
Honeybee (Apis mellifera) Venom Reinforces Viral Clearance during the Early Stage of Infection with Porcine Reproductive and Respiratory Syndrome Virus through the Up-Regulation of Th1-Specific Immune Responses

Abstract
:1. Introduction
2. Results
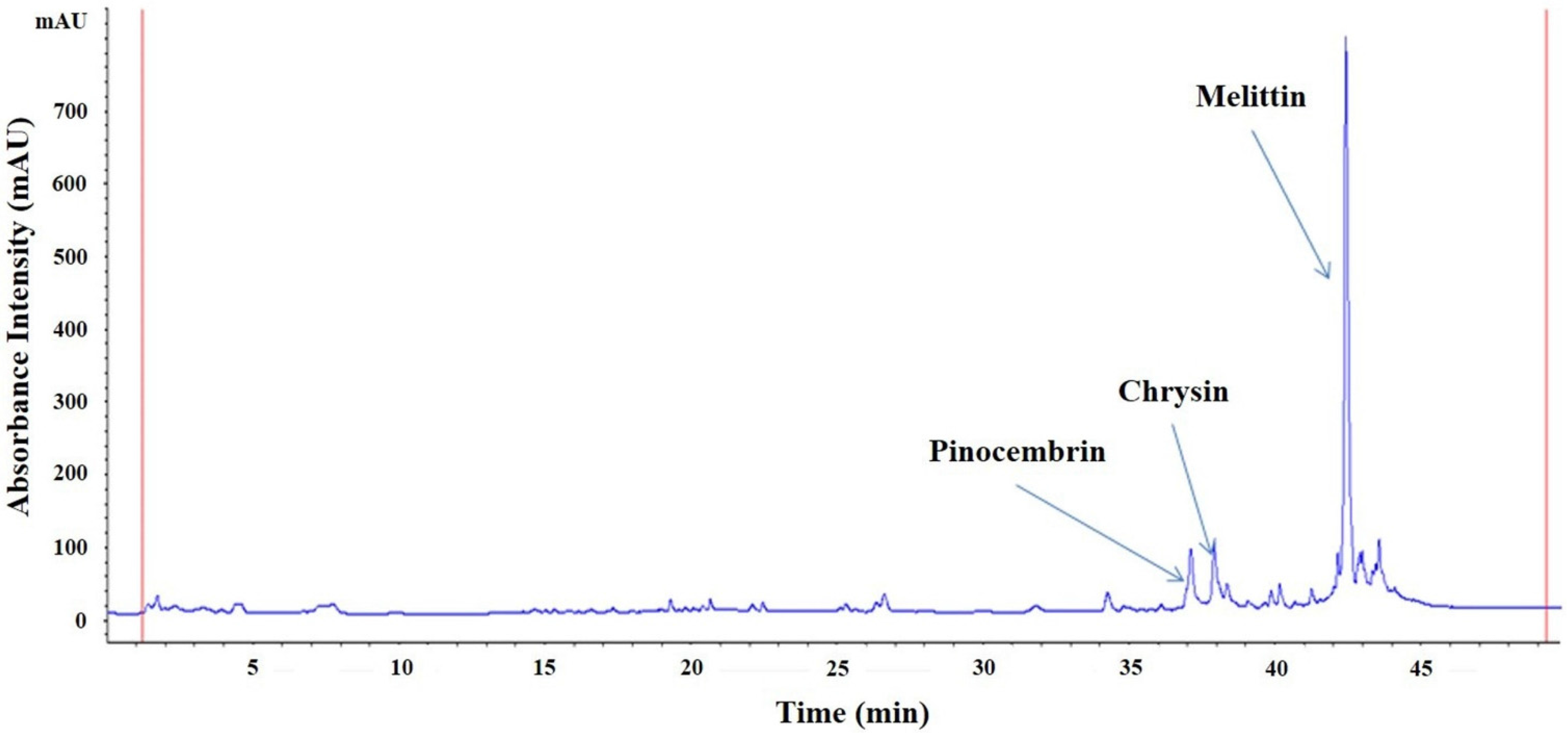
2.1. Characteristics of HBV

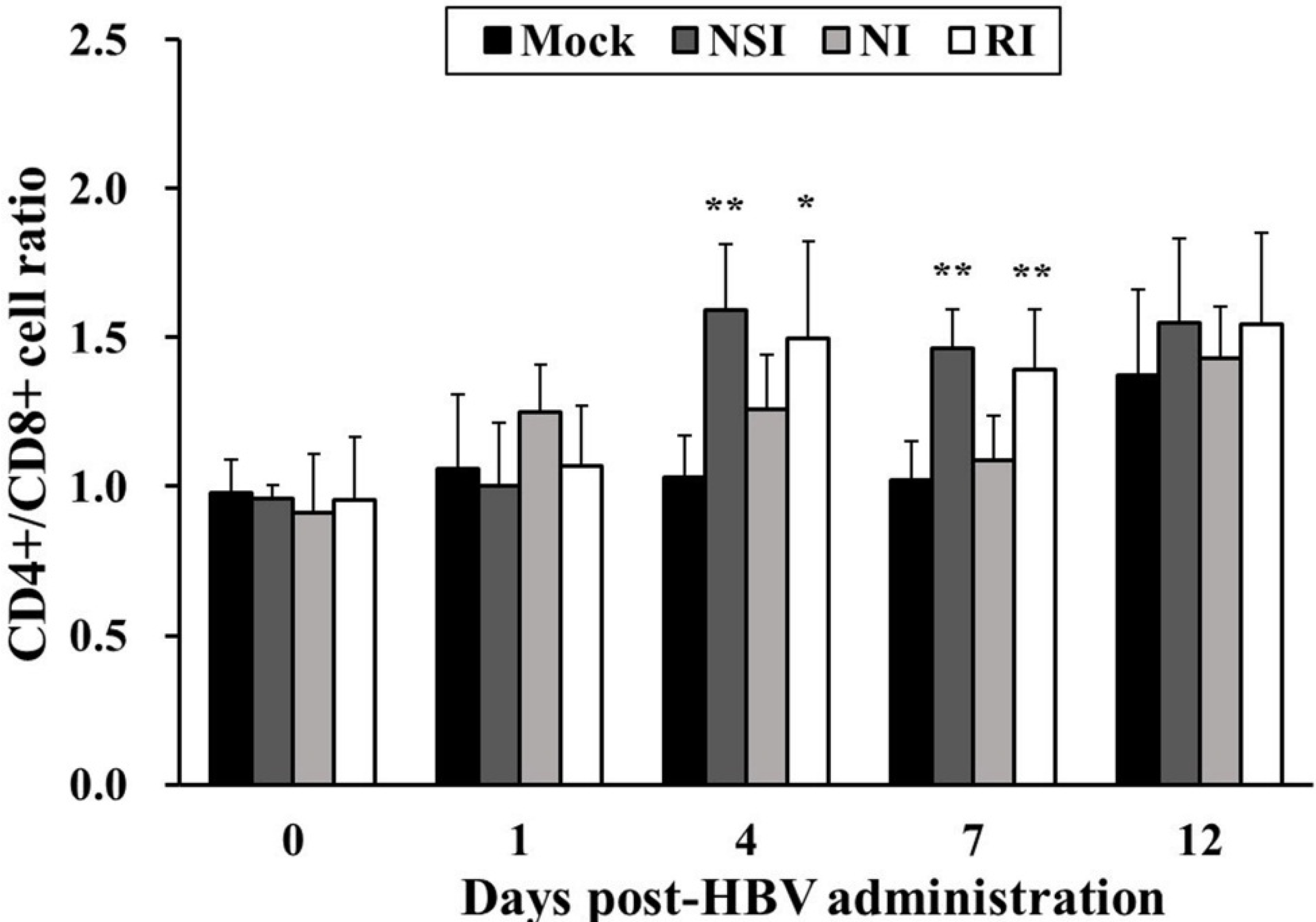
2.2. Effects of HBV on T Lymphocyte CD4+/CD8+ Subsets in Healthy Pigs

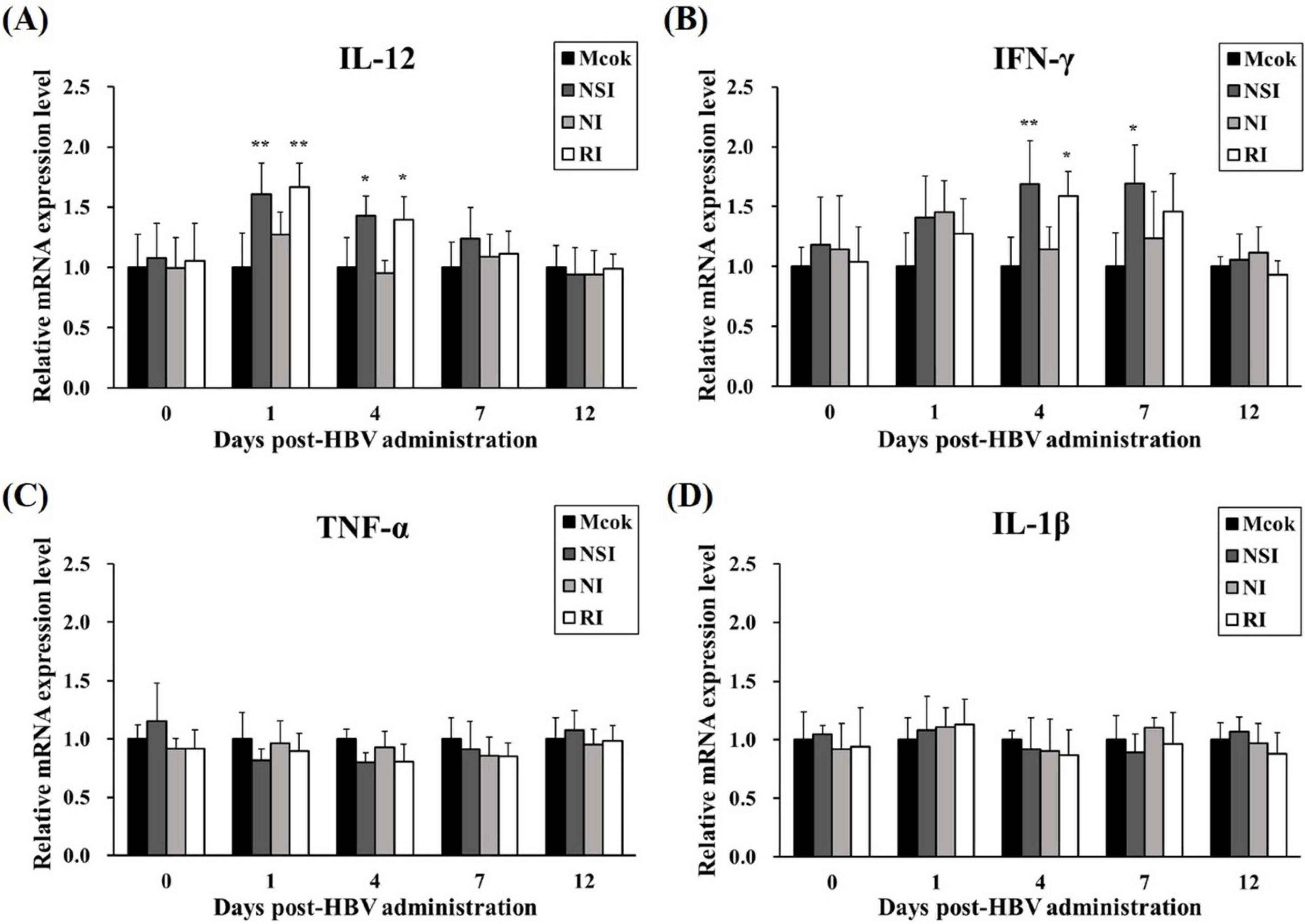
2.3. Effects of HBV on Cytokine Expression Levels in Healthy Pigs

2.4. Effects of HBV on Viral Clearance in the PRRSV Infected Pigs

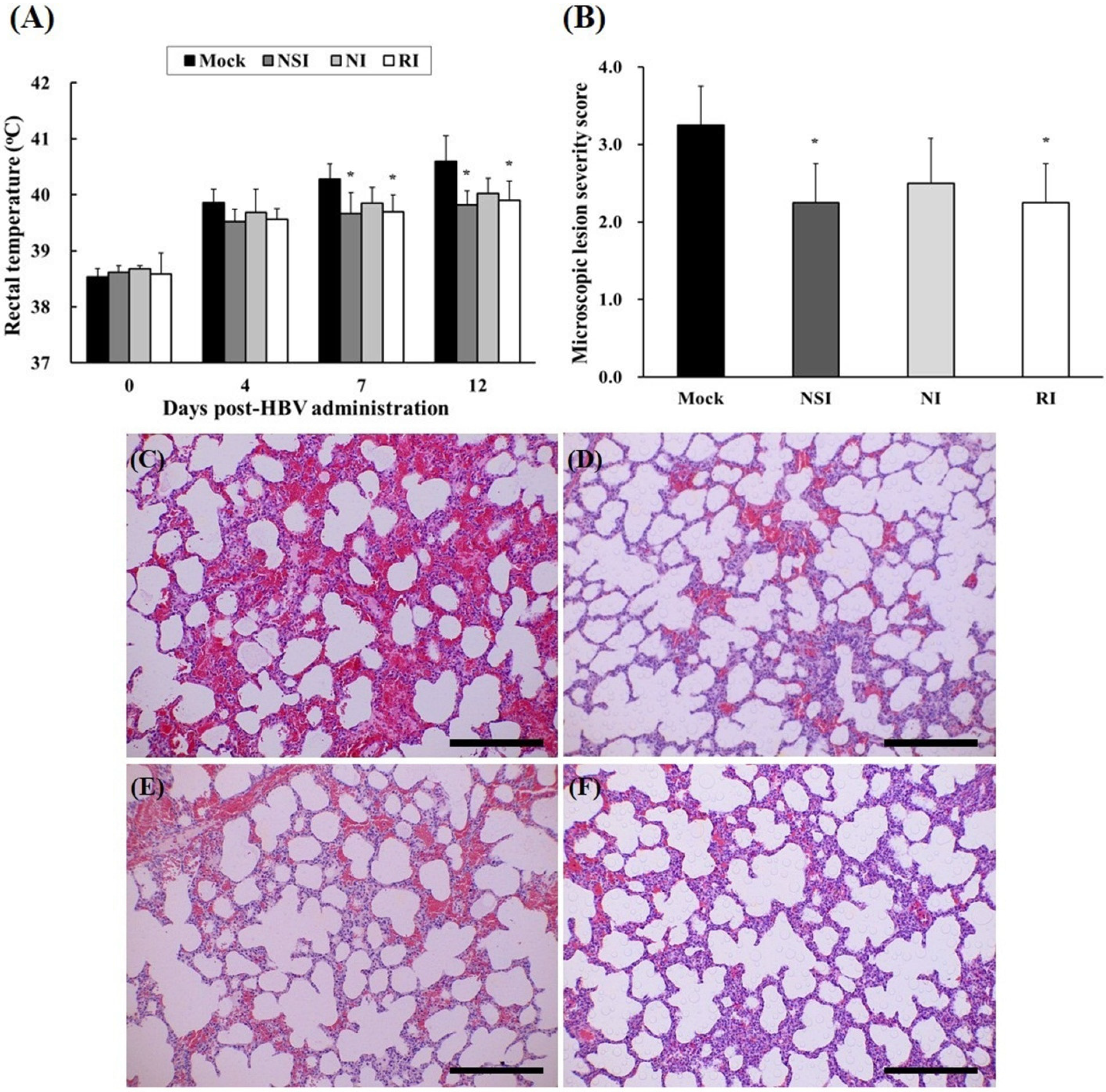
2.5. Evaluation of Body Temperature and Histopathological Examination of the PRRSV Infected Pigs
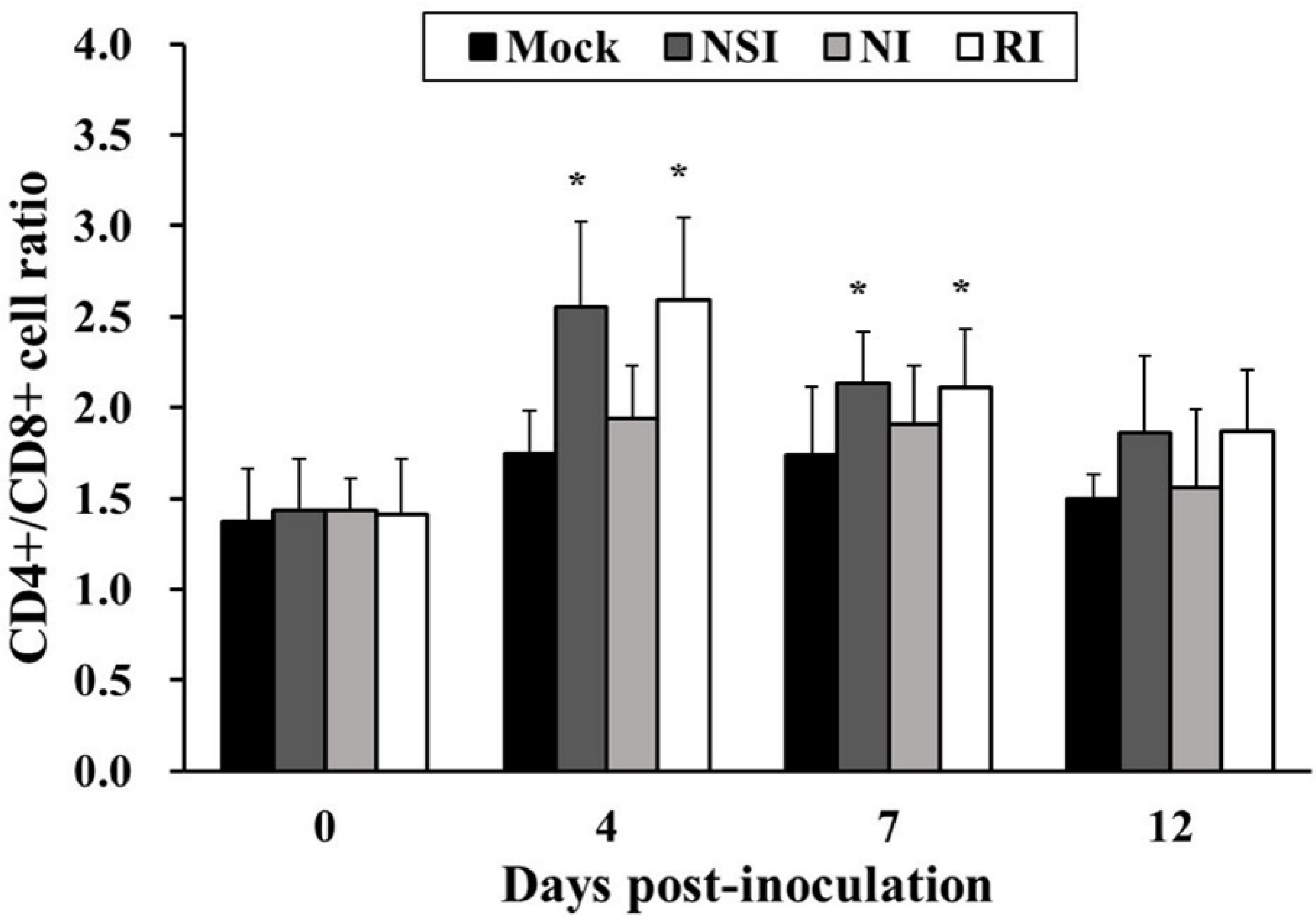
2.6. Effects of HBV on T Lymphocyte CD4+/CD8+ Subsets in the PRRSV Infected Pigs


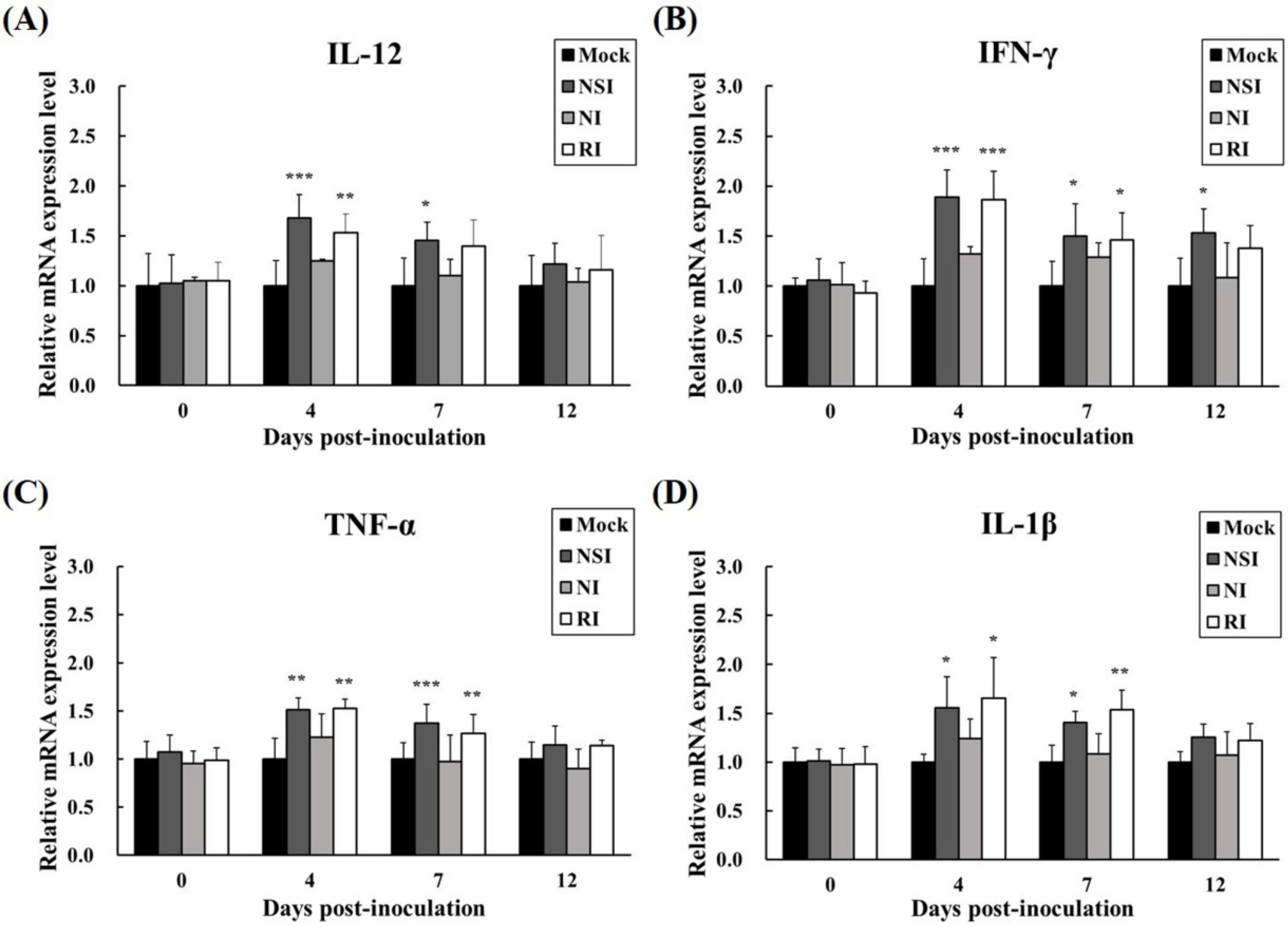
2.7. Effects of HBV on Cytokine Expression Profiles in the PRRSV Infected Pigs

3. Discussion
4. Materials and Methods
4.1. Preparation and Characterization of HBV
4.2. PRRS Virus
4.3. Animals
4.4. Experimental Protocol
4.4.1. Experiment 1: Immunostimulatory Effects of HBV on Healthy Pigs
4.4.2. Experiment 2: The Viral Clearance Effect of HBV in the Pigs Experimentally Infected with PRRSV
4.5. Isolation of PBMCs from Peripheral Blood
4.6. Analysis of T Lymphocyte CD4+/CD8+ Subsets by using Flow Cytometry
4.7. Analysis of Cytokine Expression Levels in PBMCs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5'–3') | Accession number | |
|---|---|---|---|
| TNF-α | Forward | CCCCCAGAAGGAAGAGTTTC | JF831365 |
| Reverse | CGGGCTTATCTGAGGTTTGA | ||
| IL-1β | Forward | GGCCGCCAAGATATAACTGA | NM_214055 |
| Reverse | GGACCTCTGGGTATGGCTTTC | ||
| IFN-γ | Forward | CAAAGCCATCAGTGAACTCATCA | X53085 |
| Reverse | TCTCTGGCCTTGGAACATAGTCT | ||
| IL-12 | Forward | GGAGTATAAGAAGTACAGAGTGG | U08317 |
| Reverse | GATGTCCCTGATGAAGAAGC | ||
| β-actin | Forward | CAGGTCATCACCATCGGCAACG | U07786 |
| Reverse | GACAGCACCGTGTTGGCGTAGAGGT | ||
4.8. Quantification of PRRSV Genome in Serum and Tissues using Quantitative Real-Time PCR
4.9. Histopathological Analysis in Lung Tissue of the Pigs Experimentally Infected with PRRSV
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oršolić, N. Bee venom in cancer therapy. Cancer Metastasis Rev. 2012, 31, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.; Ko, E.; Park, S.K.; Ko, S.; Jun, C.Y.; Shin, M.K.; Hong, M.C.; Bae, H. Bee venom modulates murine Th1/Th2 lineage development. Int. Immunopharmacol. 2005, 5, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Perrin-Cocon, L.; Agaugué, S.; Coutant, F.; Masurel, A.; Bezzine, S.; Lambeau, G.; André, P.; Lotteau, V. Secretory phospholipase A2 induces dendritic cell maturation. Eur. J. Immunol. 2004, 34, 2293–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramoner, R.; Putz, T.; Gander, H.; Rahm, A.; Bartsch, G.; Schaber, C.; Thurnher, M. Dendritic-cell activation by secretory phospholipase A2. Blood 2005, 105, 3583–3587. [Google Scholar] [CrossRef]
- Mataraci, E.; Dosler, S. In vitro activities of antibiotics and antimicrobial cationic peptides alone and in combination against methicillin-resistant Staphylococcus aureus biofilms. Antimicrob. Agents Chemother. 2012, 56, 6366–6371. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Han, Y.; Fu, H.; Liu, M.; Wu, J.; Chen, X.; Zhang, S.; Chen, Y. Construction and expression of sTRAIL-melittin combining enhanced anticancer activity with antibacterial activity in Escherichia coli. Appl. Microbiol. Biotechnol. 2013, 97, 2877–2884. [Google Scholar] [CrossRef] [PubMed]
- Falco, A.; Barrajón-Catalán, E.; Menéndez-Gutiérrez, M.P.; Coll, J.; Micol, V.; Estepa, A. Melittin-loaded immunoliposomes against viral surface proteins, a new approach to antiviral therapy. Antivir. Res. 2013, 97, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Son, D.J.; Lee, J.W.; Lee, Y.H.; Song, H.S.; Lee, C.K.; Hong, J.T. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 2007, 115, 246–270. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, P.; Neuner, A.; Nolkemper, S.; Zundel, C.; Nowack, H.; Sensch, K.H.; Reichling, J. Antiviral activity and mode of action of propolis extracts and selected compounds. Phytother. Res. 2010, 24 (Suppl. 1), 20–28. [Google Scholar] [CrossRef] [PubMed]
- Rasul, A.; Millimouno, F.M.; Ali Eltayb, W.; Ali, M.; Li, J.; Li, X. Pinocembrin: A novel natural compound with versatile pharmacological and biological activities. Biomed. Res. Int. 2013. [Google Scholar] [CrossRef]
- Jung, B.G.; Lee, J.A.; Park, S.B.; Hyun, P.M.; Park, J.K.; Suh, G.H.; Lee, B.J. Immunoprophylactic effects of administering honeybee (Apis melifera) venom spray against Salmonella Gallinarum in broiler chicks. J. Vet. Med. Sci. 2013, 75, 1287–1295. [Google Scholar] [CrossRef] [PubMed]
- Nuemann, E.J.; Kliebenstein, J.B.; Johnson, C.D.; Mabry, J.W.; Bush, E.J.; Seitzinger, A.H.; Green, A.L.; Zimmerman, J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. JAVMA 2005, 227, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Giménez-Lirola, L.G.; Halbur, P.G. Polymicrobial respiratory disease in pigs. Anim. Health. Res. Rev. 2011, 12, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Laguna, J.; Salguero, F.J.; Pallarés, F.J.; Carrasco, L. Immunopathogenesis of porcine reproductive and respiratory syndrome in the respiratory tract of pigs. Vet. J. 2013, 195, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, J.; Czerkinsky, C. Mucosal immunity and vaccines. Nat. Med. 2005, 11 (Suppl. 4), S45–S53. [Google Scholar] [CrossRef] [PubMed]
- Renukaradhya, G.J.; Dwivedi, V.; Manickam, C.; Binjawadagi, B.; Benfield, D. Mucosal vaccines to prevent porcine reproductive and respiratory syndrome: a new perspective. Anim. Health. Res. Rev. 2012, 13, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhao, M.; Yang, R.; Zhang, Z.; Li, Y.; Wang, J. Effect of dietary nucleotides on immune function in Balb/C mice. Int. Immunopharmacol. 2013, 17, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, V.; Manickam, C.; Binjawadagi, B.; Linhares, D.; Murtaugh, M.P.; Renukaradhya, G.J. Evaluation of immune responses to porcine reproductive and respiratory syndrome virus in pigs during early stage of infection under farm conditions. Virol. J. 2012, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, V.; Manickam, C.; Binjawadagi, B.; Renukaradhya, G.J. PLGA nanoparticle entrapped killed porcine reproductive and respiratory syndrome virus vaccine helps in viral clearance in pigs. Vet. Microbiol. 2013, 166, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Barranco, I.; Gómez-Laguna, J.; Rodríguez-Gómez, I.M.; Quereda, J.J.; Salguero, F.J.; Pallarés, F.J.; Carrasco, L. Immunohistochemical expression of IL-12, IL-10, IFN-α and IFN-γ in lymphoid organs of porcine reproductive and respiratory syndrome virus-infected pigs. Vet. Immunol. Immunopathol. 2012, 149, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Charerntantanakul, W.; Yamkanchoo, S.; Kasinrerk, W. Plasmids expressing porcine interferon gamma up-regulate pro-inflammatory cytokine and co-stimulatory molecule expression which are suppressed by porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2013, 153, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Shi, K.C.; Guo, X.; Ge, X.N.; Liu, Q.; Yang, H.C. Cytokine mRNA expression profiles in peripheral blood mononuclear cells from piglets experimentally co-infected with porcine reproductive and respiratory syndrome virus and porcine circovirus type 2. Vet. Microbiol. 2010, 140, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Laguna, J.; Salguero, F.J.; Barranco, I.; Pallarés, F.J.; Rodríguez-Gómez, I.M.; Bernabé, A.; Carrasco, L. Cytokine expression by macrophages in the lung of pigs infected with the porcine reproductive and respiratory syndrome virus. J. Comp. Pathol. 2010, 142, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, M.; Darwich, L.; Díaz, I.; de la Torre, E.; Pujols, J.; Martín, M.; Inumaru, S.; Cano, E.; Domingo, M.; Montoya, M.; et al. Cytokine profiles and phenotype regulation of antigen presenting cells by genotype-I porcine reproductive and respiratory syndrome virus isolates. Vet. Res. 2011, 42, 9. [Google Scholar] [CrossRef] [PubMed]
- Olvera, A.; Sibila, M.; Calsamiglia, M.; Segalés, J.; Domingo, M. Comparison of porcine circovirus type 2 load in serum quantified by a real time PCR in postweaning multisystemic wasting syndrome and porcine dermatitis and nephropathy syndrome naturally affected pigs. J. Virol. Method. 2004, 117, 75–80. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.B.; Chan, W.H.; Chaung, H.C.; Lien, Y.; Wu, C.C.; Huang, Y.L. Real-time PCR for quantitation of porcine reproductive and respiratory syndrome virus and porcine circovirus type 2 in naturally-infected and challenged pigs. J. Virol. Method. 2005, 124, 11–19. [Google Scholar] [CrossRef]
- Opriessnig, T.; Thacker, E.L.; Yu, S.; Fenaux, M.; Meng, X.J.; Halbur, P.G. Experimental reproduction of postweaning multisystemic wasting syndrome in pigs by dual infection with Mycoplasma hyopneumoniae and porcine circovirus type 2. Vet. Pathol. 2004, 41, 624–640. [Google Scholar] [CrossRef] [PubMed]
- Manickam, C.; Dwivedi, V.; Patterson, R.; Papenfuss, T.; Renukaradhya, G.J. Porcine reproductive and respiratory syndrome virus induces pronounced immune modulatory responses at mucosal tissues in the parental vaccine strain VR2332 infected pigs. Vet. Microbiol. 2013, 162, 68–77. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-A.; Kim, Y.-M.; Hyun, P.-M.; Jeon, J.-W.; Park, J.-K.; Suh, G.-H.; Jung, B.-G.; Lee, B.-J. Honeybee (Apis mellifera) Venom Reinforces Viral Clearance during the Early Stage of Infection with Porcine Reproductive and Respiratory Syndrome Virus through the Up-Regulation of Th1-Specific Immune Responses. Toxins 2015, 7, 1837-1853. https://doi.org/10.3390/toxins7051837
Lee J-A, Kim Y-M, Hyun P-M, Jeon J-W, Park J-K, Suh G-H, Jung B-G, Lee B-J. Honeybee (Apis mellifera) Venom Reinforces Viral Clearance during the Early Stage of Infection with Porcine Reproductive and Respiratory Syndrome Virus through the Up-Regulation of Th1-Specific Immune Responses. Toxins. 2015; 7(5):1837-1853. https://doi.org/10.3390/toxins7051837
Chicago/Turabian StyleLee, Jin-A, Yun-Mi Kim, Pung-Mi Hyun, Jong-Woon Jeon, Jin-Kyu Park, Guk-Hyun Suh, Bock-Gie Jung, and Bong-Joo Lee. 2015. "Honeybee (Apis mellifera) Venom Reinforces Viral Clearance during the Early Stage of Infection with Porcine Reproductive and Respiratory Syndrome Virus through the Up-Regulation of Th1-Specific Immune Responses" Toxins 7, no. 5: 1837-1853. https://doi.org/10.3390/toxins7051837
APA StyleLee, J.-A., Kim, Y.-M., Hyun, P.-M., Jeon, J.-W., Park, J.-K., Suh, G.-H., Jung, B.-G., & Lee, B.-J. (2015). Honeybee (Apis mellifera) Venom Reinforces Viral Clearance during the Early Stage of Infection with Porcine Reproductive and Respiratory Syndrome Virus through the Up-Regulation of Th1-Specific Immune Responses. Toxins, 7(5), 1837-1853. https://doi.org/10.3390/toxins7051837