
Effects of Citric and Lactic Acid on the Reduction of Deoxynivalenol and Its Derivatives in Feeds


Abstract
:1. Introduction
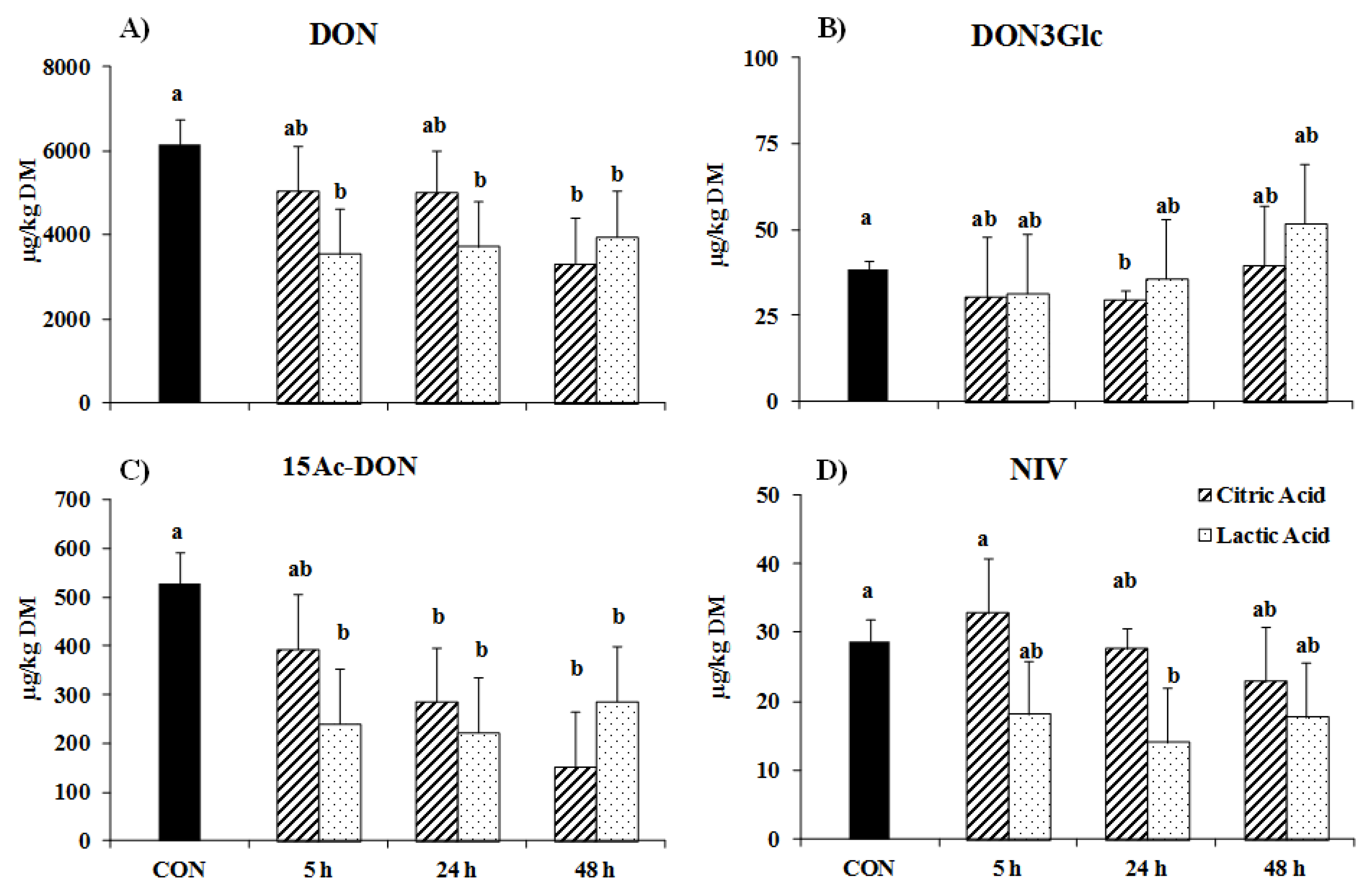
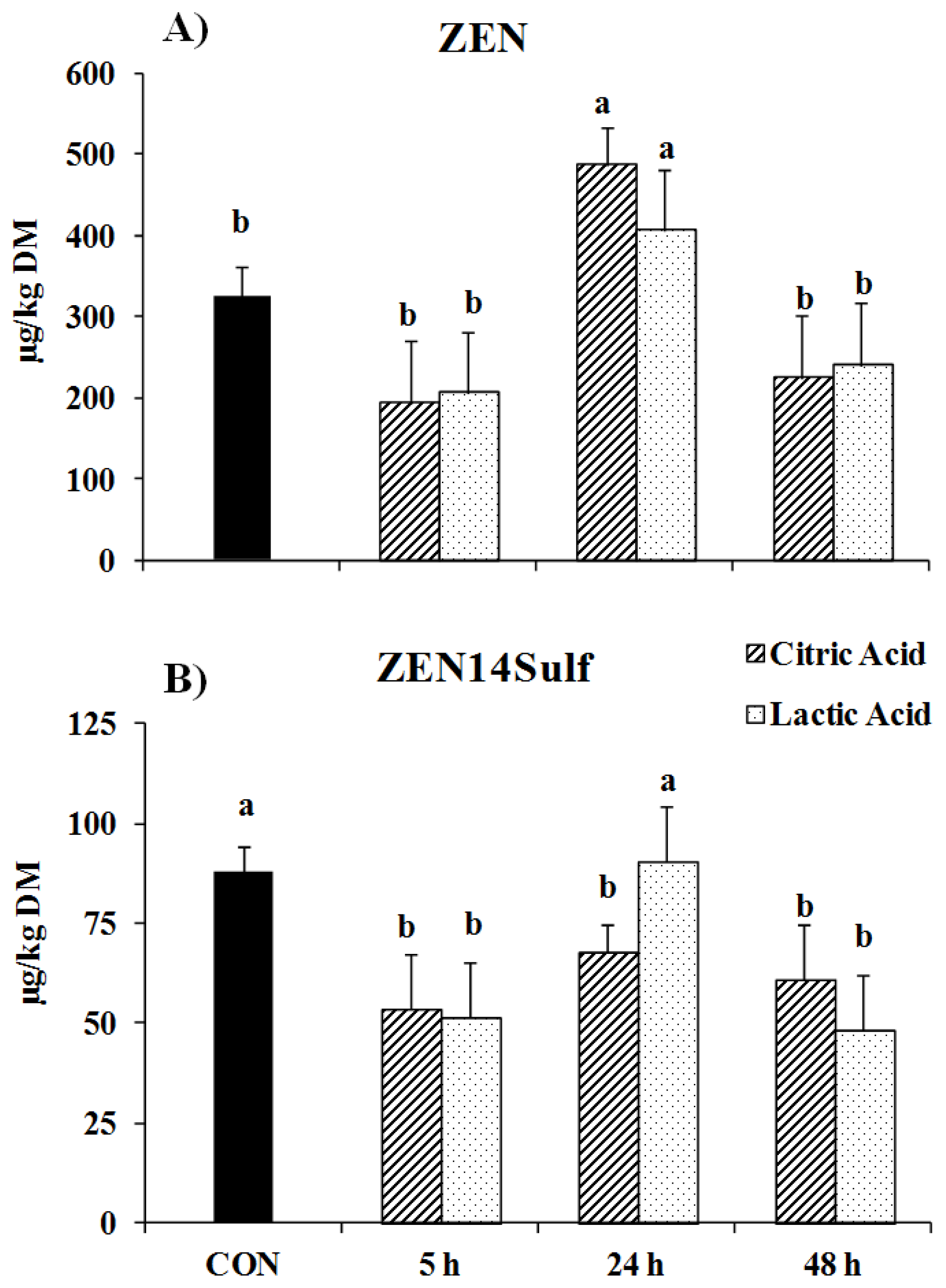
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Processing Procedure of Feed Samples with Organic Acids
5.2. Multi-Mycotoxin LC-MS/MS Analysis
5.3. Statistical Analysis
Author Contributions
Conflicts of Interest
References
- Ghareeb, K.; Awad, W.A.; Böhm, J.; Zebeli, Q. Impacts of the feed contaminant deoxynivalenol on the intestine of monogastric animals: Poultry and swine. J. Appl. Toxicol. 2015, 35, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar]
- Kabak, B.; Dobson, A.; Var, I. Strategies to prevent mycotoxin contamination of food and animal feed: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 593–619. [Google Scholar] [CrossRef] [PubMed]
- Escrivá, L.; Font, G.; Manyes, L. In vivo toxicity studies of Fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Boudergue, C.; Burel, C.; Dragacci, S.; Favrot, M.C.; Fremy, J.M.; Massimi, C.; Pringent, P.; Debongnie, P.; Pussemier, L.; Boudra, H.; et al. Review of Mycotoxin-Detoxifying Agents Used as Feed Additives: Mode of Action, Efficacy and Feed/Food Safety; CFP/EFSA/FEEDAP/2009/01; EFSA: Parma, Italy, 2009. [Google Scholar]
- Bennett, J.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarch, A.; Lebrihi, A. Review of mycotoxin reduction in food and feed: From prevention in the field to detoxification by adsorption or transformation. Food Addit. Contam. 2011, 28, 1590–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarch, A.; Andre, F.; Delaforge, M.; Lebrihi, A. Transformation of zearalenone to zearalenone-sulfate by Aspergillus spp. World Mycotoxin J. 2010, 3, 183–191. [Google Scholar] [CrossRef]
- Mendez-Albores, A.; Del Rio-Garcia, J.; Moreno-Martinez, E. Decontamination of aflatoxin duckling feed with aqueous citric acid treatment. Anim. Feed Sci. Technol. 2007, 135, 249–262. [Google Scholar] [CrossRef]
- Khol-Parisini, A.; Humer, E.; Sizmaz, Ö.; Abdel-Raheem, S.; Gruber, L.; Gasteiner, J.; Zebeli, Q. Ruminal disappearance of phosphorus and starch, reticuloruminal pH and total tract nutrient digestibility in dairy cows fed diets differing in grain processing. Anim. Feed Sci. Technol. 2015, 210, 74–85. [Google Scholar] [CrossRef]
- Metzler-Zebeli, B.U.; Deckardt, K.; Schollenberger, M.; Rodehutscord, M.; Zebeli, Q. Lactic acid and thermal treatments trigger the hydrolysis of Myo-Inositol Phosphates and modify the abundance of lower Myo-Inositol Phosphates in barley (Hordeum vulgare L.). PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Albores, A.; Martinez-Bustos, F.; Gaytan-Martinez, M.; Moreno-Martinez, E. Effect of lactic and citric acid on the stability of B-aflatoxins in extrusion-cooked sorghum. Lett. Appl. Microbiol. 2008, 47, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nelson, P.; Dignani, M.; Anaissie, E. Taxonomy, biology and clinical aspects of Fusarium species. Clin. Microbiol. Rev. 1994, 7, 479–504. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, G.; Pettersson, H.; Lundh, T. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Seeboth, J.; Solinhac, R.; Oswald, I.; Guzylack-Piriou, L. The fungal T-2 toxin alters the activation of primary macrophages induced by TLR-agonists resulting in a decrease of the inflammatory response in the pig. Vet. Res. 2012, 43. [Google Scholar] [CrossRef] [PubMed]
- Summerell, B.; Leslie, J. Fifty years of Fusarium: How could nine species have ever been enough? Fungal Divers. 2011, 50, 135–144. [Google Scholar] [CrossRef]
- Berthiller, F.; Krska, R.; Domig, K.J.; Kneifel, W.; Juge, N.; Schuhmacher, R.; Adam, G. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 2011, 206, 264–267. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Evaluation of Certain Contaminants in Food. Seventy-Second Report of the Joint FAO/WHO Expert Comitee on Food Additives; Technical Report Series 959; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Juan, C.; Ritieni, A.; Manes, J. Determination of trichothecenes and zearalenones in grain cereal, flour and bread by liquid chromatography tandem mass spectrometry. Food Chem. 2012, 134, 2389–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority (ESFA). Scientific Opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9. [Google Scholar] [CrossRef]
- Plasencia, J.; Mirocha, C. Isolation and characterization of zearalenone sulfate produced by Fusarium spp. Appl. Environ. Microbiol. 1991, 57, 146–150. [Google Scholar] [PubMed]
- Scarpino, V.; Reyneri, A.; Sulyok, M.; Krska, R.; Blandino, M. Effect of fungicide application to control Fusarium head blight and 20 Fusarium and Alternaria mycotoxins in winter wheat (Triticum aestivum L.). World Mycotoxin J. 2015, 8, 499–510. [Google Scholar] [CrossRef]
- Pedersen, P.B.; Miller, J.D. The fungal metabolite culmorin and related compounds. Nat. Toxins 1999, 7, 305–309. [Google Scholar] [CrossRef]
- Ghebremeskel, M.; Langseth, W. The occurrence of culmorin and hydroxy-culmorins in cereals. Mycopathologia 2001, 152, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Zamir, L.; Nikolakakis, A.; Huang, L.; St-Pierre, P.; Sauriol, F.; Sparace, S.; Mamer, O. Biosynthesis of 3-acetyldeoxynivalenol and sambucinol—Identification of the two oxygenation steps after trichodiene. J. Biol. Chem. 1999, 274, 12269–12277. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, B.A.; Seguin, C.; Overy, D. Trichothecenes and other secondary metabolites from Fusarium graminearum—is it just about deoxynivalenol? Can. J. Plant Pathol. 2012, 34, 325–326. [Google Scholar]
- Rotter, R.G.; Thompson, B.K.; Trenholm, H.L.; Prelusky, D.B.; Hartin, K.E.; Miller, J.D. A preliminary examinatino of potential interactions between deoxynivalenol (DON) and other selected Fusarium mycotoxins in growing pigs. Can. J. Anim. Sci. 1992, 72, 107–116. [Google Scholar] [CrossRef]
- Dowd, P.; Miller, J.; Greenhalgh, R. Toxicity and interactions of some Fusarium graminearum metabolites to caterpillars. Mycologia 1989, 81, 646–650. [Google Scholar] [CrossRef]
- Gutleb, A.; Morrison, E.; Murk, A. Cytotoxicity assays for mycotoxins produced by Fusarium strains: A review. Environ. Toxicol. Pharmacol. 2002, 11, 309–320. [Google Scholar] [CrossRef]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, V.; Grothe, T.; Mayer-Bartschmid, A.; Endermann, R.; Geschke, F.U.; Henkel, T.; Stadler, M. Altersetin, a new antibiotic from cultures of endophytic Alternaria spp. taxonomy, fermentation, isolation, structure elucidation and biological activities. J. Antibiot. 2002, 55, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Jalili, M.; Jinap, S.; Son, R. The effect of chemical treatment on reduction of aflatoxins and ochratoxin A in black and white pepper during washing. Food Addit. Contam. 2011, 28, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Albores, A.; Arambula-Villa, G.; Loarea-Pina, M.; Castano-Tostado, E.; Moreno-Martinez, E. Safety and efficacy evaluation of aqueous citric acid to degrade B-aflatoxins in maize. Food Chem. Toxicol. 2005, 43, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, K.J.; Steyn, P.S.; Fourie, L. Mycotoxins. II. The constitution of ochratoxins A, B, and C, metabolites of Aspergillus ochraceus wilh. J. Chem. Soc. 1965, 7083–7088. [Google Scholar] [CrossRef]
- Harder, H.; Khol-Parisini, A.; Zebeli, Q. Treatments with organic acids and pullulanase differently affect resistant starch and fiber composition in flour of various barley genotypes (Hordeum vulgare L.). Starch-Stärke 2014, 67, 512–520. [Google Scholar] [CrossRef]
- Harder, H.; Khol-Parisini, A.; Zebeli, Q. Modulation of resistant starch and nutrient composition of barley grain using organic acids and thermal cycling treatments. Starch-Stärke 2015, 67, 654–662. [Google Scholar] [CrossRef]
- Harder, H.; Khol-Parisini, A.; Metzler-Zebeli, B.U.; Klevenhusen, F.; Zebeli, Q. Treatment of grain with organic acids at 2 different dietary phosphours levels modulates ruminal microbial communitiy structure and fermentation patterns in vitro. J. Dairy Sci. 2015, 98, 8107–8120. [Google Scholar] [CrossRef] [PubMed]
- Diaz, G.; Krska, R.; Sulyok, M. Mycotoxins and cyanogenic glycosides in staple foods of three indigenous people of the Colombian Amazon. Food Addit. Contam. 2015, 8, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef] [PubMed]
- Debongnie, P.; Tangni, E.; Callebaut, A. Report on the 2014 Proficiency Test for the Determination of 19 Mycotoxins and 4 Sums of Mycotoxins in Rye Flour; CODA-CERVA: Tervuren, Belgium, 2015. [Google Scholar]
- De Girolamo, A.; Ciasca, B.; Stroka, J.; Bratinova, S.; Visconti, A.; Lattanzio, V.M.T. Report of the 2014 Proficiency Test for LC-MS(MS) Multi-Mycotoxin Methods, Determination of DON, FB1, FB2, ZEA, T-2, HT-2, OTA, AFB1, AFG1, AFB2, AFG2 in Maize and Determination of DON, ZEA, T-2, HT-2, OTA in Wheat; Institute of Sciences of Food Production, National Research Council of Italy: Bari, Italy, 2015. [Google Scholar]



{kind=link}
{kind=link}
| Variable | Treatment | p-Value (Contrast) | |||||
|---|---|---|---|---|---|---|---|
| CON | CA | LA | SEM | CON vs. LA | CON vs.CA | CON vs. LA + CA | |
| T2 | 1.4 | n.d. | n.d. | 0.29 | - | - | - |
| FB1 | 35.6 b | 58.0 a | 62.4 a | 5.08 | <0.01 | <0.01 | <0.01 |
| FB2 | 19.2 b | 25.2 a | 21.1 b | 1.21 | <0.01 | 0.34 | 0.02 |
| Culmorin | 127.5 a | 116.5 ab | 103.7 b | 10.09 | 0.14 | 0.30 | 0.08 |
| 5-Hydroxyculmorin | 740.4 a | 684.5 ab | 474.4 b | 20.83 | <0.01 | 0.36 | 0.03 |
| 15-Hydroxyculmorin | 314.4 a | 278.9 a | 259.0 b | 9.20 | 0.07 | 0.21 | 0.09 |
| Sambucinol | 179.0 a | 105.0 b | 92.9 b | 16.91 | <0.01 | <0.01 | <0.01 |
| Decalonectrin | 87.0 a | 69.4 ab | 60.1 b | 10.09 | 0.22 | 0.07 | 0.08 |
| Enniatin A | 1.0 | 0.9 | 0.8 | 0.08 | 0.82 | 0.20 | 0.38 |
| Enniatin A1 | 11.1 | 10.4 | 10.1 | 0.89 | 0.44 | 0.59 | 0.42 |
| Enniatin B | 40.1 | 34.1 | 50.7 | 4.61 | 0.68 | 0.37 | 0.12 |
| Enniatin B1 | 23.5 | 19.5 | 20.5 | 2.21 | 0.24 | 0.37 | 0.24 |
| Enniatin B2 | 1.2 a | 0.8 b | 1.2 a | 0.11 | 0.01 | 0.66 | 0.08 |
| Moniliformin | 10.1 | 10.1 | 16.4 | 2.27 | 0.99 | 0.11 | 0.35 |
| Beauvericin | 10.5 a | 8.5 b | 8.7 ab | 0.76 | 0.08 | 0.12 | 0.06 |
| Aurofusarin | 5306 | 5039 | 4562 | 642.5 | 0.77 | 0.42 | 0.53 |
| Antibiotic Y | 71.1 a | 30.4 b | 27.9 a | 6.50 | <0.01 | <0.01 | <0.01 |
| Apicidin | 2.3 a | 1.7 b | 2.2 a | 0.10 | <0.01 | 0.84 | 0.04 |
| Epiequisetin | 0.7 | 0.7 | 0.7 | 0.07 | 0.38 | 0.85 | 0.53 |
| Equisetin | 4.4 | 3.8 | 4.4 | 0.67 | 0.53 | 0.99 | 0.71 |
| Fusaric acid | 46.0 b | 40.1 b | 102.0 a | 24.78 | 0.84 | 0.08 | 0.33 |
| Fusarin C | 810.4 | 876.9 | 510.9 | 200.5 | 0.82 | 0.30 | 0.64 |
| Fusarinolic acid | 16.0 b | 61.1 a | 34.7 b | 10.99 | 0.01 | 0.25 | 0.03 |
| Tenuazonic acid | 65.5 b | 77.6 ab | 81.6 a | 6.06 | 0.17 | 0.08 | 0.07 |
| Alternariol | 17.1 | 15.9 | 15.0 | 0.89 | 0.36 | 0.13 | 0.17 |
| Alternariolmethylether | 7.6 | 6.8 | 8.7 | 0.71 | 0.47 | 0.27 | 0.83 |
| Altersetin | 54.7 a | 39.4 b | 30.8 b | 4.12 | 0.02 | <0.01 | <0.01 |
| Tentoxin | 0.4 b | 0.3 b | 0.9 a | 0.12 | 0.53 | 0.01 | 0.22 |
| Infectopyron | 352.0 a | 291.7 b | 298.3 b | 9.58 | <0.01 | <0.01 | <0.01 |
| Nitropropionic acid | 52.4 a | 28.1 c | 41.2 b | 1.82 | <0.01 | <0.01 | <0.01 |
| Curvularin | 5.7 | 6.7 | 7.4 | 1.33 | 0.65 | 0.44 | 0.49 |
| Asperglaucide | 8.9 a | 5.8 b | 6.3 b | 0.30 | <0.01 | <0.01 | <0.01 |
| Asperphenamate | 2.7 a | 2.0 b | 2.1 b | 0.14 | <0.01 | <0.01 | <0.01 |
| Brevianamid F | 32.2 b | 36.4 a | 26.6 b | 1.40 | 0.12 | 0.07 | 0.72 |
| Cyclo(L-Pro-L-Tyr) | 205.5 a | 139.4 b | 120.1 c | 10.11 | <0.01 | <0.01 | <0.01 |
| Cyclo(L-Pro-L-Val) | 142.1 a | 125.3 b | 112.7 c | 3.39 | <0.01 | <0.01 | <0.01 |
| Emodin | 5.3 | 4.9 | 3.8 | 0.78 | 0.74 | 0.19 | 0.34 |
| Lotaustralin | 44.8 a | 36.5 b | 38.1 ab | 3.04 | 0.12 | 0.02 | 0.04 |
| Neoechinulin A | 9.9 a | 5.2 b | 4.7 b | 0.44 | <0.01 | <0.01 | <0.01 |
| Rugulusovin | 5.2 b | 7.5 a | 8.0 a | 0.60 | <0.01 | 0.01 | <0.01 |
| Tryptophol | 197.8 a | 175.3 b | 188.6 ab | 5.81 | 0.01 | 0.27 | 0.04 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Humer, E.; Lucke, A.; Harder, H.; Metzler-Zebeli, B.U.; Böhm, J.; Zebeli, Q. Effects of Citric and Lactic Acid on the Reduction of Deoxynivalenol and Its Derivatives in Feeds. Toxins 2016, 8, 285. https://doi.org/10.3390/toxins8100285
Humer E, Lucke A, Harder H, Metzler-Zebeli BU, Böhm J, Zebeli Q. Effects of Citric and Lactic Acid on the Reduction of Deoxynivalenol and Its Derivatives in Feeds. Toxins. 2016; 8(10):285. https://doi.org/10.3390/toxins8100285
Chicago/Turabian StyleHumer, Elke, Annegret Lucke, Hauke Harder, Barbara U. Metzler-Zebeli, Josef Böhm, and Qendrim Zebeli. 2016. "Effects of Citric and Lactic Acid on the Reduction of Deoxynivalenol and Its Derivatives in Feeds" Toxins 8, no. 10: 285. https://doi.org/10.3390/toxins8100285
APA StyleHumer, E., Lucke, A., Harder, H., Metzler-Zebeli, B. U., Böhm, J., & Zebeli, Q. (2016). Effects of Citric and Lactic Acid on the Reduction of Deoxynivalenol and Its Derivatives in Feeds. Toxins, 8(10), 285. https://doi.org/10.3390/toxins8100285