
Activities and Effects of Ergot Alkaloids on Livestock Physiology and Production

Abstract
:
1. Introduction
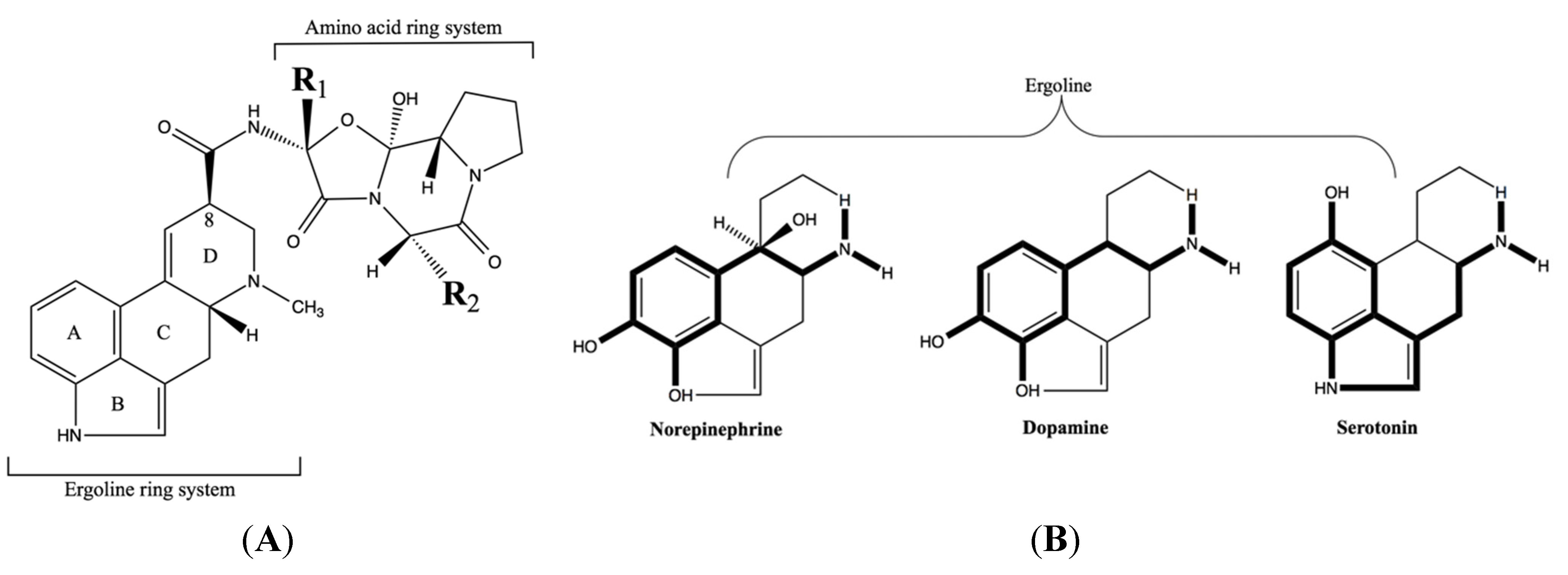
2. Ergot Alkaloid Activity

3. Gangrenous Ergotism

{kind=link}
{kind=link}
| Alkaloid | Intake/dose (mg/kg body weight) | Animal (affected/exposed) | Effect | Source |
|---|---|---|---|---|
| Ethanolic extract 1 | Not determined | Cattle (5/10) | Lameness, swelling and discoloration of coronary bands, discoloration of tail tip | [45] |
| Ergovaline | 0.016 2 | Steer (1/2) | Visible inflammation of coronary band and elongation of rear hooves | [46] |
| Ergovaline | 0.011 2 | Ewe (1/14) | Lameness and visible inflammation around coronary bands | [46] |
| Ergovaline | 0.009 3 | Cow (1/60) | Developed lameness diagnosed as fescue foot after 55 d on treatment, complete recovery after removal from study | [47] |
| Ergotoxine 4 | 25.0 5 | Rat (20%) | Tail gangrene was observed 5 to 7 d following a single i.p. dose | [48] |
| Ergotamine | 1.0 6 | Sheep (4/6) | Four out 6 died after 10d; tongue necrosis and hemorrhages around fetlock and metatarsal regions | [49] |
3.1. Blood Flow
3.2. Pathogenesis
4. Decreased Animal Productivity
4.1. Hyperthermia
4.2. Decreased Weight Gain and Intake
| Ergovaline Intake (mg/kg BW0.75) 1 | Concentration (mg/kg DM) | Animal | Effect | Source |
|---|---|---|---|---|
| 0.008 2 | 0.120 3 | Steers | Decreased dry matter intake and digestibility, no difference in NDF digestibility | [70] |
| 0.021 | 0.285 | Steer calves | Decreased dry matter intake and digestibility, no difference in NDF digestibility, increased water intake | [66] |
| 0.044 | 0.475 | Steers | Intake equal, decrease in rumen fill observed with alkaloid, passage rate not affected | [78] |
| 0.051 | 4.1 | Steers | Intake was fixed, increase rumen fill observed (% DM) | [79] |
| 0.053 | 1.17 | Wethers | Decreased dry matter intake and digestibility, altered rumen fluid kinetics | [80] |
| 0.057 | 0.96 | Ram lambs | Equal intake, lower rumen fill | [81] |
| 0.059 | 4.45 | Steers | Intake was fixed, increase in rumen fill (% DM) observed and ruminal VFA concentrations, decreased blood flow to rumen | [82] |
| 0.098 | NA4 | Steers | Intake was fixed, increased rumen fill observed (% DM), decreased particulate passage | [75] |
| 0.19 | 3.0 | Wethers | Decreased NDF digestibility, decrease in rumen fluid volume, increased water intake | [83] |
| 0.093 2 | 0.620 5 | Cows | Higher quantities of undegraded protein and NDF digestibility measured at the duodenum, increased ruminal VFA and ammonia nitrogen concentrations | [74] |
4.3. Decreased Reproduction
4.3.1. Lowered Prolactin
4.3.2. Female-Specific Effects
4.3.3. Male-Specific Effects
5. Fat Necrosis
6. Conclusions
Conflicts of Interest
References
- Strickland, J.R.; Looper, M.L.; Matthews, J.C.; Rosenkrans, C.F., Jr.; Flythe, M.D.; Brown, K.R. Board-invited review: St. Anthonyʼs Fire in livestock: Causes, mechanisms, and potential solutions. J. Anim. Sci. 2011, 89, 1603–1626. [Google Scholar] [CrossRef] [PubMed]
- Di Menna, M.E.; Finch, S.C.; Popay, A.J.; Smith, B.L. A review of the Neotyphodium lolii/Lolium perenne symbiosis and its associated effects on animal and plant health, with particular emphasis on ryegrass staggers. N. Z. Vet. J. 2012, 60, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Caldwell-Smith, A.J.; Bakau, W.J.K.; Scrivener, C.J.; Reed, K.F.M.; Blaney, B.J.; Bryden, W.L. Ergot Toxins and Animal Disease in Australia; CABI Publishing: Wallingford, UK, 2007. [Google Scholar]
- Miyazaki, S.; Fukumura, M.; Yoshioka, M.; Yamanaka, N. Detection of endophyte toxins in the imported perennial ryegrass straw. J. Vet. Med. Sci. 2001, 63, 1013–1015. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, S.; Ishizaki, I.; Ishizaka, M.; Kanbara, T.; Ishiguro-Takeda, Y. Lolitrem B residue in fat tissues of cattle consuming endophyte-infected perennial ryegrass straw. J. Vet. Diagn. Investig. 2004, 16, 340–342. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, C.W.; Ahn, G.C.; Park, E.K.; Kim, C.M.; Park, K.K. Ergovaline levels in tall fescue and its effect on performance of lactating cows. Anim. Feed Sci. Technol. 2007, 136, 330–337. [Google Scholar] [CrossRef]
- Alabdouli, K.O.; Blythe, L.L.; Duringer, J.M.; Elkhouly, A.; Kassab, A.; Askar, M.; Mohammed, E.E.; Al-Juboori, A.; Craig, A.M. Physiological effects of endophyte-infected perennial ryegrass straw on female camels in the Middle East. Emir. J. Food Agric. 2013, 26, 82–92. [Google Scholar]
- Lean, I.J. Association between feeding perennial ryegrass (Lolium perenne cultivar Grasslands impact) containing high concentrations of ergovaline, and health and productivity in a herd of lactating dairy cows. Aust Vet. J. 2001, 79, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Blaney, B.J.; McLennan, S.R.; Kidd, J.F.; Connell, J.A.; McKenzie, R.A.; Downing, J.A. Effect of sorghum ergot (Claviceps africana) on the performance of steers (Bos taurus) in a feedlot. Anim. Prod. Sci. 2011, 51, 156. [Google Scholar] [CrossRef]
- Young, C.A.; Hume, D.E.; McCulley, R.L. Forages and pastures symposium: Fungal endophytes of tall fescue and perennial ryegrass: Pasture friend or foe? J. Anim. Sci. 2013, 91, 2379–2394. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Le, T.N.; Ludlow, E.J.; Smith, K.F.; Forster, J.W.; Guthridge, K.M.; Spangenberg, G.C. Characterisation of novel perennial ryegrass host-Neotyphodium endophyte associations. Crop. Pasture Sci. 2013, 64, 716. [Google Scholar] [CrossRef]
- Reed, K.F.M.; Nie, Z.N.; Walker, L.V.; Kearney, G. Fluctuations in the concentration of ergovaline and lolitrem B produced by the wild-type endophyte (Neotyphodium lolii) in perennial ryegrass (Lolium perenne) pasture. Anim. Prod. Sci. 2011, 51, 1098–1108. [Google Scholar] [CrossRef]
- McCulley, R.L.; Bush, L.P.; Carlisle, A.E.; Ji, H.; Nelson, J.A. Warming reduces tall fescue abundance but stimulates toxic alkaloid concentrations in transition zone pastures of the U.S. Front. Chem 2014, 2, 88. [Google Scholar] [CrossRef] [PubMed]
- Aiken, G.E.; Strickland, J.R. Forages and pastures symposium: Managing the tall fescue-fungal endophyte symbiosis for optimum forage-animal production. J. Anim. Sci. 2013, 91, 2369–2378. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, I.J. A Note on the Cause of Tall Fescue Lameness in Cattle. Aust. Vet. J. 1949, 25, 27–28. [Google Scholar] [CrossRef]
- Schmidt, S.P.; Osborn, T.G. Effects of endophyte-infected tall fescue on animal performance. Agric. Ecosyst. Environ. 1993, 44, 233–262. [Google Scholar] [CrossRef]
- Rumsey, T.S.; Stuedemann, J.A.; Wilkinson, S.R.; Williams, D.J. Chemical composition of necrotic fat lesions in beef cows grazing fertilized “Kentucky-31” tall fescue. J. Anim. Sci. 1979, 48, 673–682. [Google Scholar] [PubMed]
- Lyons, P.C.; Plattner, R.D.; Bacon, C.W. Occurrence of peptide and clavine ergot alkaloids in tall fescue grass. Science 1986, 232, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Mace, W.J.; Lunn, K.L.; Kaur, N.; Lloyd-West, C.M. Variation in the expression of ergot alkaloids between individual tillers of perennial ryegrass. Front. Chem. 2014, 2, 107. [Google Scholar] [CrossRef] [PubMed]
- Lea, K.; Smith, L.; Gaskill, C.; Coleman, R.; Smith, S.R. Ergovaline stability in tall fescue based on sample handling and storage methods. Front. Chem. 2014, 2, 76. [Google Scholar] [CrossRef] [PubMed]
- Flieger, M.; Wurst, M.; Shelby, R. Ergot alkaloids—Sources, structures and analytical methods. Folia Microbiol. 1997, 42, 3–30. [Google Scholar] [CrossRef]
- Berde, B.; Stürmer, E. Introduction to the Pharmacology of Ergot Alkaloids and Related Compounds as a Basis of Their Therapeutic Action; Springer: Berlin, Germany, 1978. [Google Scholar]
- Pertz, H.H.; Eckart, E. Ergot Alkaloids And Their Derivatives As Ligands For. Serotoninergic, Dopaminergic, And Adrenergic Receptors; Harwood Academic Publishers: Amsterdam, The Netherlands, 1999. [Google Scholar]
- Goddard, W.A., 3rd; Abrol, R. 3-Dimensional structures of G protein-coupled receptors and binding sites of agonists and antagonists. J. Nutr. 2007, 137, 1528S–1538S. [Google Scholar] [PubMed]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef]
- Klotz, J.L.; Kirch, B.H.; Aiken, G.E.; Bush, L.P.; Strickland, J.R. Effects of selected combinations of tall fescue alkaloids on the vasoconstrictive capacity of fescue-naive bovine lateral saphenous veins. J. Anim. Sci. 2008, 86, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Klotz, J.L.; Kirch, B.H.; Aiken, G.E.; Bush, L.P.; Strickland, J.R. Contractile response of fescue-naive bovine lateral saphenous veins to increasing concentrations of tall fescue alkaloids. J. Anim. Sci. 2010, 88, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.S.; Sachs, N.; Uluer, A.; Glennon, R.A.; Westkaemper, R.B.; Roth, B.L. Differential ergoline and ergopeptine binding to 5-hydroxytryptamine2A receptors: Ergolines require an aromatic residue at position 340 for high affinity binding. Mol. Pharmacol. 1995, 47, 450–457. [Google Scholar] [PubMed]
- Berde, B. Ergot Compounds: A Synopsis; Raven Press: New York, NY, USA, 1980. [Google Scholar]
- Klotz, J.L.; Bush, L.P.; Smith, D.L.; Shafer, W.D.; Smith, L.L.; Arrington, B.C.; Strickland, J.R. Ergovaline-induced vasoconstriction in an isolated bovine lateral saphenous vein bioassay. J. Anim. Sci. 2007, 85, 2330–2336. [Google Scholar] [CrossRef] [PubMed]
- Dyer, D.C. Evidence that ergovaline acts on serotonin receptors. Life Sci. 1993, 53, PL223–PL228. [Google Scholar] [CrossRef]
- Pesqueira, A.; Harmon, D.L.; Branco, A.F.; Klotz, J.L. Bovine lateral saphenous veins exposed to ergopeptine alkaloids do not relax. J. Anim. Sci. 2014, 92, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Schöning, C.; Flieger, M.; Pertz, H.H. Complex interaction of ergovaline with 5-HT2A, 5-HT1B/1D, and alpha1 receptors in isolated arteries of rat and guinea pig. J. Anim. Sci. 2001, 79, 2202–2209. [Google Scholar] [PubMed]
- Egert, A.M.; Kim, D.H.; Schrick, F.N.; Harmon, D.L.; Klotz, J.L. Dietary exposure to ergot alkaloids decreases contractility of bovine mesenteric vasculature. J. Anim. Sci. 2014, 92, 1768–1779. [Google Scholar] [CrossRef] [PubMed]
- Millan, M.J.; Marin, P.; Bockaert, J.; Mannoury la Cour, C. Signaling at G-protein-coupled serotonin receptors: Recent advances and future research directions. Trends Pharmacol. Sci. 2008, 29, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.M.; Brady, A.E.; Nickols, H.H.; Wang, Q.; Limbird, L.E. Membrane trafficking of G protein-coupled receptors. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 559–609. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Liao, S.F.; Strickland, J.R.; Boling, J.A.; Matthews, J.C. Bovine neuronal vesicular glutamate transporter activity is inhibited by ergovaline and other ergopeptines. J. Dairy Sci. 2011, 94, 3331–3341. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.D.; Xue, Y.; Strickland, J.R.; Boling, J.A.; Matthews, J.C. Ergopeptines bromocriptine and ergovaline and the dopamine type-2 receptor inhibitor domperidone inhibit bovine equilibrative nucleoside transporter 1-like activity. J. Agric. Food Chem. 2011, 59, 9691–9699. [Google Scholar] [CrossRef] [PubMed]
- Mulac, D.; Humpf, H.U. Cytotoxicity and accumulation of ergot alkaloids in human primary cells. Toxicology 2011, 282, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Mulac, D.; Huwel, S.; Galla, H.J.; Humpf, H.U. Permeability of ergot alkaloids across the blood-brain barrier in vitro and influence on the barrier integrity. Mol. Nutr. Food Res. 2012, 56, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Stadler, P.A.; Sturmer, E. Comparative studies on the pharmacological properties of stereoisomers of ergotamine and dihydro-ergotamine. Naunyn Schmied. Arch. Pharmakol. 1970, 266, 457–458. [Google Scholar]
- Smith, D.J.; Shappell, N.W. Technical note: Epimerization of ergopeptine alkaloids in organic and aqueous solvents. J. Anim. Sci. 2002, 80, 1616–1622. [Google Scholar] [CrossRef]
- Strickland, J.R.; Aiken, G.E.; Klotz, J.L. Ergot alkaloid induced blood vessel dysfunction contributes to fescue toxicosis. Forage Grazinglands 2009. [Google Scholar] [CrossRef]
- Vuong, P.N.; Berry, C.L. The Pathology of Vessels; Springer-Verlag: Paris, France, 2002; p. 527. [Google Scholar]
- Williams, M.; Shaffer, S.R.; Garner, G.B.; Yates, S.G.; Tookey, H.L.; Kintner, L.D.; Nelson, S.L.; McGinity, J.T. Induction of fescue foot syndrome in cattle by fractionated extracts of toxic fescue hay. Am. J. Vet. Res. 1975, 36, 1353–1357. [Google Scholar] [PubMed]
- Tor-Agbidye, J.; Blythe, L.L.; Craig, A.M. Correlation of endophyte toxins (ergovaline and lolitrem B) with clinical disease: Fescue foot and perennial ryegrass staggers. Vet. Hum. Toxicol. 2001, 43, 140–146. [Google Scholar] [PubMed]
- Merrill, M.L.; Bohnert, D.W.; Harmon, D.L.; Craig, A.M.; Schrick, F.N. The ability of a yeast-derived cell wall preparation to minimize the toxic effects of high-ergot alkaloid tall fescue straw in beef cattle. J. Anim. Sci. 2007, 85, 2596–2605. [Google Scholar] [CrossRef] [PubMed]
- Griffith, R.W.; Grauwiler, J.; Hodel, C.H.; Leist, K.H.; Matter, B. Toxicologic Considerations; Springer Verlag: Berlin, Germany, 1978. [Google Scholar]
- Greatorex, J.C.; Mantle, P.G. Experimental ergotism in sheep. Res. Vet. Sci. 1973, 15, 337–345. [Google Scholar] [PubMed]
- Cross, D.L.; Redmond, L.M.; Strickland, J.R. Equine fescue toxicosis: Signs and solutions. J. Anim. Sci. 1995, 73, 899–908. [Google Scholar] [PubMed]
- McDowell, K.J.; Moore, E.S.; Parks, A.G.; Bush, L.P.; Horohov, D.W.; Lawrence, L.M. Vasoconstriction in horses caused by endophyte-infected tall fescue seed is detected with Doppler ultrasonography. J. Anim. Sci. 2013, 91, 1677–1684. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, D.R.; Miller, W.M.; Seath, D.M.; Yates, S.G.; Tookey, H.L.; Wolff, I.A. Nature of Fescue Toxicity and Progress toward Identification of the Toxic Entity. J. Dairy Sci. 1963, 46, 416–422. [Google Scholar] [CrossRef]
- Walls, J.R.; Jacobson, D.R. Skin temperature and blood flow in the tail of dairy heifers administered extracts of toxic tall fescue. J. Anim. Sci. 1970, 30, 420–423. [Google Scholar] [PubMed]
- Strickland, J.R.; Bailey, E.M.; Abney, L.K.; Oliver, J.W. Assessment of the mitogenic potential of the alkaloids produced by endophyte (Acremonium coenophialum)-infected tall fescue (Festuca arundinacea) on bovine vascular smooth muscle in vitro. J. Anim. Sci. 1996, 74, 1664–1671. [Google Scholar] [PubMed]
- Aiken, G.E.; Kirch, B.H.; Strickland, J.R.; Bush, L.P.; Looper, M.L.; Schrick, F.N. Hemodynamic responses of the caudal artery to toxic tall fescue in beef heifers. J. Anim. Sci. 2007, 85, 2337–2345. [Google Scholar] [CrossRef] [PubMed]
- Aiken, G.E.; Flythe, M.D. Vasoconstrictive responses by the carotid and auricular arteries in goats to ergot alkaloid exposure. Front. Chem. 2014, 2, 101. [Google Scholar] [CrossRef] [PubMed]
- Scottish Agricultural College Veterinary Services. BVD virus causes heavy losses in a Scottish cattle herd. Vet. Rec. 2007, 160, 281–284. [Google Scholar]
- Bush, L.; Boling, J.; Yates, S. Animal Disorders. In Agronomy Monograph 20; Buckner, R.C., Bush, L.P., Eds.; ASA, CSSA, SSSA: Madison, WI, USA, 1979. [Google Scholar]
- EFSA. Scientific Opinion on Ergot Alkaloids in Food and Feed. EFSA J. 2012, 10, 158. [Google Scholar]
- Paterson, J.; Forcherio, C.; Larson, B.; Samford, M.; Kerley, M. The effects of fescue toxicosis on beef cattle productivity. J. Anim. Sci. 1995, 73, 889–898. [Google Scholar] [PubMed]
- Thompson, F.N.; Stuedemann, J.A. Pathophysiology of fescue toxicosis. Agric. Ecosyst. Environ. 1993, 44, 263–281. [Google Scholar] [CrossRef]
- Strickland, J.R.; Oliver, J.W.; Cross, D.L. Fescue toxicosis and its impact on animal agriculture. Vet. Hum. Toxicol. 1993, 35, 454–464. [Google Scholar] [PubMed]
- Waller, J.C. Endophyte Effects on Cattle. In Tall Fescue for the Twenty-first Century; Fribourg, H.A., Hannaway, D.B., West, C.P., Eds.; ASA, CSSA, SSSA: Madison, WI, USA, 2009. [Google Scholar]
- Spiers, D.E.; Eichen, P.A.; Scharf, B.; Settivari, R.S.; Vellios, H.; Johnson, J.; Bryant, J.; Kishore, D. Fescue Toxicosis and Heat Stress: Recent Advances. In Epichloae, Endophytes of Cool Season Grasses: Implications, Utilization and Biology; Young, C.A., Aiken, G.E., McCulley, R.L., Strickland, J.R., Schardl, C.L., Eds.; Noble Foundation: Ardmore, OK, USA, 2012; pp. 20–23. [Google Scholar]
- Al-Haidary, A.; Spiers, D.E.; Rottinghaus, G.E.; Garner, G.B.; Ellersieck, M.R. Thermoregulatory ability of beef heifers following intake of endophyte-infected tall fescue during controlled heat challenge. J. Anim. Sci. 2001, 79, 1780–1788. [Google Scholar] [PubMed]
- Aldrich, C.G.; Paterson, J.A.; Tate, J.L.; Kerley, M.S. The effects of endophyte-infected tall fescue consumption on diet utilization and thermal regulation in cattle. J. Anim. Sci. 1993, 71, 164–170. [Google Scholar] [PubMed]
- Rhodes, M.T.; Paterson, J.A.; Kerley, M.S.; Garner, H.E.; Laughlin, M.H. Reduced blood flow to peripheral and core body tissues in sheep and cattle induced by endophyte-infected tall fescue. J. Anim. Sci. 1991, 69, 2033–2043. [Google Scholar] [PubMed]
- Eisemann, J.H.; Huntington, G.B.; Williamson, M.; Hanna, M.; Poore, M. Physiological responses to known intake of ergot alkaloids by steers at environmental temperatures within or greater than their thermoneutral zone. Front. Chem. 2014, 2, 96. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.A., Jr.; Hemken, R.W.; Boling, J.A.; Harmon, R.J.; Buckner, R.C.; Bush, L.P. Summer fescue toxicity in dairy steers fed tall fescue seed. J. Anim. Sci. 1984, 58, 1057–1061. [Google Scholar] [PubMed]
- Matthews, A.K.; Poore, M.H.; Huntington, G.B.; Green, J.T. Intake, digestion, and N metabolism in steers fed endophyte-free, ergot alkaloid-producing endophyte-infected, or nonergot alkaloid-producing endophyte-infected fescue hay. J. Anim. Sci. 2005, 83, 1179–1185. [Google Scholar] [PubMed]
- Burns, J.C. Nutritive Value. In Tall Fescue for the Twenty-First Century; Fribourg, H.A., Hannaway, D.B., West, C.P., Eds.; ASA, CSSA, SSSA: Madison, WI, USA, 2009. [Google Scholar]
- Ferrell, C.L. Energy Metabolism. In The Ruminant Animal Digestive Physiology and Nutrition; Church, D.C., Ed.; Waveland Press, Inc.: Englewood Cliffs, NJ, USA, 1993; pp. 250–268. [Google Scholar]
- Goetsch, A.L.; Jones, A.L.; Stokes, S.R.; Beers, K.W.; Piper, E.L. Intake, digestion, passage rate and serum prolactin in growing dairy steers fed endophyte-infected fescue with noninfected fescue, clover or wheat straw. J. Anim. Sci. 1987, 64, 1759–1768. [Google Scholar] [PubMed]
- Schumann, B.; Lebzien, P.; Ueberschar, K.H.; Spilke, J.; Holtershinken, M.; Danicke, S. Effects of the level of feed intake and ergot contaminated concentrate on ruminal fermentation and on physiological parameters in cows. Mycotoxin Res. 2008, 24, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Koontz, A.F.; Kim, D.H.; McLeod, K.R.; Klotz, J.L.; Harmon, D.L. Effect of fescue toxicosis on whole body energy and nitrogen balance, in situ degradation and ruminal passage rates in Holstein steers. Anim. Prod. Sci. 2014, 55, 988–998. [Google Scholar] [CrossRef]
- Westendorf, M.L.; Mitchell, G.E., Jr.; Tucker, R.E.; Bush, L.P.; Petroski, R.J.; Powell, R.G. In vitro and in vivo ruminal and physiological responses to endophyte-infected tall fescue. J. Dairy Sci. 1993, 76, 555–563. [Google Scholar] [CrossRef]
- Fahey, G.C.; Berger, L.L. Carbohydrate Nutrition in Ruminants. In The Ruminant Animal Digestive Physiology and Nutrition; Church, D.C., Ed.; Waveland Press Inc.: Englewood Cliffs, NJ, USA, 1993; pp. 269–297. [Google Scholar]
- Stamm, M.M.; DelCurto, T.; Horney, M.R.; Brandyberry, S.D.; Barton, R.K. Influence of alkaloid concentration of tall fescue straw on the nutrition, physiology, and subsequent performance of beef steers. J. Anim. Sci. 1994, 72, 1068–1075. [Google Scholar] [PubMed]
- Koontz, A.F.; Kim, D.H.; Foote, A.P.; Bush, L.P.; Klotz, J.L.; McLeod, K.R.; Harmon, D.L. Alteration of fasting heat production during fescue toxicosis in Holstein steers. J. Anim. Sci. 2013, 91, 3881–3888. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, C.G.; Rhodes, M.T.; Miner, J.L.; Kerley, M.S.; Paterson, J.A. The effects of endophyte-infected tall fescue consumption and use of a dopamine antagonist on intake, digestibility, body-temperature, and blood-constituents in sheep. J. Anim. Sci. 1993, 71, 158–163. [Google Scholar] [PubMed]
- Emile, J.C.; Bony, S.; Ghesquiere, M. Influence of consumption of endophyte-infested tall fescue hay on performance of heifers and lambs. J. Anim. Sci. 2000, 78, 358–364. [Google Scholar] [PubMed]
- Foote, A.P.; Kristensen, N.B.; Klotz, J.L.; Kim, D.H.; Koontz, A.F.; McLeod, K.R.; Bush, L.P.; Schrick, F.N.; Harmon, D.L. Ergot alkaloids from endophyte-infected tall fescue decrease reticuloruminal epithelial blood flow and volatile fatty acid absorption from the washed reticulorumen. J. Anim. Sci. 2013, 91, 5366–5378. [Google Scholar] [CrossRef] [PubMed]
- Hannah, S.M.; Paterson, J.A.; Williams, J.E.; Kerley, M.S.; Miner, J.L. Effects of increasing dietary levels of endophyte-infected tall fescue seed on diet digestibility and ruminal kinetics in sheep. J. Anim. Sci. 1990, 68, 1693–1701. [Google Scholar] [PubMed]
- Dalziel, J.E.; Dunstan, K.E.; Finch, S.C. Combined effects of fungal alkaloids on intestinal motility in an in vitro rat model. J. Anim. Sci. 2013, 91, 5177–5182. [Google Scholar] [CrossRef] [PubMed]
- McLeay, L.M.; Smith, B.L. Effects of ergotamine and ergovaline on the electromyographic activity of smooth muscle of the reticulum and rumen of sheep. Am. J. Vet. Res. 2006, 67, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.P.; Littler, R.A.; Smith, B.L.; McLeay, L.M. Effects and mechanisms of action of the ergopeptides ergotamine and ergovaline and the effects of peramine on reticulum motility of sheep. Am. J. Vet. Res. 2009, 70, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Klotz, J.L.; Aiken, G.E.; Johnson, J.M.; Brown, K.R.; Bush, L.P.; Strickland, J.R. Antagonism of lateral saphenous vein serotonin receptors from steers grazing endophyte-free, wild-type, or novel endophyte-infected tall fescue. J. Anim. Sci. 2013, 91, 4492–4500. [Google Scholar] [CrossRef] [PubMed]
- Briejer, M.R.; Mathis, C.; Schuurkes, J.A. 5-HT receptor types in the rat ileum longitudinal muscle: Focus on 5-HT2 receptors mediating contraction. Neurogastroenterol. Motil. 1997, 9, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Galligan, J.J. 5-HT1A and 5-HT4 receptors mediate inhibition and facilitation of fast synaptic transmission in enteric neurons. Am. J. Physiol. 1994, 266, G230–G238. [Google Scholar] [PubMed]
- Talley, N.J. Review article: 5-hydroxytryptamine agonists and antagonists in the modulation of gastrointestinal motility and sensation: Clinical implications. Aliment. Pharmacol. Ther. 1992, 6, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Egert, A.M.; Klotz, J.L.; McLeod, K.R.; Harmon, D.L. Development of a methodology to measure the effect of ergot alkaloids on forestomach motility using real-time wireless telemetry. Front. Chem. 2014, 2, 90. [Google Scholar] [CrossRef] [PubMed]
- Klotz, J.L.; Kim, D.H.; Foote, A.P.; Harmon, D.L. Effects of ergot alkaloid exposure on serotonin receptor mRNA in the smooth muscle of the bovine gastrointestinal tract. J. Anim. Sci. 2014, 92, 890–891. [Google Scholar]
- Porter, J.K.; Thompson, F.N., Jr. Effects of fescue toxicosis on reproduction in livestock. J. Anim. Sci. 1992, 70, 1594–1603. [Google Scholar] [PubMed]
- Hurley, W.L.; Convey, E.M.; Leung, K.; Edgerton, L.A.; Hemken, R.W. Bovine prolactin, TSH, T4 and T3 concentrations as affected by tall fescue summer toxicosis and temperature. J. Anim. Sci. 1980, 51, 374–379. [Google Scholar] [PubMed]
- Gooneratne, S.R.; Scannell, M.; Wellby, M.; Fletcher, L. Changes in concentrations of lysergol in urine and prolactin in plasma, rectal temperature and respiration rate in sheep selected for resistance or susceptibility to ryegrass staggers and fed ergovaline. N. Z. Vet. J. 2011, 59, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Monroe, J.L.; Cross, D.L.; Hudson, L.W.; Hendricks, D.M.; Kennedy, S.W.; Bridges, W.C., Jr. Effect of selenium and endophyte-contaminated fescue on performance and reproduction in mares. J. Equine Vet. Sci. 1988, 8, 148–153. [Google Scholar] [CrossRef]
- Schillo, K.K.; Leshin, L.S.; Boling, J.A.; Gay, N. Effects of endophyte-infected fescue on concentrations of prolactin in blood sera and the anterior pituitary and concentrations of dopamine and dopamine metabolites in brains of steers. J. Anim. Sci. 1988, 66, 713–718. [Google Scholar] [PubMed]
- Houdebine, L.M.; Djiane, J.; Dusanter-Fourt, I.; Martel, P.; Kelly, P.A.; Devinoy, E.; Servely, J.L. Hormonal action controlling mammary activity. J. Dairy Sci. 1985, 68, 489–500. [Google Scholar] [CrossRef]
- Strahan, S.R.; Hemken, R.W.; Jackson, J.A., Jr.; Buckner, R.C.; Bush, L.P.; Siegel, M.R. Performance of lactating dairy cows fed tall fescue forage. J. Dairy Sci. 1987, 70, 1228–1234. [Google Scholar] [CrossRef]
- Stilham, W.D.; Brown, C.J.; Daniels, L.B.; Piper, E.L.; Fetherstone, H.E. Toxic fescue linked to reduced milk output in ewes. Arkansas Farm. Res. 1992, 31, 9. [Google Scholar]
- McClanahan, L.K.; Aiken, G.E.; Dougherty, C.T. Case Study: Influence of Rough Hair Coats and Steroid Implants on the Performance and Physiology of Steers Grazing Endophyte-Infected Tall Fescue in the Summer. Prof. Anim. Sci. 2008, 24, 269–276. [Google Scholar]
- Thompson, D.L., Jr.; Hoffman, R.; DePew, C.L. Prolactin administration to seasonally anestrous mares: Reproductive, metabolic, and hair-shedding responses. J. Anim. Sci. 1997, 75, 1092–1099. [Google Scholar] [PubMed]
- Aiken, G.E.; Klotz, J.L.; Looper, M.L.; Tabler, S.F.; Schrick, F.N. Disrupted hair follicle activity in cattle grazing endophyte-infected tall fescue in the summer insulates core body temperatures. Prof. Anim. Sci. 2011, 27, 336–343. [Google Scholar]
- Jones, K.L.; King, S.S.; Griswold, K.E.; Cazac, D.; Cross, D.L. Domperidone can ameliorate deleterious reproductive effects and reduced weight gain associated with fescue toxicosis in heifers. J. Anim. Sci. 2003, 81, 2568–2574. [Google Scholar] [PubMed]
- Seals, R.C.; Schuenemann, G.M.; Lemaster, J.W.; Saxton, A.M.; Waller, J.C.; Schrick, F.N. Follicular dynamics in beef heifers consuming ergotamine tartrate as a model of endophyte-infected tall fescue consumption. J. Anim. Vet. Adv. 2005, 4, 97–102. [Google Scholar]
- Burke, J.M.; Rorie, R.W. Changes in ovarian function in mature beef cows grazing endophyte infected tall fescue. Theriogenology 2002, 57, 1733–1742. [Google Scholar] [CrossRef]
- Burke, J.M.; Rorie, R.W.; Piper, E.L.; Jackson, W.G. Reproductive responses to grazing endophyte-infected tall fescue by postpartum beef cows. Theriogenology 2001, 56, 357–369. [Google Scholar] [CrossRef]
- Burke, J.M.; Jackson, W.G.; Robson, G.A. Seasonal changes in body weight and condition, and pregnancy and lambing rates of sheep on endophyte-infected tall fescue in the south-eastern United States. Small Rumin. Res. 2002, 44, 141–151. [Google Scholar] [CrossRef]
- Schmidt, S.P.; Danilson, D.A.; Holliman, J.A.; Grimes, H.W.; Webster, W.B. Fescue Fungus Supresses Growth and Reproduction in Replacement Beef Heifers. Highlights Agric. Res., Alabama Agric. Exp. Station 1986, 33, 15. [Google Scholar]
- Schuenemann, G.M.; Hockett, M.E.; Edwards, J.L.; Rohrbach, N.R.; Breuel, K.F.; Schrick, F.N. Embryo development and survival in beef cattle administered ergotamine tartrate to simulate fescue toxicosis. Reprod. Biol. 2005, 5, 137–150. [Google Scholar] [PubMed]
- Oliver, J.W.; Schultze, A.E.; Rohrbach, B.W.; Fribourg, H.A.; Ingle, T.; Waller, J.C. Alterations in hemograms and serum biochemical analytes of steers after prolonged consumption of endophyte-infected tall fescue. J. Anim. Sci. 2000, 78, 1029–1035. [Google Scholar] [PubMed]
- Strickland, J.R.; Cross, D.L.; Birrenkott, G.P.; Grimes, L.W. Effect of ergovaline, loline, and dopamine antagonists on rat pituitary cell prolactin release in vitro. Am. J. Vet. Res. 1994, 55, 716–721. [Google Scholar] [PubMed]
- Louw, B.P.; Lishman, A.W.; Botha, W.A.; Baumgartner, J.P. Failure to demonstrate a role for the acute release of prolactin at oestrus in the ewe. J. Reprod. Fertil. 1974, 40, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Browning, R., Jr.; Schrick, F.N.; Thompson, F.N.; Wakefield, T., Jr. Reproductive hormonal responses to ergotamine and ergonovine in cows during the luteal phase of the estrous cycle. J. Anim. Sci. 1998, 76, 1448–1454. [Google Scholar] [PubMed]
- Browning, R., Jr.; Thompson, F.N.; Sartin, J.L.; Leite-Browning, M.L. Plasma concentrations of prolactin, growth hormone, and luteinizing hormone in steers administered ergotamine or ergonovine. J. Anim. Sci. 1997, 75, 796–802. [Google Scholar] [PubMed]
- Hodson, D.J.; Henderson, H.L.; Townsend, J.; Tortonese, D.J. Photoperiodic modulation of the suppressive actions of prolactin and dopamine on the pituitary gonadotropin responses to gonadotropin-releasing hormone in sheep. Biol. Reprod. 2012, 86, 122. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.W. Physiological Manifestations of Endophyte Toxicosis in Ruminant and Laboratory Species. In Neotyphodium/Grass Interactions; Springer: New York, NY, 1997; pp. 311–346. [Google Scholar]
- Duckett, S.K.; Andrae, J.G.; Pratt, S.L. Exposure to ergot alkaloids during gestation reduces fetal growth in sheep. Front. Chem. 2014, 2, 68. [Google Scholar] [CrossRef] [PubMed]
- Watson, R.H.; McCann, M.A.; Parish, J.A.; Hoveland, C.S.; Thompson, F.N.; Bouton, J.H. Productivity of cow-calf pairs grazing tall fescue pastures infected with either the wild-type endophyte or a nonergot alkaloid-producing endophyte strain, AR542. J. Anim. Sci. 2004, 82, 3388–3393. [Google Scholar] [PubMed]
- Robinson, D.L.; Cafe, L.M.; Greenwood, P.L. Meat Science and Muscle Biology Symposium: Developmental programming in cattle: Consequences for growth, efficiency, carcass, muscle, and beef quality characteristics. J. Anim. Sci. 2013, 91, 1428–1442. [Google Scholar] [CrossRef] [PubMed]
- Cross, D.L. Toxic Effects of the Endophyte in Horses. In Tall Fescue for the Twenty-First Century; Fribourg, H.A., Hannaway, D.B., West, C.P., Eds.; ASA, CSSA, SSSA: Madison, WI, USA, 2009. [Google Scholar]
- Putnam, M.R.; Bransby, D.I.; Schumacher, J.; Boosinger, T.R.; Bush, L.; Shelby, R.A.; Vaughan, J.T.; Ball, D.; Brendemuehl, J.P. Effects of the fungal endophyte Acremonium coenophialum in fescue on pregnant mares and foal viability. Am. J. Vet. Res. 1991, 52, 2071–2074. [Google Scholar] [PubMed]
- Forsyth, I.A. Variation among species in the endocrine control of mammary growth and function: The roles of prolactin, growth hormone, and placental lactogen. J. Dairy Sci. 1986, 69, 886–903. [Google Scholar] [CrossRef]
- Schuenemann, G.M.; Edwards, J.L.; Davis, M.D.; Blackmon, H.E.; Scenna, F.N.; Rohrbach, N.R.; Saxton, A.M.; Adair, H.S.; Hopkins, F.M.; Waller, J.C.; et al. Effects of administration of ergotamine tartrate on fertility of yearling beef bulls. Theriogenology 2005, 63, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Schuenemann, G.M.; Edwards, J.L.; Hopkins, F.M.; Rohrbach, N.R.; Adair, H.S.; Scenna, F.N.; Waller, J.C.; Oliver, J.W.; Saxton, A.M.; Schrick, F.N. Fertility aspects in yearling beef bulls grazing endophyte-infected tall fescue pastures. Reprod. Fertil. Dev. 2005, 17, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Pratt, S.L.; Stowe, H.M.; Whitlock, B.K.; Strickland, L.; Miller, M.; Calcatera, S.M.; Dimmick, M.D.; Aiken, G.E.; Schrick, F.N.; Long, N.M.; et al. Bulls grazing Kentucky 31 tall fescue exhibit impaired growth, semen quality, and decreased semen freezing potential. Theriogenology 2015, 83, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Looper, M.L.; Johnson, Z.B.; Rorie, R.W.; Rosenkrans, C.F., Jr. Involvement of signaling pathways in bovine sperm motility, and effect of ergot alkaloids. Vitro Cell Dev. Biol. Anim. 2009, 45, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Looper, M.L.; Rorie, R.W.; Person, C.N.; Lester, T.D.; Hallford, D.M.; Aiken, G.E.; Roberts, C.A.; Rottinghaus, G.E.; Rosenkrans, C.F., Jr. Influence of toxic endophyte-infected fescue on sperm characteristics and endocrine factors of yearling Brahman-influenced bulls. J. Anim. Sci. 2009, 87, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Pratt, S.L.; Calcatera, S.M.; Stowe, H.M.; Dimmick, M.A.; Schrick, F.N.; Duckett, S.K.; Andrae, J.G. Identification of bovine prolactin in seminal fluid, and expression and localization of the prolactin receptor and prolactin-inducible protein in the testis and epididymis of bulls exposed to ergot alkaloids. Theriogenology 2015, 83, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Stowe, H.M.; Calcatera, S.M.; Dimmick, M.A.; Andrae, J.G.; Duckett, S.K.; Pratt, S.L. The bull sperm microRNAome and the effect of fescue toxicosis on sperm microRNA expression. PLoS ONE 2014, 9, e113163. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.J.; Tyler, D.E.; Papp, E. Abdominal fat necrosis as a herd problem in Georgia cattle. J. Am. Vet. Med. Assoc. 1969, 154, 1017–1021. [Google Scholar] [PubMed]
- Smith, G.W.; Rotstein, D.S.; Brownie, C.F. Abdominal fat necrosis in a pygmy goat associated with fescue toxicosis. J. Vet. Diagn. Investig. 2004, 16, 356–359. [Google Scholar] [CrossRef]
- Wolfe, B.A.; Bush, M.; Monfort, S.L.; Mumford, S.L.; Pessier, A.; Montali, R.J. Abdominal lipomatosis attributed to tall fescue toxicosis in deer. J. Am. Vet. Med. Assoc. 1998, 213, 1754, 1783–1786. [Google Scholar]
- Vitovec, J.; Proks, C.; Valvoda, V. Lipomatosis (fat necrosis) in cattle and pigs. J. Comp. Pathol. 1975, 85, 53–59. [Google Scholar] [CrossRef]
- White, S.L. Fat necrosis in horses. J. Equine Vet. Sci. 1993, 13, 615–616. [Google Scholar] [CrossRef]
- Stuedemann, J.A.; Rumsey, T.S.; Bond, J.; Wilkinson, S.R.; Bush, L.P.; Williams, D.J.; Caudle, A.B. Association of blood cholesterol with occurrence of fat necrosis in cows and tall fescue summer toxicosis in steers. Am. J. Vet. Res. 1985, 46, 1990–1995. [Google Scholar] [PubMed]
- Realini, C.E.; Duckett, S.K.; Hill, N.S.; Hoveland, C.S.; Lyon, B.G.; Sackmann, J.R.; Gillis, M.H. Effect of endophyte type on carcass traits, meat quality, and fatty acid composition of beef cattle grazing tall fescue. J. Anim. Sci. 2005, 83, 430–439. [Google Scholar] [PubMed]
- Filipov, N.M.; Thompson, F.N.; Stuedemann, J.A.; Elsasser, T.H.; Kahl, S.; Sharma, R.P.; Young, C.R.; Stanker, L.H.; Smith, C.K. Increased responsiveness to intravenous lipopolysaccharide challenge in steers grazing endophyte-infected tall fescue compared with steers grazing endophyte-free tall fescue. J. Endocrinol. 1999, 163, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Greatorex, J.C.; Mantle, P.G. Effect or rye ergot on the pregnant sheep. J. Reprod. Fertil. 1974, 37, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Foote, A.; Penner, G.; Walpole, M.; Klotz, J.; Brown, K.; Bush, L.; Harmon, D. Acute exposure to ergot alkaloids from endophyte-infected tall fescue does not alter absorptive or barrier function of the isolated ruminal epithelium. Animal 2014, 8, 1106–1112. [Google Scholar] [CrossRef]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: meta-analysis of published experiments in animals. Toxins (Basel) 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klotz, J.L. Activities and Effects of Ergot Alkaloids on Livestock Physiology and Production. Toxins 2015, 7, 2801-2821. https://doi.org/10.3390/toxins7082801
Klotz JL. Activities and Effects of Ergot Alkaloids on Livestock Physiology and Production. Toxins. 2015; 7(8):2801-2821. https://doi.org/10.3390/toxins7082801
Chicago/Turabian StyleKlotz, James L. 2015. "Activities and Effects of Ergot Alkaloids on Livestock Physiology and Production" Toxins 7, no. 8: 2801-2821. https://doi.org/10.3390/toxins7082801
APA StyleKlotz, J. L. (2015). Activities and Effects of Ergot Alkaloids on Livestock Physiology and Production. Toxins, 7(8), 2801-2821. https://doi.org/10.3390/toxins7082801