
A Novel Peptide-Binding Motifs Inference Approach to Understand Deoxynivalenol Molecular Toxicity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
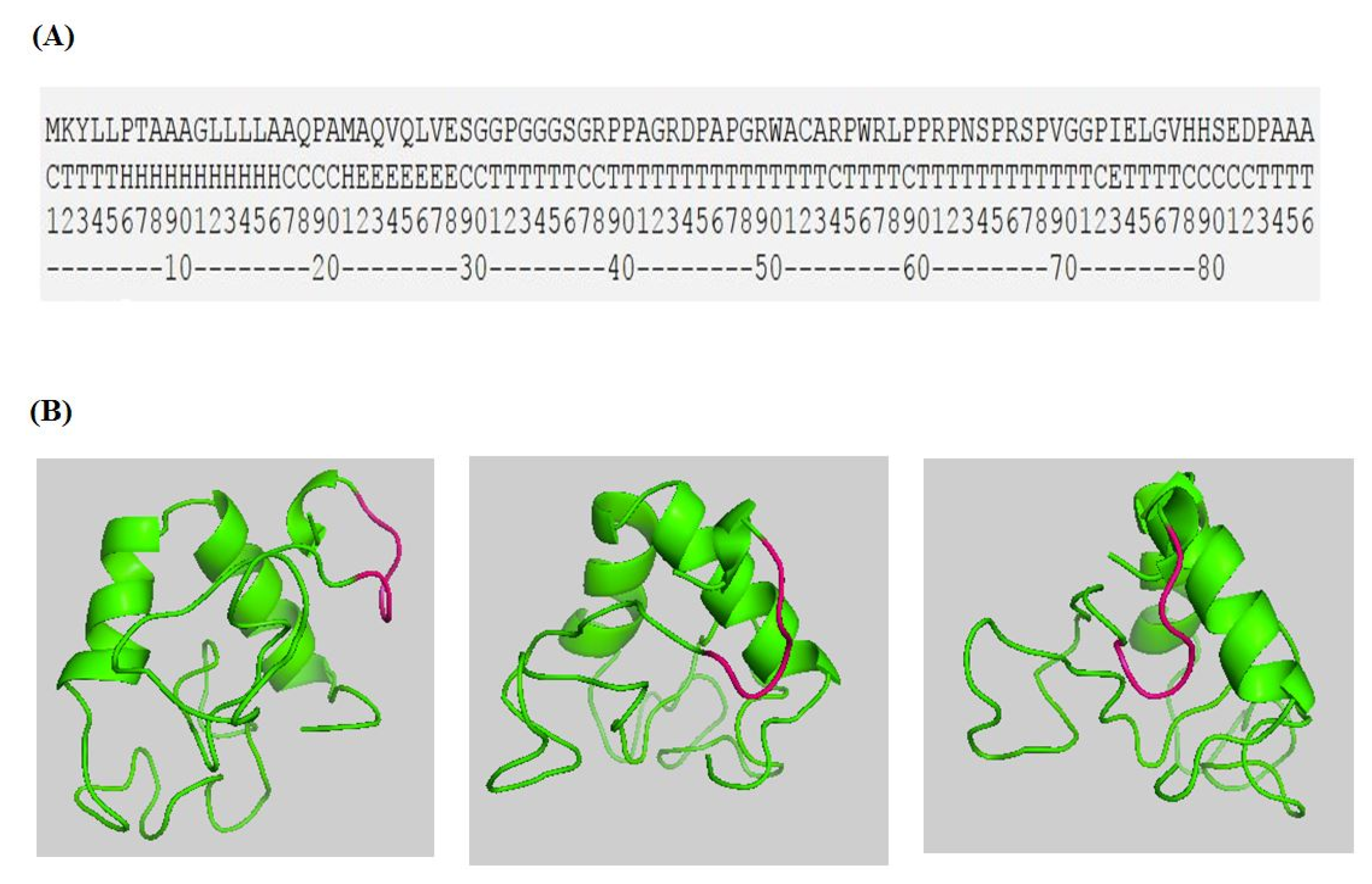
:1. Introduction
2. Cellular Toxicity of DON
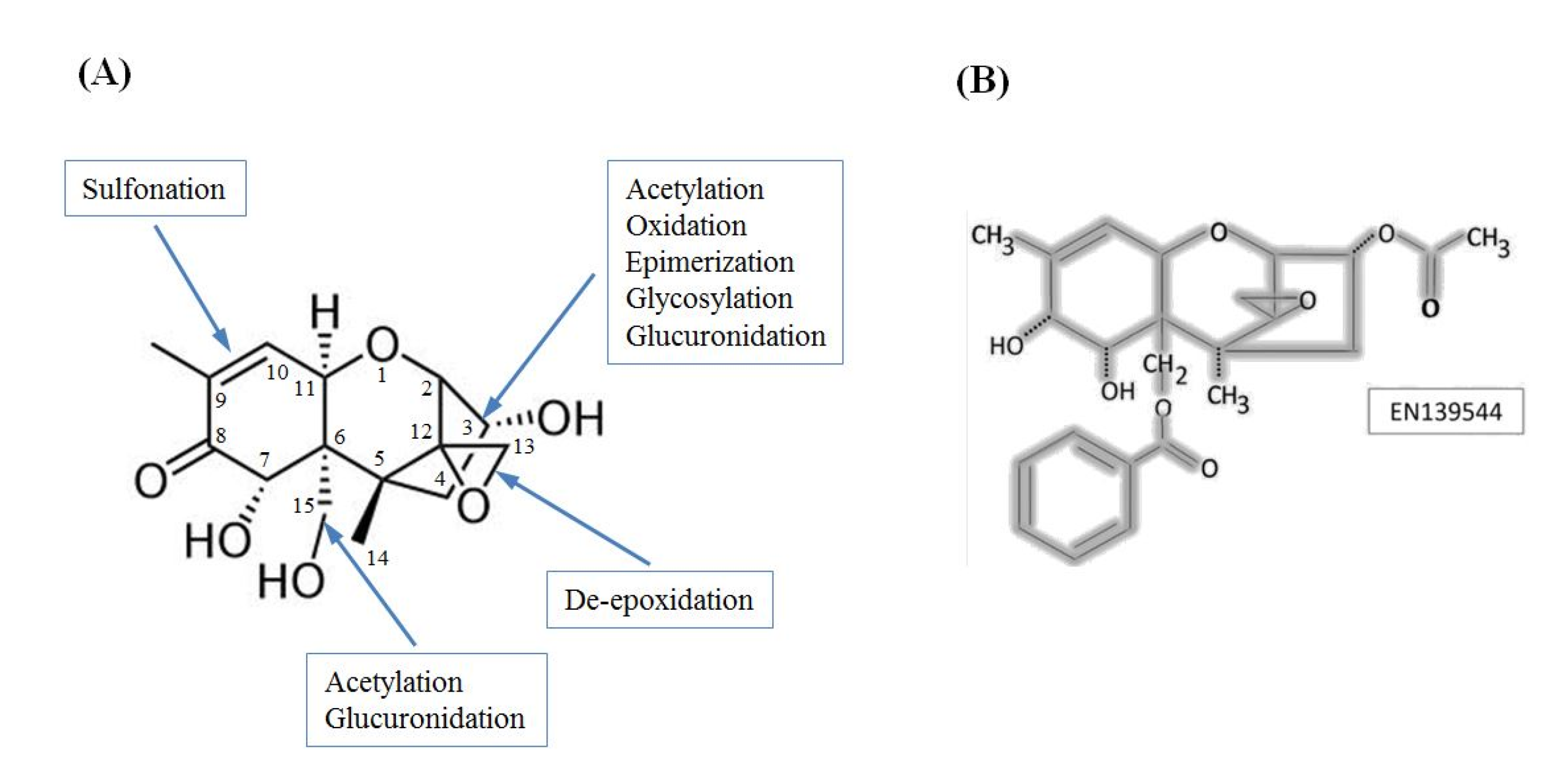
3. The Influence of DON Chemical Groups on its Toxicity


4. Exploring DON Interactions with Cellular Targets



5. Mining Genomes and Proteomes for DON Binding Consensus Motifs
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, X.; Zhao, L.; Fan, Y.; Jia, Y.; Sun, L.; Ma, S.; Ji, C.; Ma, Q.; Zhang, J. Occurrence of Mycotoxins in Feed Ingredients and Complete Feeds Obtained from the Beijing Region of China. J. Anim. Sci. Biotechnol. 2014, 5, 37. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, I.; Naehrer, K. A Three-Year Survey on the Worldwide Occurrence of Mycotoxins in Feedstuffs and Feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Savi, G.D.; Piacentini, K.C.; Tibola, C.S.; Scussel, V.M. Mycoflora and Deoxynivalenol in Whole Wheat Grains (Triticum Aestivum l.) from Southern Brazil. Food Addit. Contam. Part B Surveill. 2014, 7, 232–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratz, S.W.; Richardson, A.J.; Duncan, G.; Holtrop, G. Annual Variation of Dietary Deoxynivalenol Exposure during Years of Different Fusarium Prevalence: A Pilot Biomonitoring Study. Food Addit. Contam. Part A 2014, 31, 1579–1585. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M. From the Gut to the Brain: Journey and Pathophysiological Effects of the Food-Associated Trichothecene Mycotoxin Deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public Health Impacts of Foodborne Mycotoxins. Annu. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Sirot, V.; Fremy, J.M.; Leblanc, J.C. Dietary Exposure to Mycotoxins and Health Risk Assessment in the Second French Total Diet Study. Food Chem. Toxicol. 2013, 52, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Raad, F.; Nasreddine, L.; Hilan, C.; Bartosik, M.; Parent-Massin, D. Dietary Exposure to Aflatoxins, Ochratoxin A and Deoxynivalenol from a Total Diet Study in an Adult Urban Lebanese Population. Food Chem. Toxicol. 2014, 73, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Dwivedi, P.D.; Pandey, H.P.; Das, M. Role of Oxidative Stress in Deoxynivalenol Induced Toxicity. Food Chem. Toxicol. 2014, 72C, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Saint-Cyr, M.J.; Perrin-Guyomard, A.; Houee, P.; Rolland, J.G.; Laurentie, M. Evaluation of an Oral Subchronic Exposure of Deoxynivalenol on the Composition of Human Gut Microbiota in a Model of Human Microbiota-Associated Rats. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Wache, Y.J.; Valat, C.; Postollec, G.; Bougeard, S.; Burel, C.; Oswald, I.P.; Fravalo, P. Impact of Deoxynivalenol on the Intestinal Microflora of Pigs. Int. J. Mol. Sci. 2009, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, M.S.; Roux, J.; Mounien, L.; Dallaporta, M.; Troadec, J.D. Advances in Deoxynivalenol Toxicity Mechanisms: The Brain as a Target. Toxins 2012, 4, 1120–1138. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Bates, M.A.; Bursian, S.J.; Flannery, B.; Zhou, H.R.; Link, J.E.; Zhang, H.; Pestka, J.J. Peptide yy3-36 and 5-Hydroxytryptamine Mediate Emesis Induction by the Trichothecene Deoxynivalenol (Vomitoxin). Toxicol. Sci. 2013. [Google Scholar] [CrossRef]
- Yang, W.; Yu, M.; Fu, J.; Bao, W.; Wang, D.; Hao, L.; Yao, P.; Nüssler, A.K.; Yan, H.; Liu, L. Deoxynivalenol Induced Oxidative Stress and Genotoxicity in Human Peripheral Blood Lymphocytes. Food Chem. Toxicol. 2014, 64, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Takakura, N.; Nesslany, F.; Fessard, V.; Le Hegarat, L. Absence of in vitro Genotoxicity Potential of the Mycotoxin Deoxynivalenol in Bacteria and in Human tk6 and Heparg Cell Lines. Food Chem. Toxicol. 2014, 66, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Videmann, B.; Tep, J.; Cavret, S.; Lecoeur, S. Epithelial Transport of Deoxynivalenol: Involvement of Human P-Glycoprotein (abcb1) and Multidrug Resistance-Associated Protein 2 (abcc2). Food Chem. Toxicol. 2007, 45, 1938–1947. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Toxicity, Mechanisms and Animal Health Risks. Anim. Feed Sci. Technol. 2007, 137, 283–298. [Google Scholar] [CrossRef]
- Rotter, B.A. Invited Review: Toxicology of Deoxynivalenol (Vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Shifrin, V.I.; Anderson, P. Trichothecene Mycotoxins Trigger a Ribotoxic Stress Response that Activates C-Jun N-Terminal Kinase and p38 Mitogen-Activated Protein Kinase and Induces Apoptosis. J. Biol. Chem. 1999, 274, 13985–13992. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Tsybulskyy, D.; Lucioli, J.; Laffitte, J.; Callu, P.; Lyazhri, F.; Grosjean, F.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. Toxicity of Deoxynivalenol and its Acetylated Derivatives on the Intestine: Differential Effects on Morphology, Barrier Function, Tight Junction Proteins, and Mitogen-Activated Protein Kinases. Toxicol. Sci. 2012, 130, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Middlebrook, J.L.; Leatherman, D.L. Binding of t-2 Toxin to Eukaryotic Cell Ribosomes. Biochem. Pharmacol. 1989, 38, 3103–3110. [Google Scholar] [CrossRef]
- Schmeits, P.C.; Katika, M.R.; Peijnenburg, A.A.; van Loveren, H.; Hendriksen, P.J. DON Shares a Similar Mode of Action as the Ribotoxic Stress Inducer Anisomycin while TBTO Shares ER Stress Patterns with the ER Stress Inducer Thapsigargin based on Comparative Gene Expression Profiling in Jurkat T Cells. Toxicol. Lett. 2014, 224, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.; Gray, J.S.; Li, M.; Vines, L.; Kim, J.; Pestka, J.J. Hematopoietic Cell Kinase Associates with the 40s Ribosomal Subunit and Mediates the Ribotoxic Stress Response to Deoxynivalenol in Mononuclear Phagocytes. Toxicol. Sci. 2010, 115, 444–452. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Zhou, H.R.; Pestka, J.J. Targets and Intracellular Signaling Mechanisms for Deoxynivalenol-Induced Ribosomal RNA Cleavage. Toxicol. Sci. 2012, 127, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Cundliffe, E.; Cannon, M.; Davies, J. Mechanism of Inhibition of Eukaryotic Protein Synthesis by Trichothecene Fungal Toxins. Proc. Natl. Acad. Sci. USA 1974, 71, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.-M.; Campbell, I.; McLaughlin, C.; Vaughan, M. Binding of Trichodermin to Mammalian Ribosomes and its Inhibition by Other 12,13-Epoxytrichothecenes. Mol. and Cell. Biochem. 1974, 3, 215–219. [Google Scholar] [CrossRef]
- Wu, W.; He, K.; Zhou, H.R.; Berthiller, F.; Adam, G.; Sugita-Konishi, Y.; Watanabe, M.; Krantis, A.; Durst, T.; Zhang, H.; et al. Effects of Oral Exposure to Naturally-Occurring and Synthetic Deoxynivalenol Congeners on Proinflammatory Cytokine and Chemokine MRNA Expression in the Mouse. Toxicol. Appl. Pharmacol. 2014, 278, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of Action, Human Exposure, and Toxicological Relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Roh, Y.S.; Kim, J.; Lim, C.W.; Kim, B. Differential Immune Modulation by Deoxynivalenol (Vomitoxin) in Mice. Toxicol. Lett. 2013, 221, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J.; Zhou, H.R.; Moon, Y.; Chung, Y.J. Cellular and Molecular Mechanisms for Immune Modulation by Deoxynivalenol and Other Trichothecenes: Unraveling a Paradox. Toxicol. Lett. 2004, 153, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Hirano, S.; Kataoka, T. Deoxynivalenol Induces Ectodomain Shedding of TNF Receptor 1 and thereby Inhibits the TNF-α-induced NF-κB Signaling Pathway. Eur. J. Pharmacol. 2013, 701, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Nougayrède, J.-P.; Del Rio, J.-C.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.-P.; Kolf-Clauw, M.; Oswald, I.P. The Food Contaminant Deoxynivalenol, Decreases Intestinal Barrier Permeability and Reduces Claudin Expression. Toxicol. Appl. Pharmacol. 2009, 237, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wu, W.; Wang, X.; He, C.; Yue, H.; Ren, Z.; Zhang, H. Inhibitory Effects of Deoxynivalenol on Gastric Secretion in Rats. J. Food Prot. 2014, 77, 1367–1371. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Fantini, J. Some Food-Associated Mycotoxins as Potential Risk Factors in Humans Predisposed to Chronic Intestinal Inflammatory Diseases. Toxicon 2010, 56, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhou, H.R.; He, K.; Pan, X.; Sugita-Konishi, Y.; Watanabe, M.; Zhang, H.; Pestka, J.J. Role of Cholecystokinin in Anorexia Induction Following Oral Exposure to the 8-Ketotrichothecenes Deoxynivalenol, 15-Acetyldeoxynivalenol, 3-Acetyldeoxynivalenol, Fusarenon X, and Nivalenol. Toxicol. Sci. 2014, 138, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Näslund, E.; Hellström, P.M. Appetite Signaling: From Gut Peptides and Enteric Nerves to Brain. Physiol. Behav. 2007, 92, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Noetzel, S.; Stengel, A.; Inhoff, T.; Goebel, M.; Wisser, A.S.; Bannert, N.; Wiedenmann, B.; Klapp, B.F.; Tache, Y.; Monnikes, H.; et al. CCK-8S Activates c-Fos in a Dose-Dependent Manner in Nesfatin-1 Immunoreactive Neurons in the Paraventricular Nucleus of the Hypothalamus and in the Nucleus of the Solitary Tract of the Brainstem. Regul. Pept. 2009, 157, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Flannery, B.M.; Clark, E.S.; Pestka, J.J. Anorexia Induction by the Trichothecene Deoxynivalenol (Vomitoxin) is Mediated by the Release of the Gut Satiety Hormone Peptide YY. Toxicol. Sci. 2012, 130, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Xiao, H.; Ren, W.; Yin, J.; Tan, B.; Liu, G.; Li, L.; Nyachoti, C.M.; Xiong, X.; Wu, G. Therapeutic Effects of Glutamic Acid in Piglets Challenged with Deoxynivalenol. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Lessard, M.; Savard, C.; Deschene, K.; Lauzon, K.; Pinilla, V.A.; Gagnon, C.A.; Lapointe, J.; Guay, F.; Chorfi, Y. Impact of Deoxynivalenol (DON) Contaminated Feed on Intestinal Integrity and Immune Response in Swine. Food Chem. Toxicol. 2015, 80, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Del Regno, M.; Adesso, S.; Popolo, A.; Quaroni, A.; Autore, G.; Severino, L.; Marzocco, S. Nivalenol Induces Oxidative Stress and Increases Deoxynivalenol Pro-Oxidant Effect in Intestinal Epithelial Cells. Toxicol. Appl. Pharmacol. 2015, 285, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Bianco, G.; Fontanella, B.; Severino, L.; Quaroni, A.; Autore, G.; Marzocco, S. Nivalenol and Deoxynivalenol Affect Rat Intestinal Epithelial Cells: A Concentration Related Study. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.C.; Hou, Y.J.; Han, J.; Liu, H.L.; Cui, X.S.; Kim, N.H.; Sun, S.C. Effect of Mycotoxin-Containing Diets on Epigenetic Modifications of Mouse Oocytes by Fluorescence Microscopy Analysis. Microsc. Microanal. 2014, 20, 1158–1161. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer. Some Naturally-Occurring Substances: Some Food Items and Constituents, Heterocyclic Amines and Mycotoxins, Lyon, France, 1993; Volume 56, 397–444.
- Zhou, T.; He, J.; Gong, J. Microbial transformation of trichothecene mycotoxins. World Mycotoxin J. 2008, 1, 23–30. [Google Scholar] [CrossRef]
- Young, J.C.; Zhou, T.; Yu, H.; Zhu, H.; Gong, J. Degradation of Trichothecene Mycotoxins by Chicken Intestinal Microbes. Food Chem. Toxicol. 2007, 45, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Sundstøl Eriksen, G.; Pettersson, H.; Lundh, T. Comparative Cytotoxicity of Deoxynivalenol, Nivalenol, their Acetylated Derivatives and De-Epoxy Metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Dänicke, S.; Hegewald, A.-K.; Kahlert, S.; Kluess, J.; Rothkötter, H.J.; Breves, G.; Döll, S. Studies on the Toxicity of Deoxynivalenol (DON), Sodium Metabisulfite, DON-Sulfonate (DONS) and De-Epoxy-DON for Porcine Peripheral Blood Mononuclear Cells and the Intestinal Porcine Epithelial Cell Lines IPEC-1 and IPEC-J2, and on Effects of DON and DONS on Piglets. Food Chem. Toxicol. 2010, 48, 2154–2162. [Google Scholar] [PubMed]
- Shima, J.; Takase, S.; Takahashi, Y.; Iwai, Y.; Fujimoto, H.; Yamazaki, M.; Ochi, K. Novel Detoxification of the Trichothecene Mycotoxin Deoxynivalenol by a Soil Bacterium Isolated by Enrichment Culture. Appl. Environ. Microbiol. 1997, 63, 3825–3830. [Google Scholar] [PubMed]
- Ikunaga, Y.; Sato, I.; Grond, S.; Numaziri, N.; Yoshida, S.; Yamaya, H.; Hiradate, S.; Hasegawa, M.; Toshima, H.; Koitabashi, M.; et al. Nocardioides sp. Strain WSN05-2, Isolated from a Wheat Field, Degrades Deoxynivalenol, Producing the Novel Intermediate 3-epi-Deoxynivalenol. Appl. Microbiol. Biotechnol. 2011, 89, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; He, J. Bacterial Isolate, Methods of Isolating Bacterial Isolates and Methods for Detoxification of Trichothecene Mycotoxins. U.S. Patent US13,500,239, 6 October 2010. [Google Scholar]
- Karlovsky, P. Biological Detoxification of the Mycotoxin Deoxynivalenol and its Use in Genetically Engineered Crops and Feed Additives. Appl. Microbiol. Biotechnol. 2011, 91, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Nagl, V.; Woechtl, B.; Schwartz-Zimmermann, H.E.; Hennig-Pauka, I.; Moll, W.D.; Adam, G.; Berthiller, F. Metabolism of the Masked Mycotoxin Deoxynivalenol-3-Glucoside in Pigs. Toxicol. Lett. 2014, 229, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Poppenberger, B.; Berthiller, F.; Lucyshyn, D.; Sieberer, T.; Schuhmacher, R.; Krska, R.; Kuchler, K.; Glossl, J.; Luschnig, C.; Adam, G. Detoxification of the Fusarium Mycotoxin Deoxynivalenol by a UDP-Glucosyltransferase from Arabidopsis Thaliana. J. Biol. Chem. 2003, 278, 47905–47914. [Google Scholar] [CrossRef] [PubMed]
- Maul, R.; Warth, B.; Schebb, N.H.; Krska, R.; Koch, M.; Sulyok, M. In vitro Glucuronidation Kinetics of Deoxynivalenol by Human and Animal Microsomes and Recombinant Human UGT Enzymes. Arch. Toxicol. 2014, 89, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Murphy, P.; Cunnick, J.; Hendrich, S. Synthesis and Characterization of Deoxynivalenol Glucuronide: Its Comparative Immunotoxicity with Deoxynivalenol. Food Chem. Toxicol. 2007, 45, 1846–1855. [Google Scholar] [CrossRef] [PubMed]
- Fruhmann, P.; Weigl-Pollack, T.; Mikula, H.; Wiesenberger, G.; Adam, G.; Varga, E.; Berthiller, F.; Krska, R.; Hametner, C.; Frohlich, J. Methylthiodeoxynivalenol (MTD): Insight into the Chemistry, Structure and Toxicity of Thia-Michael Adducts of Trichothecenes. Org. Biomol. Chem. 2014, 12, 5144–5150. [Google Scholar] [CrossRef] [PubMed]
- Kadota, T.; Furusawa, H.; Hirano, S.; Tajima, O.; Kamata, Y.; Sugita-Konishi, Y. Comparative Study of Deoxynivalenol, 3-Acetyldeoxynivalenol, and 15-Acetyldeoxynivalenol on Intestinal Transport and IL-8 Secretion in the Human Cell Line Caco-2. Toxicol. in Vitro 2013, 27, 1888–1895. [Google Scholar] [CrossRef] [PubMed]
- Danicke, S.; Kersten, S.; Valenta, H.; Breves, G. Inactivation of Deoxynivalenol-Contaminated Cereal Grains with Sodium Metabisulfite: A Review of Procedures and Toxicological Aspects. Mycotoxin Res. 2012, 28, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.; Puel, O.; Oswald, I.P. Toxicological Interactions between the Mycotoxins Deoxynivalenol, Nivalenol and their Acetylated Derivatives in Intestinal Epithelial Cells. Arch. Toxicol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, I.T.; Park, J.H.; Choi, P.S.; Saier, M.H., Jr. A Family of Gram-Negative Bacterial Outer Membrane Factors that Function in the Export of Proteins, Carbohydrates, Drugs and Heavy Metals from Gram-Negative Bacteria. FEMS Microbiol. Lett. 1997, 156, 1–8. [Google Scholar] [CrossRef]
- Garvey, G.S.; McCormick, S.P.; Rayment, I. Structural and Functional Characterization of the TRI101 Trichothecene 3-O-Acetyltransferase from Fusarium Sporotrichioides and Fusarium Graminearum: Kinetic insights to Combating Fusarium Head Blight. J. Biol. Chem. 2008, 283, 1660–1669. [Google Scholar] [CrossRef] [PubMed]
- Khatibi, P.A.; Newmister, S.A.; Rayment, I.; McCormick, S.P.; Alexander, N.J.; Schmale, D.G., 3rd. Bioprospecting for Trichothecene 3-O-Acetyltransferases in the Fungal Genus Fusarium Yields Functional Enzymes with Different Abilities to Modify the Mycotoxin Deoxynivalenol. Appl. Environ. Microbiol. 2011, 77, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; Proctor, R.H.; Dyer, R.B.; Plattner, R.D. Characterization of a Fusarium 2-Gene Cluster Involved in Trichothecene C-8 Modification. J. Agric. Food Chem. 2003, 51, 7936–7944. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Matsumoto, G.; Shingu, Y.; Yoneyama, K.; Yamaguchi, I. The Mystery of the Trichothecene 3-O-Acetyltransferase Gene. Analysis of the Region around TRI101 and Characterization of its Homologue from Fusarium Sporotrichioides. FEBS Lett. 1998, 435, 163–168. [Google Scholar] [CrossRef]
- Kimura, M.; Kaneko, I.; Komiyama, M.; Takatsuki, A.; Koshino, H.; Yoneyama, K.; Yamaguchi, I. Trichothecene 3-O-Acetyltransferase Protects Both the Producing Organism and Transformed Yeast from Related Mycotoxins. Cloning and Characterization of TRI101. J. Biol. Chem. 1998, 273, 1654–1661. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Sato, I.; Ishizaka, M.; Yoshida, S.; Koitabashi, M.; Yoshida, S.; Tsushima, S. Bacterial Cytochrome P450 System Catabolizing the Fusarium Toxin Deoxynivalenol. Appl. Environ. Microbiol. 2013, 79, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, W.; Boddu, J.; Shin, S.; Poppenberger, B.; Berthiller, F.; Lemmens, M.; Muehlbauer, G.J.; Adam, G. Validation of a Candidate Deoxynivalenol-Inactivating UDP-Glucosyltransferase from Barley by Heterologous Expression in Yeast. Mol. Plant-Microbe Interact. 2010, 23, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Tu, Z.; Xu, Y.; He, Q.; Fu, J.; Liu, X.; Tao, Y. Isolation and Characterisation of Deoxynivalenol Affinity Binders from a Phage Display Library Based on Single-Domain Camelid Heavy Chain Antibodies (VHHS). Food Agric. Immunol. 2012, 23, 123–131. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y. Ab initio Protein Structure Assembly Using Continuous Structure Fragments and Optimized Knowledge-Based Force Field. Proteins 2012, 80, 1715–1735. [Google Scholar] [CrossRef] [PubMed]
- Holm, L.; Rosenstrom, P. Dali Server: Conservation Mapping in 3D. Nucleic Acids Res. 2010, 38, W545–W549. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Y.I.; Lepp, D.; He, J.; Zhou, T. Draft Genome Sequences of Devosia sp. Strain 17-2-E-8 and Devosia Riboflavina Strain IFO13584. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, Y.I.; Watts, C.; Li, X.-Z.; Zhou, T. A Novel Peptide-Binding Motifs Inference Approach to Understand Deoxynivalenol Molecular Toxicity. Toxins 2015, 7, 1989-2005. https://doi.org/10.3390/toxins7061989
Hassan YI, Watts C, Li X-Z, Zhou T. A Novel Peptide-Binding Motifs Inference Approach to Understand Deoxynivalenol Molecular Toxicity. Toxins. 2015; 7(6):1989-2005. https://doi.org/10.3390/toxins7061989
Chicago/Turabian StyleHassan, Yousef I., Christena Watts, Xiu-Zhen Li, and Ting Zhou. 2015. "A Novel Peptide-Binding Motifs Inference Approach to Understand Deoxynivalenol Molecular Toxicity" Toxins 7, no. 6: 1989-2005. https://doi.org/10.3390/toxins7061989
APA StyleHassan, Y. I., Watts, C., Li, X.-Z., & Zhou, T. (2015). A Novel Peptide-Binding Motifs Inference Approach to Understand Deoxynivalenol Molecular Toxicity. Toxins, 7(6), 1989-2005. https://doi.org/10.3390/toxins7061989