom: Recent Discoveries and Current State of Knowledge



Abstract
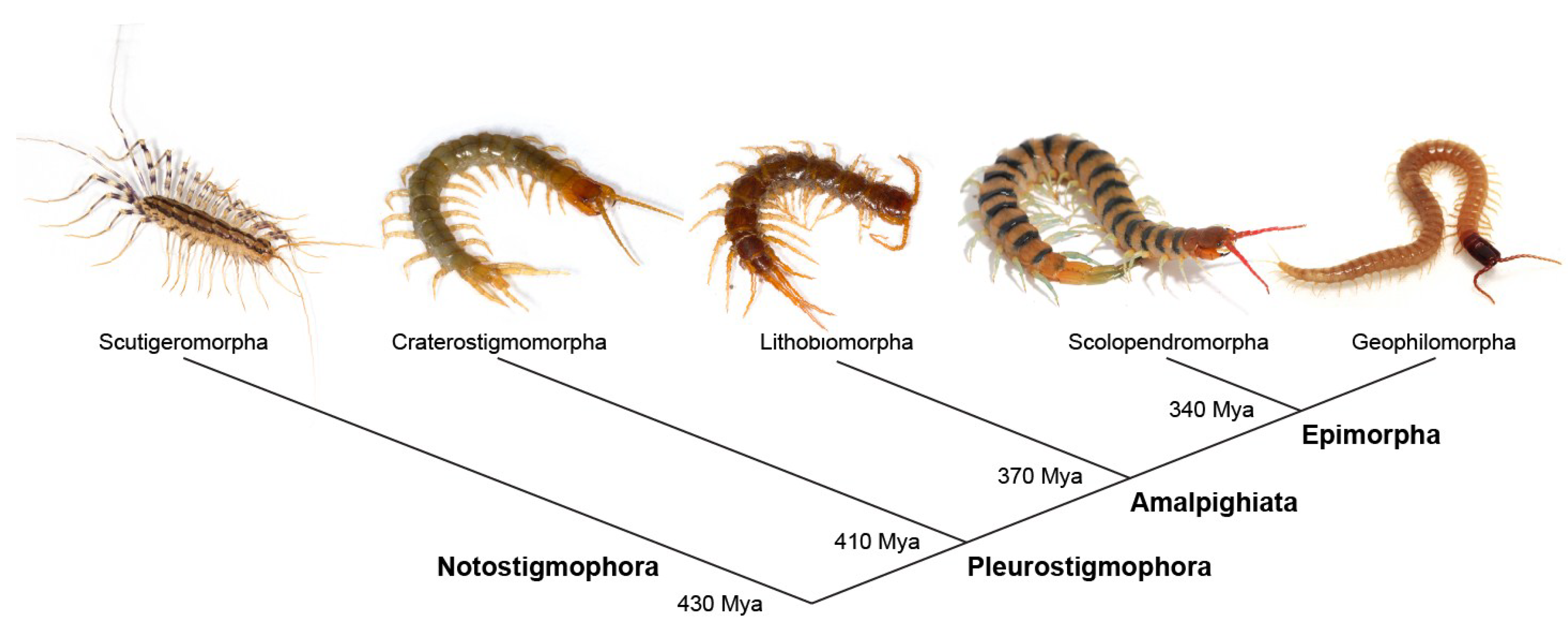
:1. Introduction

2. Venom Apparatus
3. Molecular and Pharmacological Diversity
3.1. Molecular and Pharmacological Diversity—Enzymes
3.1.1. Metalloproteases

{kind=link}
{kind=link}
| Family name | Type | Function | Earliest known recruitment | |
|---|---|---|---|---|
| Enzymes | ||||
| Protease M12A | Zinc metalloendopeptidase | Unknown, potential spreading factor | Basal | |
| Protease S1 | Serine protease | Potentially involved in activation of toxins | Basal | |
| Protease S8 | Serine protease | Potentially involved in activation of toxins | Scolopendridae | |
| γ-GT | γ-Glutamyltransferase | Platelet aggregating activity, hemolytic to mouse and rabbit hemocytes | Basal | |
| Chitinase | Glycoside hydrolase family 18 | Unknown | Scolopendridae | |
| Lysozyme C | Glycoside hydrolase family 22 | Potential antimicrobial component | Scolopendridae | |
| Hyaluronidase | Glycoside hydrolase family 56 | Degrades glycosaminoglycans, potentially facilitating the spread of venom components | Scolopendridae | |
| GDH | Glucose dehydrogenase | Unknown | Basal | |
| Carboxylesterase | Type B carboxylesterase | Unknown | Basal | |
| CentiPAD | Peptidylarginine deiminase | Venom activity unknown; catalyses deamination of the guanidine group of arginine residues, potentially involved in post-translational modification of toxins | Thereuopoda longicornis | |
| ScolPLA2 | Phospholipase type A2 | Venom activity unknown; venom PLA2 can be myotoxic, inflammatory, and neurotoxic | Scolopendridae | |
| Non-enzymatic proteins | ||||
| β-PFTx | β-Pore-forming toxin | Potentially cytotoxic via formation of polymeric pore structures in cell membranes | Basal | |
| CentiCAP1 | CAP protein | Unknown | Basal | |
| CentiCAP2 | CAP protein | CaV channel antagonist (KC144967); Trypsin inhibitor (KC144061) | Scolopendridae | |
| CentiCAP3 | CAP protein | Unknown | Scolopendra morsitans | |
| LDLA protein | LDLA-repeat domain containing protein | Unknown | Basal | |
| Cystatin | Cystatin | Potential protease inhibitor | Ethmostigmus rubripes | |
| Transferrin | Transferrin | Potential antimicrobial component | Basal | |
| DUF3472 | Protein containing a domain of unknown function type 3472 | Unknown | Scolopendridae | |
| DUF1397 | Protein containing a domain of unknown function type 1397 | Unknown | Thereuopoda longicornis | |
| Completely uncharacterized proteins | ||||
| Family 1 | Unknown | Unknown | Scolopendridae | |
| Family 2 | Unknown | Unknown | Scolopendra morsitans | |
| Family 3 | Unknown | Unknown | Scolopendrinae | |
| Family 4 | Unknown | Unknown | Thereuopoda longicornis | |
| Family 5 | Similar to hypothetical protein from Drosophila mojavensis (XP_002005038.1, BLAST E-value 4.42E-4) | Unknown | Scolopendridae | |
| Family 6 | Unknown | Unknown | Scolopendridae | |
| Family 7 | Similar to hypothetical protein from Chthionobacter flavus (EDY20616.1, BLAST E-value 6.13E-7) | Unknown | Scolopendra morsitans | |
| Family 8 | Unknown | Unknown | Thereuopoda longicornis | |
| Family 9 | Unknown | Unknown | Scolopendra morsitans | |
| Family 10 | Unknown | Unknown | Scolopendra morsitans | |
| Family 11 | Unknown | Unknown | Scolopendra spp. | |
| Peptides | ||||
| SCUTX 1 | 2 cysteines | C–C | Unknown (e.g., GASR01000100) | Thereuopoda longicornis |
| SCUTX 2 | 8 cysteines, includes SLPTX family 27 | C–C–C–CC–CC | Unknown (e.g., GASR01000101; JZ722897–9) | Basal |
| SCUTX 3 | Proline-rich linear peptides | Unknown (e.g., GASR01000107) | Thereuopoda longicornis | |
| SLPTX 1 | 6 cysteines and a type 2 chitin-binding domain | C–C–C–C–C–C | Unknown (e.g., GASI01000092) | Basal |
| SLPTX 2 | Defensin-like with 6 cysteines | C–C–C–C–CxC | Unknown (e.g., GASI01000163) | Ethmostigmus rubripes |
| SLPTX 3 | Helical peptides with 6 cysteines | C–C–C–CC–C | Unknown; KV antagonist (JN646114); NaV channel antagonist (UniProt: PODL36) | Scolopendra spp. |
| SLPTX 4 | 4 cysteines; transcripts may encode additional linear peptides upstream of cysteine-rich peptide | C–C–C–C | Unknown; KV channel antagonist (KC144226); putative synergistic mode of action for peptides encoded by multidomain transcripts (e.g., U-SLPTX4-Er1.1 and U-SLPTX4-Er1.2 from KF130724). | Scolopendridae |
| SLPTX 5 | 5–11 cysteines | C–C–C–C–C–C–C–C–C–C–C | Unknown; CaV channel agonist (JN646117) | Scolopendrinae |
| SLPTX 6 | 4 cysteines | CxC–CxC | Unknown (e.g., GASH01000180) | Scolopendra morsitans |
| SLPTX 7 | Putative ICK fold with 6 cysteines | C–C–C–C–CC | KV channel antagonist (JN646115) | Scolopendra subspinipes |
| SLPTX 8 | Multiple linear peptides encoded by the same transcript, sometimes upstream of cysteine-rich peptides with 6 cysteines | C–C–C–CCC | Unknown (e.g., KF130762, JZ722863); putative synergistic mode of action (e.g., U-SLPTX8-Er5.1a and U-SLPTX8-Er5.2a from KF130754) | Scolopendridae |
| SLPTX 9 | 6–8 cysteines; transcripts may encode additional linear peptides downstream of cysteine-rich peptide | C–CxC–C–C–C | Unknown; putative synergistic mode of action for peptides encoded by multidomain transcripts (e.g., U-SLPTX9-Er4.1a and U-SLPTX9-Er4.2a from KF130739) | Scolopendridae |
| SLPTX 10 | 6 cysteines | C–C–C–CC–C | Unknown; KV channel antagonist (KC144849); CaV channel antagonist (KC144448) | Scolopendridae |
| SLPTX 11 | 4–18 cysteines | C–C–CxC–C–C–C–CxC–C–C–C–CxC–C–C (e.g., KC144104); C–CxC–C (e.g., JN646116) | Unknown; KV channel antagonists (e.g., JN646116, KC144104); Anticoagulant (KC144430) | Scolopendra spp. |
| SLPTX 12 | 7 cysteines | C–C–CxC–CxC–C | Unknown (e.g., GASI01000120) | Scolopendridae |
| SLPTX 13 | 8 cysteines | C–C–CC–C–C–CxC | Unknown; CaV channel antagonists (JN646118) | Scolopendridae |
| SLPTX 14 | 8 cysteines | C–C–C–CC–CxCxC | Unknown (e.g., GASI01000125) | Scolopendridae |
| SLPTX 15 | 4–6 cysteines | C–C–CxC | Unknown; KV channel antagonists (KC144556); NaV antagonists (KC144793); CaV channel antagonists (KC145039) | Scolopendridae |
| SLPTX 16 | Von Willebrand factor type C; peptides with 3–9 but predominantly 8 cysteines | C–C–C–C–C–CC–C; C–C–C–C–C–CCC–C (e.g., GASI01000127) | Unknown (e.g., GASI01000135) | Scolopendridae |
| SLPTX 17 | Predominantly 8 cysteines | C–C–C–CC–C–C–C | Unknown (e.g., GASI01000156) | Ethmostigmus rubripes |
| SLPTX 18 | Colipase-like peptides with 10 cysteines | C–C–CC–C–C–CxC–C–C | Putative colipase, same superfamily as AVIT-toxins which induce smooth muscle contraction and hyperalgesia (GASI01000011) | Ethmostigmus rubripes |
| SLPTX 19 | 12 cysteines | C–C–C–C–CC–C–C–C–C–CC | Putative carboxypeptidase inhibitor (e.g., GASH01000169) | Basal |
| SLPTX 20 | 6 cysteines | C–C–C–C–CC | Unknown (e.g., GASH01000170) | Scolopendrinae |
| SLPTX 21 | Linear diuretic hormone-like peptide | Unknown (e.g., GASH01000171) | Scolopendra morsitans | |
| SLPTX 22 | Linear hypertrehalosaemic hormone-like peptide | Unknown (e.g., GASI01000170) | Scolopendridae | |
| SLPTX 23 | Linear peptide | Unknown (e.g., GASH01000173) | Ethmostigmus rubripes | |
| SLPTX 24 | Linear peptide | Unknown (e.g., GASH01000177) | Ethmostigmus rubripes | |
| SLPTX 25 | Linear peptide | Unknown (e.g., GASH01000182) | Ethmostigmus rubripes | |
| SLPTX 26 | 7 cysteines | C–C–C–C–C–CC | Unknown (JZ722896) | Scolopendra subspinipes mutilans [32] |
| SLPTX 28 | 3 cysteines | C–CC | Unknown (JZ722900) | Scolopendra subspinipes mutilans [32] |

3.1.2. Serine Proteases
3.1.3. γ-Glutamyl Transpeptidase
3.1.4. Glycoside Hydrolases
3.1.5. Phospholipase A2
3.1.6. Other Enzymes
3.2. Molecular and Pharmacological Diversity—Non-Enzymatic Proteins
3.2.1. Centipede β-Pore-Forming Toxins
3.2.2. CAP Proteins
3.2.3. LDLA Domain-Containing Proteins
3.2.4. Other Non-Enzymatic Proteins
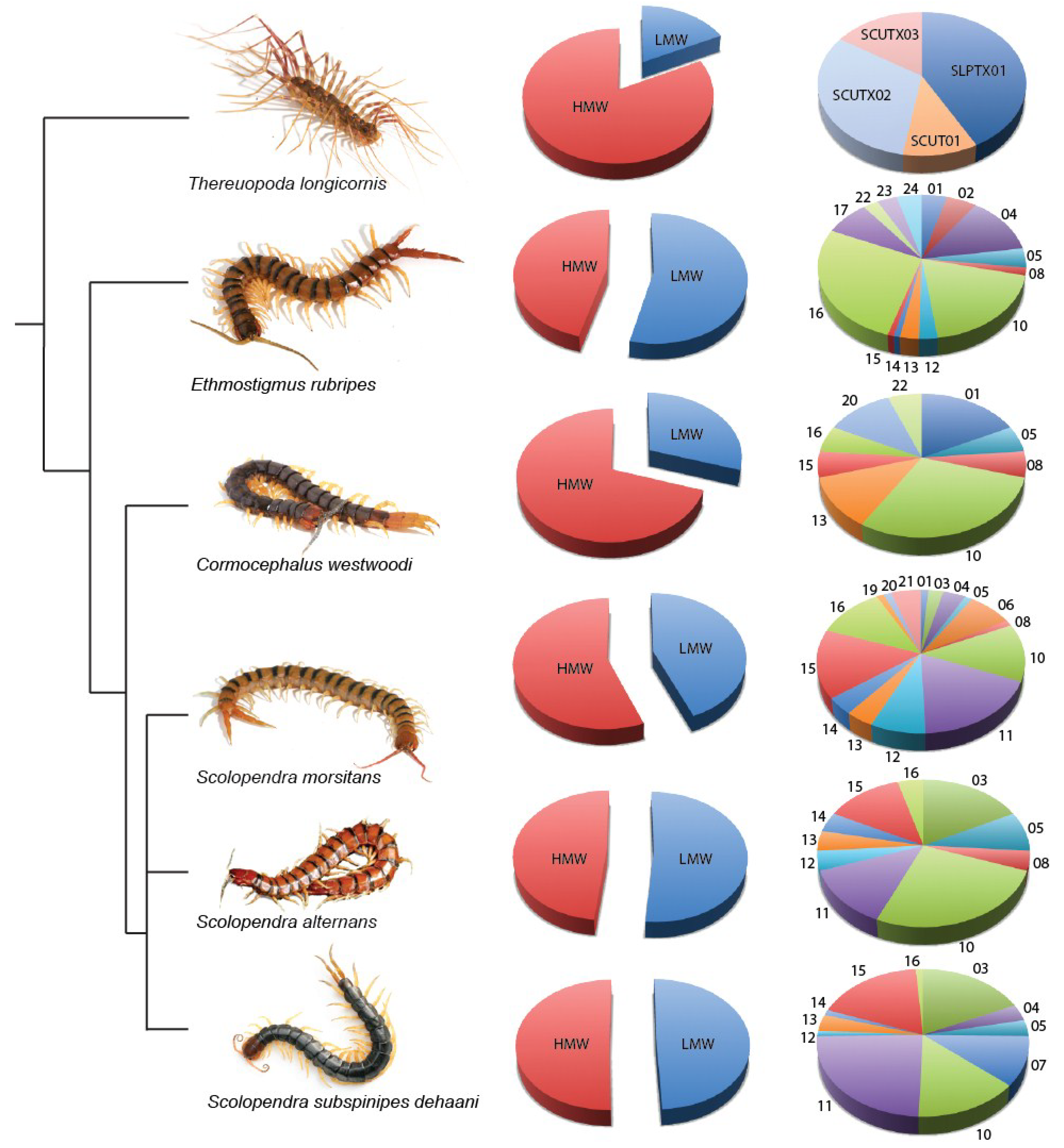
3.3. Molecular and Pharmacological Diversity—Peptides
3.3.1. Molecular Diversity of Centipede Venom Peptides
3.3.2. Pharmacological Diversity of Centipede Venom Peptides
4. Clinical Importance of Centipede Stings
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bonato, L.; Edgecombe, G.; Lewis, J.; Minelli, A.; Pereira, L.; Shelley, R.; Zapparoli, M. A common terminology for the external anatomy of centipedes (Chilopoda). ZooKeys 2010, 69, 17–51. [Google Scholar] [CrossRef] [PubMed]
- Undheim, E.A.B.; King, G.F. On the venom system of centipedes (Chilopoda), a neglected group of venomous animals. Toxicon 2011, 57, 512–524. [Google Scholar] [CrossRef] [PubMed]
- Fernández, R.; Laumer, C.E.; Vahtera, V.; Libro, S.; Kaluziak, S.; Sharma, P.P.; Pérez-Porro, A.R.; Edgecombe, G.D.; Giribet, G. Evaluating topological conflict in centipede phylogeny using transcriptomic data sets. Mol. Biol. Evol. 2014, 31, 1500–1513. [Google Scholar] [CrossRef] [PubMed]
- Shear, W.A.; Jeram, A.J.; Selden, P.A. Centipede legs (Arthropoda, Chilopoda, Scutigeromorpha) from the Silurian and Devonian of Britain and the Devonian of North America. Am. Mus. Novit. 1998, 3231, 1–16. [Google Scholar]
- Anderson, L.I.; Trewin, N.H. An Early Devonian arthropod fauna from the Windyfield cherts, Aberdeenshire, Scotland. Palaeontology 2003, 46, 467–509. [Google Scholar] [CrossRef]
- Dunlop, J.A. Geological history and phylogeny of Chelicerata. Arthropod Struct. Dev. 2010, 39, 124–142. [Google Scholar] [CrossRef] [PubMed]
- Selden, P.A.; Shear, W.A.; Sutton, M.D. Fossil evidence for the origin of spider spinnerets, and a proposed arachnid order. Proc. Natl. Acad. Sci. USA 2008, 105, 20781–20785. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.M.; Fox, G.A.; Nelsen, D.R.; Hayes, W.K. Variation in venom yield and protein concentration of the centipedes Scolopendra polymorpha and Scolopendra subspinipes. Toxicon 2014, 82, 30–51. [Google Scholar] [CrossRef] [PubMed]
- Von Reumont, B.M.; Campbell, L.I.; Jenner, R.A. Quo vadis venomics? A roadmap to neglected venomous invertebrates. Toxins (Basel) 2014, 6, 3488–3551. [Google Scholar] [CrossRef]
- Ménez, A.; Zimmerman, K.; Zimmerman, S.; Heatwole, H. Venom apparatus and toxicity of the centipede Ethmostigmus rubripes (Chilopoda, Scolopendridae). J. Morphol. 1990, 206, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Ernst, A.; Rosenberg, J. Structure and distribution of sensilla coeloconica on the maxillipedes of Chilopoda. Afr. Invertebr. 2003, 44, 155–168. [Google Scholar]
- Haug, J.T.; Haug, C.; Schweigert, G.; Sombke, A. The evolution of centipede venom claws—Open questions and possible answers. Arthropod Struct. Dev. 2014, 43, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Antoniazzi, M.M.; Pedroso, C.M.; Knysak, I.; Martins, R.; Guizze, S.P.G.; Jared, C.; Barbaro, K.C. Comparative morphological study of the venom glands of the centipede Cryptops iheringi, Otostigmus pradoi and Scolopendra viridicornis. Toxicon 2009, 53, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Duboscq, O. Recherches sur les chilopodes. Arch. Zool. Exp. Gén. 1898, 6, 481–650. [Google Scholar]
- Lewis, J.G.E. The scolopendromorph centipedes of Mauritius and Rodrigues and their adjacent islets (Chilopoda: Scolopendromorpha). J. Nat. Hist. 2002, 36, 79–106. [Google Scholar] [CrossRef]
- Edgecombe, G.D.; Koch, M. Phylogeny of scolopendromorph centipedes (Chilopoda): Morphological analysis featuring characters from the peristomatic area. Cladistics 2008, 24, 872–901. [Google Scholar] [CrossRef]
- Pereira, L.A.; Uliana, M.; Minelli, A. Geophilomorph centipedes (Chilopoda) from termite mounds in the northern Pantanal wetland of Mato Grosso, Brazil. Stud. Neotrop. Fauna Environ. 2007, 42, 33–48. [Google Scholar] [CrossRef]
- Dugon, M.M.; Hayden, L.; Black, A.; Arthur, W. Development of the venom ducts in the centipede Scolopendra: An example of recapitulation. Evol. Dev. 2012, 14, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.; Hilken, G. Fine structural organization of the poison gland of Lithobius forficatus (Chilopoda, Lithobiomorpha). Nor. J. Entomol. 2006, 53, 119–127. [Google Scholar]
- Dugon, M.M.; Arthur, W. Comparative studies on the structure and development of the venom-delivery system of centipedes, and a hypothesis on the origin of this evolutionary novelty. Evol. Dev. 2012, 14, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.H.; Abu-Sinna, G.; el-Shabaka, H.A.; abd el-Aal, A. Proteins, lipids, lipoproteins and some enzyme characterizations of the venom extract from the centipede Scolopendra morsitans. Toxicon 1983, 21, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.H.; Batty, C.S. 5-Hydroxytryptamine content of some arthropod venoms and venom-containing parts. Toxicon 1963, 1, 165–170. [Google Scholar] [CrossRef]
- Gomes, A.; Datta, A.; Sarangi, B.; Kar, P.K.; Lahiri, S.C. Isolation, purification and pharmacodynamics of a toxin from the venom of the centipede Scolopendra subspinipes dehaani Brandt. Indian J. Exp. Biol. 1983, 21, 203–207. [Google Scholar] [PubMed]
- Stankiewicz, M.; Hamon, A.; Benkhalifa, R.; Kadziela, W.; Hue, B.; Lucas, S.; Mebs, D.; Pelhate, M. Effects of a centipede venom fraction on insect nervous system, a native Xenopus oocyte receptor and on an expressed Drosophila muscarinic receptor. Toxicon 1999, 37, 1431–1445. [Google Scholar] [CrossRef] [PubMed]
- Rates, B.; Bemquerer, M.P.; Richardson, M.; Borges, M.H.; Morales, R.A.V.; de Lima, M.E.; Pimenta, A. Venomic analyses of Scolopendra viridicornis nigra and Scolopendra angulata (Centipede, Scolopendromorpha): Shedding light on venoms from a neglected group. Toxicon 2007, 49, 810–826. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-C.; Zhang, R.; Zhao, F.; Chen, Z.-M.; Liu, H.-W.; Wang, Y.-J.; Jiang, P.; Zhang, Y.; Wu, Y.; Ding, J.-P.; et al. Venomic and transcriptomic analysis of centipede Scolopendra subspinipes dehaani. J. Proteome Res. 2012, 11, 6197–6212. [Google Scholar] [PubMed]
- Malta, M.B.; Lira, M.S.; Soares, S.L.; Rocha, G.C.; Knysak, I.; Martins, R.; Guizze, S.P.G.; Santoro, M.L.; Barbaro, K.C. Toxic activities of Brazilian centipede venoms. Toxicon 2008, 52, 255–263. [Google Scholar] [CrossRef] [PubMed]
- González-Morales, L.; Diego-GarcÌa, E.; Segovia, L.; Gutiérrez, M.D.C.; Possani, L.D. Venom from the centipede Scolopendra viridis Say: Purification, gene cloning and phylogenetic analysis of a phospholipase A2. Toxicon 2009, 54, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Undheim, E.A.B.; Jones, A.; Clauser, K.R.; Holland, J.H.; Pineda, S.S.; King, G.F.; Fry, B.G. Clawing through evolution: Toxin diversification and convergence in the ancient lineage Chilopoda (Centipedes). Mol. Biol. Evol. 2014, 31, 2124–2148. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.C. Extra-oral digestion in predaceous terrestrial Arthropoda. Annu. Rev. Entomol. 1995, 40, 85–103. [Google Scholar] [CrossRef]
- González-Morales, L.; Pedraza-Escalona, M.; Diego-Garcia, E.; Restano-Cassulini, R.; Batista, C.V.F.; Gutiérrez, M.C.; Possani, L.D. Proteomic characterization of the venom and transcriptomic analysis of the venomous gland from the Mexican centipede Scolopendra viridis. J. Proteomics 2014, 111, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Rong, M.; Yang, S.; Wen, B.; Mo, G.; Kang, D.; Liu, J.; Lin, Z.; Jiang, W.; Li, B.; Du, C.; et al. Peptidomics combined with cDNA library unravel the diversity of centipede venom. J. Proteomics 2014, 114, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.J.; Rawlings, N.D.; Woessner, J.F. Handbook of Proteolytic Enzymes, 2nd ed.; Elsevier: London, UK, 2004. [Google Scholar]
- Brust, A.; Sunagar, K.; Undheim, E.A.; Vetter, I.; Yang, D.C.; Casewell, N.R.; Jackson, T.N.; Koludarov, I.; Alewood, P.F.; Hodgson, W.C.; et al. Differential evolution and neofunctionalization of snake venom metalloprotease domains. Mol. Cell. Proteomics 2013, 12, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, R.B.; Wille, A.C.; Chaim, O.M.; Appel, M.H.; Silva, D.T.; Franco, C.R.; Toma, L.; Mangili, O.C.; Gremski, W.; Dietrich, C.P.; et al. Identification, cloning, expression and functional characterization of an astacin-like metalloprotease toxin from Loxosceles intermedia (brown spider) venom. Biochem. J. 2007, 406, 355–363. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, D.C.; Aerts, M.; Brunain, M.; Desjardins, C.A.; Jacobs, F.J.; Werren, J.H.; Devreese, B. Insights into the venom composition of the ectoparasitoid wasp Nasonia vitripennis from bioinformatic and proteomic studies. Insect Mol. Biol. 2010, 19, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Francischetti, I.M.; Mather, T.N.; Ribeiro, J.M. Cloning of a salivary gland metalloprotease and characterization of gelatinase and fibrin(ogen)lytic activities in the saliva of the Lyme disease tick vector Ixodes scapularis. Biochem. Biophys. Res. Commun. 2003, 305, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Ruder, T.; Sunagar, K.; Undheim, E.A.; Ali, S.A.; Wai, T.-C.; Low, D.H.; Jackson, T.N.; King, G.F.; Antunes, A.; Fry, B.G. Molecular phylogeny and evolution of the proteins encoded by coleoid (Cuttlefish, Octopus, and Squid) posterior venom glands. J. Mol. Evol. 2013, 76, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Weston, A.J.; Chung, R.; Dunlap, W.C.; Morandini, A.C.; Marques, A.C.; Moura-da-Silva, A.M.; Ward, M.; Padilla, G.; da Silva, L.F.; Andreakis, N.; et al. Proteomic characterisation of toxins isolated from nematocysts of the South Atlantic jellyfish Olindias sambaquiensis. Toxicon 2013, 71, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Wong, E.S.; Papenfuss, A.T.; Whittington, C.M.; Warren, W.C.; Belov, K. A limited role for gene duplications in the evolution of platypus venom. Mol. Biol. Evol. 2012, 29, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Praher, D.; Schlesinger, A.; Ayalon, A.; Tal, Y.; Technau, U. Analysis of soluble protein contents from the nematocysts of a model sea anemone sheds light on venom evolution. Mar. Biotechnol. 2013, 15, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Bond, J.S.; Beynon, R.J. The astacin family of metalloendopeptidases. Protein Sci. 1995, 4, 1247–1261. [Google Scholar] [CrossRef] [PubMed]
- King, G.F.; Hardy, M.C. Spider-venom peptides: Structure, pharmacology, and potential for control of insect pests. Annu. Rev. Entomol. 2013, 58, 475–496. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Luo, F.; He, J.; Cao, Z.; Miao, L. Molecular cloning and characterization of a new cDNA sequence encoding a venom peptide from the centipede Scolopendra subspinipes mutilans. Mol. Biol. (Mosk.) 2012, 46, 570–575. [Google Scholar] [CrossRef]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.A.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Human Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Low, D.H.W.; Sunagar, K.; Undheim, E.A.B.; Ali, S.A.; Alagon, A.C.; Jones, A.; Jackson, T.N.W.; Gonzalez, S.P.; King, G.F.; Antunes, A.; et al. Dracula’s children: The proteomics, transcriptomics and molecular evolution of vampire bat venom proteins. J. Proteomics 2013, 89, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; He, Y.; Zhao, R.; Wu, Y.; Li, W.; Cao, Z. Extreme diversity of scorpion venom peptides and proteins revealed by transcriptomic analysis: Implication for proteome evolution of scorpion venom arsenal. J. Proteomics 2012, 75, 1563–1576. [Google Scholar] [CrossRef] [PubMed]
- Knapp, O.; Stiles, B.; Popoff, M.R. The aerolysin-like toxin family of cytolytic, pore-forming toxins. Open Toxinol. J. 2010, 3, 53–68. [Google Scholar] [CrossRef]
- Siezen, R.J.; Leunissen, J.A.M. Subtilases: The superfamily of subtilisin-like serine proteases. Protein Sci. 1997, 6, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Undheim, E.A.B.; Sunagar, K.; Hamilton, B.R.; Jones, A.; Venter, D.J.; Fry, B.G.; King, G.F. Multifunctional warheads: Diversification of the toxin arsenal of centipedes via novel multidomain transcripts. J. Proteomics 2014, 102, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Courtay, C.; Oster, T.; Michelet, F.; Visvikis, A.; Diederich, M.; Wellman, M.; Siest, G. y-glutamyltransferase: Nucleotide sequence of the human pancreatic cDNA. Evidence for a ubiquitous y-glutamyltransferase polypeptide in human tissues. Biochem. Pharmacol. 1992, 43, 2527–2533. [Google Scholar] [CrossRef] [PubMed]
- Falabella, P.; Riviello, L.; Caccialupi, P.; Rossodivita, T.; Teresa Valente, M.; Luisa de Stradis, M.; Tranfaglia, A.; Varricchio, P.; Gigliotti, S.; Graziani, F.; et al. A γ-glutamyl transpeptidase of Aphidius ervi venom induces apoptosis in the ovaries of host aphids. Insect Biochem. Mol. Biol. 2007, 37, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Joshi, J.; Karanth, K.P. Cretaceous–Tertiary diversification among select scolopendrid centipedes of South India. Mol. Phylogenet. Evol. 2011, 60, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, P.G.; Beckmann, A.; Warnken, U.; Schnolzer, M.; Schuler, A.; Bornberg-Bauer, E.; Holstein, T.W.; Ozbek, S. Proteome of Hydra nematocyst. J. Biol. Chem. 2012, 287, 9672–9681. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-H.; Jia, X.-T.; Zhao, L.; Li, C.-Z.; Zhang, S.; Chen, Y.-G.; Weng, S.-P.; He, J.-G. Identification and functional characterization of Dicer2 and five single VWC domain proteins of Litopenaeus vannamei. Dev. Comp. immunol. 2011, 35, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Fernandes-Pedrosa Mde, F.; Junqueira-de-Azevedo Ide, L.; Goncalves-de-Andrade, R.M.; Kobashi, L.S.; Almeida, D.D.; Ho, P.L.; Tambourgi, D.V. Transcriptome analysis of Loxosceles laeta (Araneae, Sicariidae) spider venomous gland using expressed sequence tags. BMC Genomics 2008, 9, 279. [Google Scholar] [CrossRef] [PubMed]
- Kramer, K.J.; Muthukrishnan, S. Insect chitinases: Molecular biology and potential use as biopesticides. Insect Biochem. Mol. Biol. 1998, 27, 887–900. [Google Scholar] [CrossRef]
- Chung, L.P.; Keshav, S.; Gordon, S. Cloning the human lysozyme cDNA: Inverted Alu repeat in the mRNA and in situ hybridization for macrophages and Paneth cells. Proc. Natl. Acad. Sci. USA 1988, 85, 6227–6231. [Google Scholar] [CrossRef] [PubMed]
- Girish, K.S.; Shashidharamurthy, R.; Nagaraju, S.; Gowda, T.V.; Kemparaju, K. Isolation and characterization of hyaluronidase a “spreading factor” from Indian cobra (Naja naja) venom. Biochimie 2004, 86, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Nentwig, L.; Schaller, J.; Nentwig, W. Biochemistry, toxicology and ecology of the venom of the spider Cupiennius salei (Ctenidae). Toxicon 2004, 43, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Long-Rowe, K.O.; Burnett, J.W. Characteristics of hyaluronidase and hemolytic activity in fishing tentacle nematocyst venom of Chrysaora quinquecirrha. Toxicon 1994, 32, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Wong, E.S.; Morgenstern, D.; Mofiz, E.; Gombert, S.; Morris, K.M.; Temple-Smith, P.; Renfree, M.B.; Whittington, C.M.; King, G.F.; Warren, W.C.; et al. Proteomics and deep sequencing comparison of seasonally active venom glands in the platypus reveals novel venom peptides and distinct expression profiles. Mol. Cell. Proteomics 2012, 11, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Violette, A.; Leonardi, A.; Piquemal, D.; Terrat, Y.; Biass, D.; Dutertre, S.; Noguier, F.; Ducancel, F.; Stocklin, R.; Krizaj, I.; Favreau, P. Recruitment of glycosyl hydrolase proteins in a cone snail venomous arsenal: Further insights into biomolecular features of Conus venoms. Mar. Drugs 2012, 10, 258–280. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.M.; Ghomashchi, F.; Gelb, M.H.; Dooley, D.J.; Stoehr, S.J.; Giordani, A.B.; Naisbitt, S.R.; Olivera, B.M. Conodipine-M, a novel phospholipase A2 isolated from the venom of the marine snail Conus magus. J. Biol. Chem. 1995, 270, 3518–3526. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G. From genome to “venome”: Molecular origin and evolution of the snake venom proteome inferred from phylogenetic analysis of toxin sequences and related body proteins. Genome Res. 2005, 15, 403–420. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, M.D.C.; Abarca, C.; Possani, L.D. A toxic fraction from scolopendra venom increases the basal release of neurotransmitters in the ventral ganglia of crustaceans. Comp. Biochem. Physiol. C 2003, 135, 205–214. [Google Scholar]
- Bak, T.G. Studies on glucose dehydrogenase of Aspergillus oryzae. II. Purification and physical and chemical properties. Biochim. Biophysi. Acta 1967, 139, 277–293. [Google Scholar] [CrossRef]
- Shirai, H.; Blundell, T.L.; Mizuguchi, K. A novel superfamily of enzymes that catalyze the modification of guanidino groups. Trends Biochem. Sci. 2001, 26, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Heitz, J.R.; Norment, B.R. Characteristics of an alkaline phosphatase activity in brown recluse venom. Toxicon 1974, 12, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.C.A.; Guimarães, L.H.S.; Liberato, J.L.; Polizeli, M.D.L.T.D.M.; dos Santos, W.F. Acid and alkaline phosphatase activities of a fraction isolated from Parawixia bistriata spider venom. Toxicon 2006, 47, 854–858. [Google Scholar] [CrossRef] [PubMed]
- Sulkowski, E.; Björk, W.; Laskowski, M. A specific and nonspecific alkaline monophosphatase in the venom of Bothrops atrox and their occurrence in the purified venom phosphodiesterase. J. Biol. Chem. 1963, 238, 2477–2486. [Google Scholar] [PubMed]
- Tu, A.T.; Chua, A. Acid and alkaline phosphomonoesterase activities in snake venoms. Comp. Biochem. Physiol. C: Toxicol. Pharmacol. 1966, 17, 297–307. [Google Scholar] [CrossRef]
- Undheim, E.A.B.; Georgieva, D.N.; Thoen, H.H.; Norman, J.A.; Mork, J.; Betzel, C.; Fry, B.G. Venom on ice: First insights into Antarctic octopus venoms. Toxicon 2010, 56, 897–913. [Google Scholar] [CrossRef] [PubMed]
- Aird, S.D. Ophidian envenomation strategies and the role of purines. Toxicon 2002, 40, 335–393. [Google Scholar] [CrossRef] [PubMed]
- Dhananjaya, B.L.; D’Souza, C.J.M. The pharmacological role of nucleotidases in snake venoms. Cell Biochem. Funct. 2010, 28, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Quistad, G.B.; Dennis, P.A.; Skinner, W.S. Insecticidal activity of spider (Araneae), centipede (Chilopoda), scorpion (Scorpionidae), and snake (Serpentes) venoms. J. Econ. Entom. 1992, 85, 33–39. [Google Scholar] [CrossRef]
- Peichoto, M.E.; Mackessy, S.P.; Teibler, P.; Tavares, F.L.; Burckhardt, P.L.; Breno, M.C.; Acosta, O.; Santoro, M.L. Purification and characterization of a cysteine-rich secretory protein from Philodryas patagoniensis snake venom. Comp. Biochem. Physiol. C 2009, 150, 79–84. [Google Scholar]
- Hoffman, D.R. Hymenoptera venom allergens. Clin. Rev. Allergy Immunol. 2006, 30, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Daly, N.L.; Scanlon, M.J.; Djordjevic, J.T.; Kroon, P.A.; Smith, R. Three-dimensional structure of a cysteine-rich repeat from the low-density lipoprotein receptor. Proc. Natl. Acad. Sci. USA 1995, 92, 6334–6338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Gomez, F.; Ortiz-Pineda, P.A.; Rojas-Cartagena, C.; Suarez-Castillo, E.C.; Garcia-Arraras, J.E. Immune-related genes associated with intestinal tissue in the sea cucumber Holothuria glaberrima. Immunogenetics 2008, 60, 57–71. [Google Scholar] [CrossRef]
- Fry, B.G.; Undheim, E.A.B.; Ali, S.A.; Debono, J.; Scheib, H.; Ruder, T.; Jackson, T.N.; Morgenstern, D.; Cadwallader, L.; Whitehead, D.; et al. Squeezers and leaf-cutters: Differential diversification and degeneration of the venom system in toxicoferan reptiles. Mol. Cell. Proteomics 2013, 12, 1881–1899. [Google Scholar] [CrossRef] [PubMed]
- Abrahamson, M.; Alvarez-Fernandez, M.; Nathanson, C.-M. Cystatins. Biochem. Soc. Symposia 2003, 70, 179–199. [Google Scholar]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Buczek, O.; Bulaj, G.; Olivera, B.M. Conotoxins and the posttranslational modification of secreted gene products. Cell. Mol. Life Sci. 2005, 62, 3067–3079. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Hernández, V.; Ortiz, E.; Rendón-Anaya, M.; Schwartz, E.F.; Becerril, B.; Corzo, G.; Possani, L.D. Scorpion and spider venom peptides: Gene cloning and peptide expression. Toxicon 2011, 58, 644–663. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Undheim, E.A.; Chan, A.H.; Koludarov, I.; Muñoz-Gómez, S.A.; Antunes, A.; Fry, B.G. Evolution stings: The origin and diversification of scorpion toxin peptide scaffolds. Toxins 2013, 5, 2456–2487. [Google Scholar] [CrossRef] [PubMed]
- Palagi, A.; Koh, J.M.; Leblanc, M.; Wilson, D.; Dutertre, S.; King, G.F.; Nicholson, G.M.; Escoubas, P. Unravelling the complex venom landscapes of lethal Australian funnel-web spiders (Hexathelidae: Atracinae) using LC-MALDI-TOF mass spectrometry. J. Proteomics 2013, 80, 292–310. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Kong, Y.; Zhai, L.; Wu, X.; Jia, P.; Liu, J.; Yu, H. Two novel antimicrobial peptides from centipede venoms. Toxicon 2009, 55, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liu, Z.; Xiao, Y.; Li, Y.; Rong, M.; Liang, S.; Zhang, Z.; Yu, H.; King, G.F.; Lai, R. Chemical punch packed in venoms makes centipedes excellent predators. Mol. Cell. Proteomics 2012, 11, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xiao, Y.; Kang, D.; Liu, J.; Li, Y.; Undheim, E.A.B.; Klint, J.K.; Rong, M.; Lai, R.; King, G.F. Discovery of a selective NaV1.7 inhibitor from centipede venom with analgesic efficacy exceeding morphine in rodent pain models. Proc. Natl. Acad. Sci. USA 2013, 110, 17534–17539. [Google Scholar] [CrossRef] [PubMed]
- Wenhua, R.; Shuangquan, Z.; Daxiang, S.; Kaiya, Z.; Guang, Y. Induction, purification and characterization of an antibacterial peptide scolopendrin I from the venom of centipede Scolopendra subspinipes mutilans. Indian J. Biochem. Biophys. 2006, 43, 88–93. [Google Scholar] [PubMed]
- Kong, Y.; Hui, J.; Shao, Y.; Huang, S.; Chen, H.; Wei, J.-F. Cytotoxic and anticoagulant peptide from Scolopendra subspinipes mutilans venom. Afr. J. Pharm. Pharmacol. 2013, 7, 2238–2245. [Google Scholar] [CrossRef]
- Kuhn-Nentwig, L. Antimicrobial and cytolytic peptides of venomous arthropods. Cell. Mol. Life Sci. 2003, 60, 2651–2668. [Google Scholar] [CrossRef] [PubMed]
- Knysak, I.; Martins, R.; Bertim, C.R. Epidemiological aspects of centipede (Scolopendromorphae: Chilopoda) bites registered in Greater S. Paulo, SP, Brazil. Rev. Saúde Pública 1998, 32, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Balit, C.R.; Harvey, M.S.; Waldock, J.M.; Isbister, G.K. Prospective study of centipede bites in Australia. Clin. Toxicol. 2004, 42, 41–48. [Google Scholar] [CrossRef]
- Lin, T.J.; Yang, C.C.; Yang, G.Y.; Ger, J.; Tsai, W.J.; Deng, J.F. Features of centipede bites in Taiwan. Trop. Geogr. Med. 1995, 47, 300–302. [Google Scholar] [PubMed]
- Forrester, J.A.; Holstege, C.P.; Forrester, J.D. Fatalities from venomous and nonvenomous animals in the United States (1999–2007). Wilderness Environ. Med. 2012, 23, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Langley, R.L. Animal-related fatalities in the United States—An update. Wilderness Environ. Med. 2005, 16, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Asa, K.; Imachi, T.; Yamaguchi, Y.; Yoshida, K. Centipede inflicted postmortem injury. J. Forensic Sci. 1999, 44, 849–850. [Google Scholar] [PubMed]
- Veraldi, S.; Çuka, E.; Gaiani, F. Scolopendra bites: A report of two cases and review of the literature. Int. J. Dermatol. 2014, 53, 869–872. [Google Scholar] [CrossRef] [PubMed]
- Blay, E.R. Treatment of centipede bites. Br. Med. J. 1955, 2, 1619. [Google Scholar] [CrossRef]
- Marsh, F. Centipede Bites. Br. Med. J. 1957, 2, 825. [Google Scholar] [CrossRef]
- Uzel, A.P.; Steinmann, G.; Bertino, R.; Korsaga, A. Dermohypodermite bactérienne et phlegmon du membre supérieur par morsure de scolopendre: À propos de deux cas. Chir. Main 2009, 28, 322–325. [Google Scholar] [CrossRef] [PubMed]
- Veraldi, S.; Chiaratti, A.; Sica, L. Centipede bite: A case report. Arch. Dermatol. 2010, 146, 807–808. [Google Scholar] [CrossRef] [PubMed]
- Serinken, M.; Erdur, B.; Sener, S.; Kabay, B.; Cevik, A. A case of mortal necrotizing fasciitis of the trunk resulting from a centipede (Scolopendra moritans) bitesite. Int. J. Emerg. Med. 2005, 2, 1–8. [Google Scholar]
- Pineda, E.V. A fatal case of centipede bite. J. Philippine Is. Med. Assoc. 1923, 3, 59–63. [Google Scholar]
- Remington, C.L. The bite and habits of a giant centipede (Scolopendra subspinipes) in the Philippine Islands. Am. J. Trop. Med. Hyg. 1950, 1, 453. [Google Scholar]
- Lewis, J.G.E.; Daszak, P.; Jones, C.G.; Cottingham, J.D.; Wenman, E.; Maljkovic, A. Field observations on three scolopendrid centipedes from Mauritius and Rodrigues (Indian Ocean) (Chilopoda: Scolopendromorpha). Int. J. Myriapodol. 2010, 3, 123–137. [Google Scholar] [CrossRef]
- Spillner, E.; Blank, S.; Jakob, T. Hymenoptera allergens: From venom to “venome”. Front. Immunol. 2014, 5, 77. [Google Scholar] [CrossRef] [PubMed]
- Harada, S.; Yoshizaki, Y.; Natsuaki, M.; Shimizu, H.; Fukuda, H.; Nagai, H.; Ikeda, T. Three cases of centipede allergy: Analysis of cross reactivity with bee allergy. Arerugi 2005, 54, 1279–1284. [Google Scholar] [PubMed]
- Bush, S.P.; King, B.O.; Noris, R.L.; Stockwell, S.A. Centipede envenomation. Wilderness Environ. Med. 2001, 12, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Chaou, C.H.; Chen, C.K.; Chen, J.C.; Chiu, T.F.; Lin, C.C. Comparisons of ice packs, hot water immersion, and analgesia injection for the treatment of centipede envenomations in Taiwan. Clin. Toxicol. 2009, 47, 659–662. [Google Scholar] [CrossRef]
- Fung, H.T. Centipede bite victims: A review of patients presenting to two emergency departments in Hong Kong. Hong Kong Med. J. 2011, 17, 381–385. [Google Scholar] [PubMed]
- Chipman, A.D.; Ferrier, D.E.K.; Brena, C.; Qu, J.; Hughes, D.S.T.; Schröder, R.; Torres-Oliva, M.; Znassi, N.; Jiang, H.; Almeida, F.C.; et al. The first myriapod genome sequence reveals conservative arthropod gene content and genome organisation in the centipede Strigamia maritima. PLoS Biol. 2014, 12, e1002005. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Undheim, E.A.B.; Fry, B.G.; King, G.F. Centipede Venom: Recent Discoveries and Current State of Knowledge. Toxins 2015, 7, 679-704. https://doi.org/10.3390/toxins7030679
Undheim EAB, Fry BG, King GF. Centipede Venom: Recent Discoveries and Current State of Knowledge. Toxins. 2015; 7(3):679-704. https://doi.org/10.3390/toxins7030679
Chicago/Turabian StyleUndheim, Eivind A. B., Bryan G. Fry, and Glenn F. King. 2015. "Centipede Venom: Recent Discoveries and Current State of Knowledge" Toxins 7, no. 3: 679-704. https://doi.org/10.3390/toxins7030679
APA StyleUndheim, E. A. B., Fry, B. G., & King, G. F. (2015). Centipede Venom: Recent Discoveries and Current State of Knowledge. Toxins, 7(3), 679-704. https://doi.org/10.3390/toxins7030679