
A Topographical Atlas of Shiga Toxin 2e Receptor Distribution in the Tissues of Weaned Piglets

Abstract
:1. Introduction
2. Results
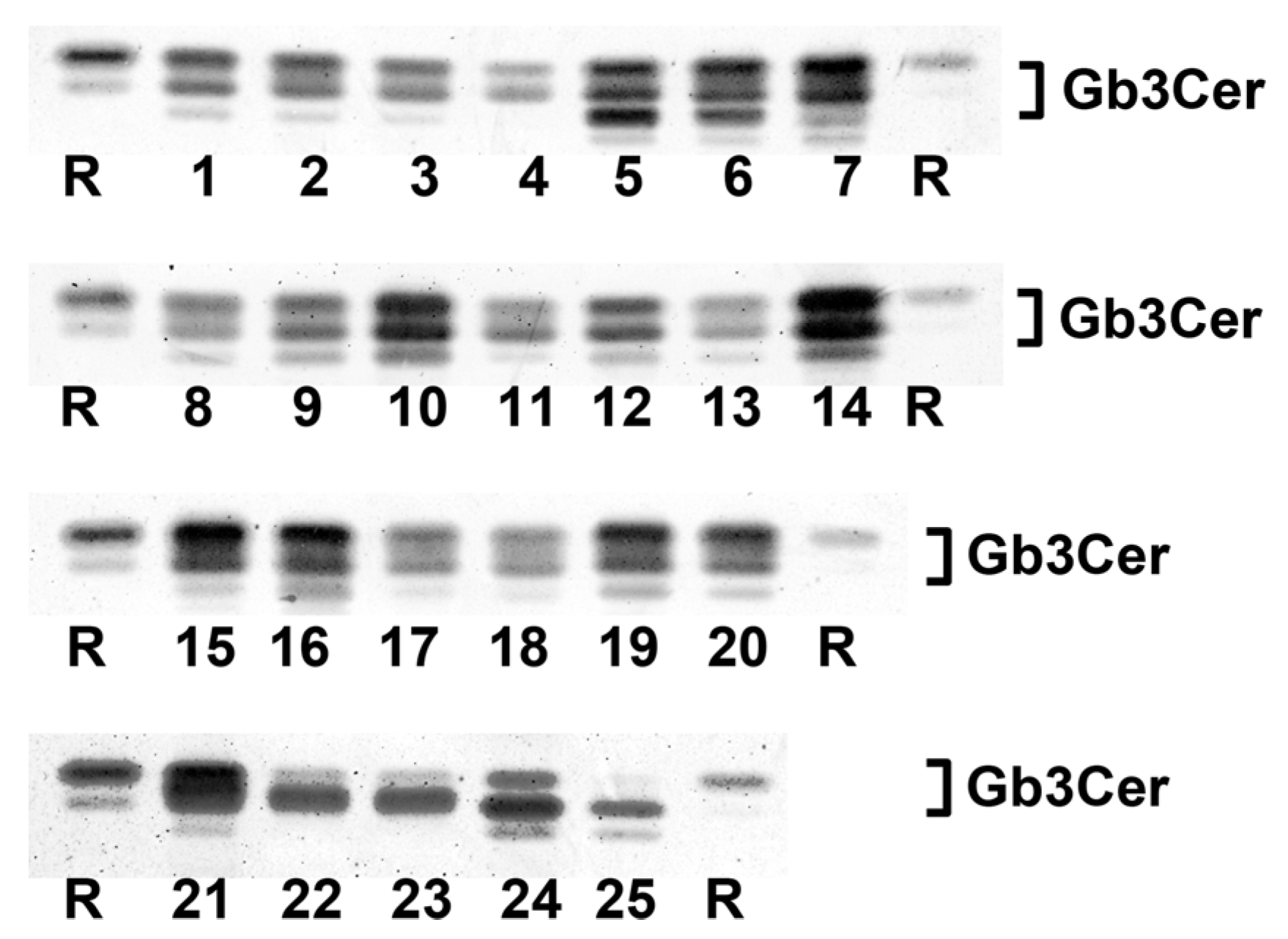
2.1. Antibody-Mediated Detection of Gb3Cer in Tissues and Organs of Male Piglet
2.2. Gb3Cer Lipoforms in Tissues and Organs of Male Piglet
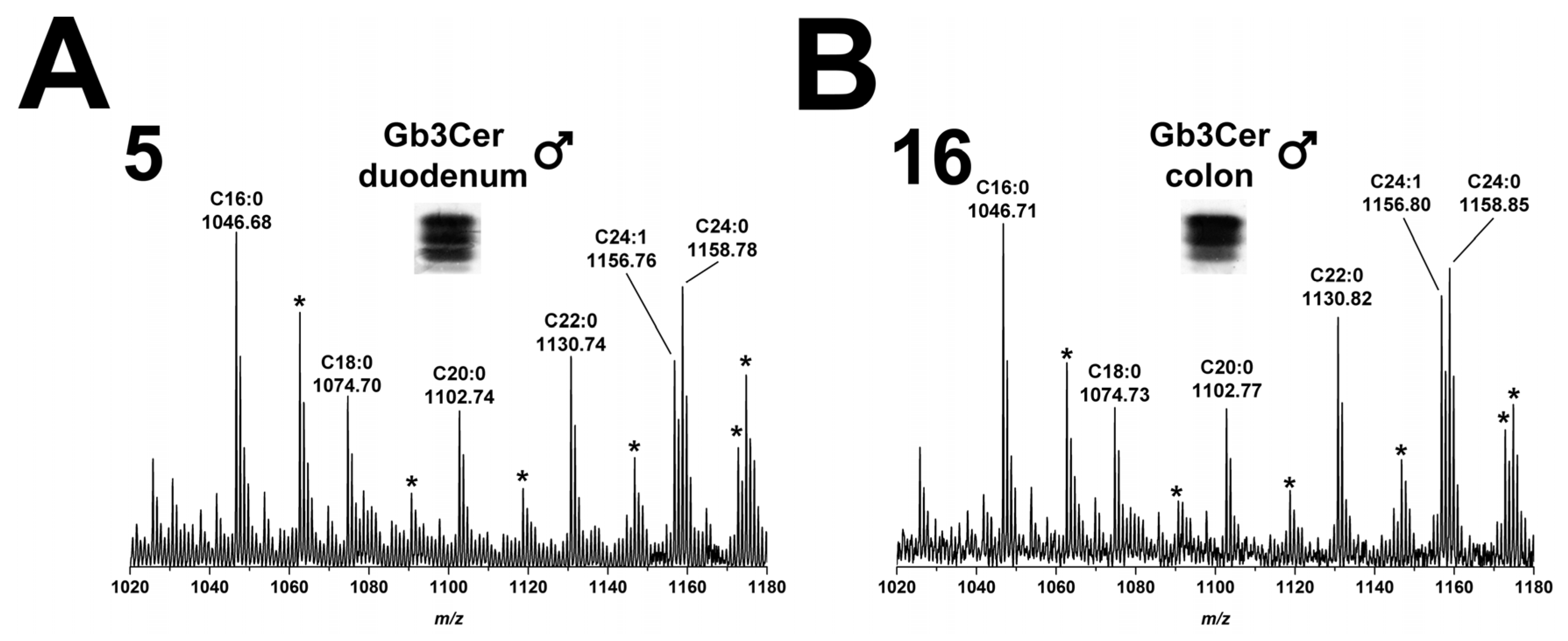
2.2.1. Gb3Cer Lipoforms in Duodenum and Colon
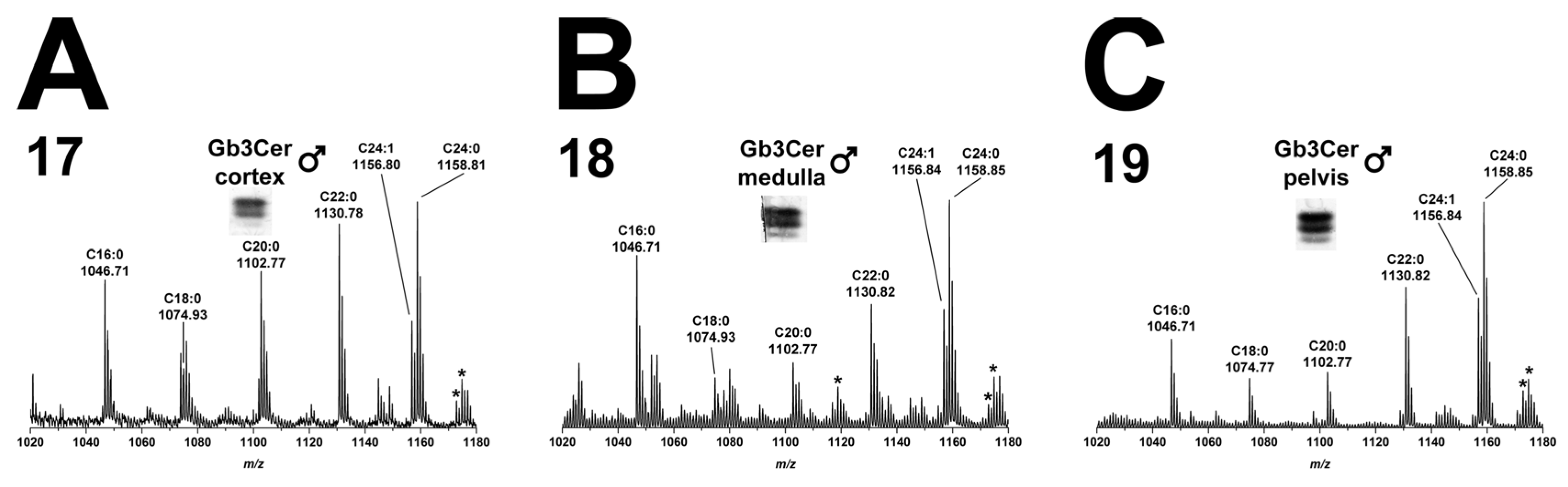
2.2.2. Gb3Cer Lipoforms in Kidney Cortex, Medulla, and Pelvis
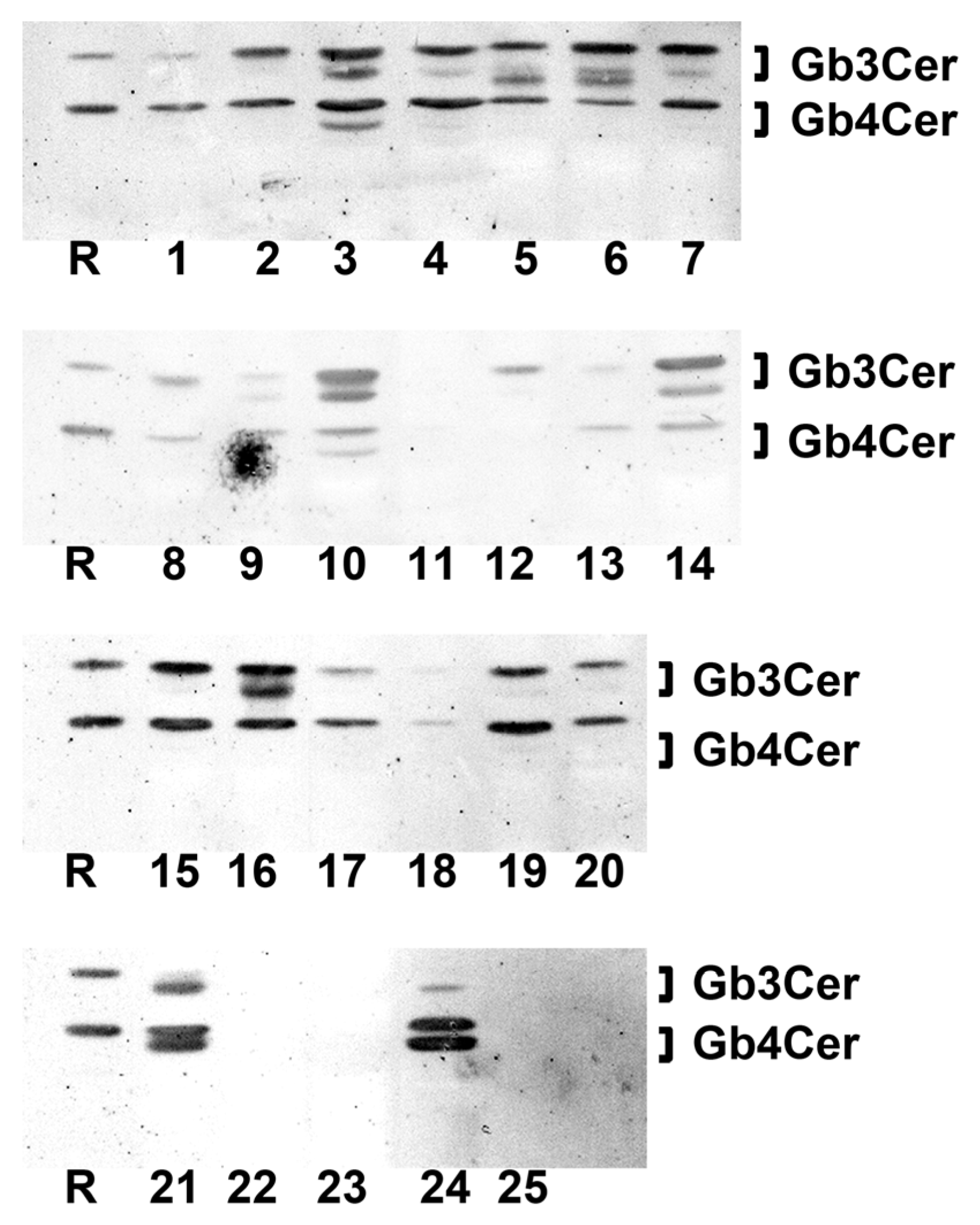
2.3. Antibody-Mediated Detection of Gb4Cer in Tissues and Organs of Male Piglet
2.4. Antibody-Mediated Detection of Forssman GSL in Tissues and Organs of Male Piglet
2.5. Gb4Cer Lipoforms in Tissues and Organs of Male Piglet
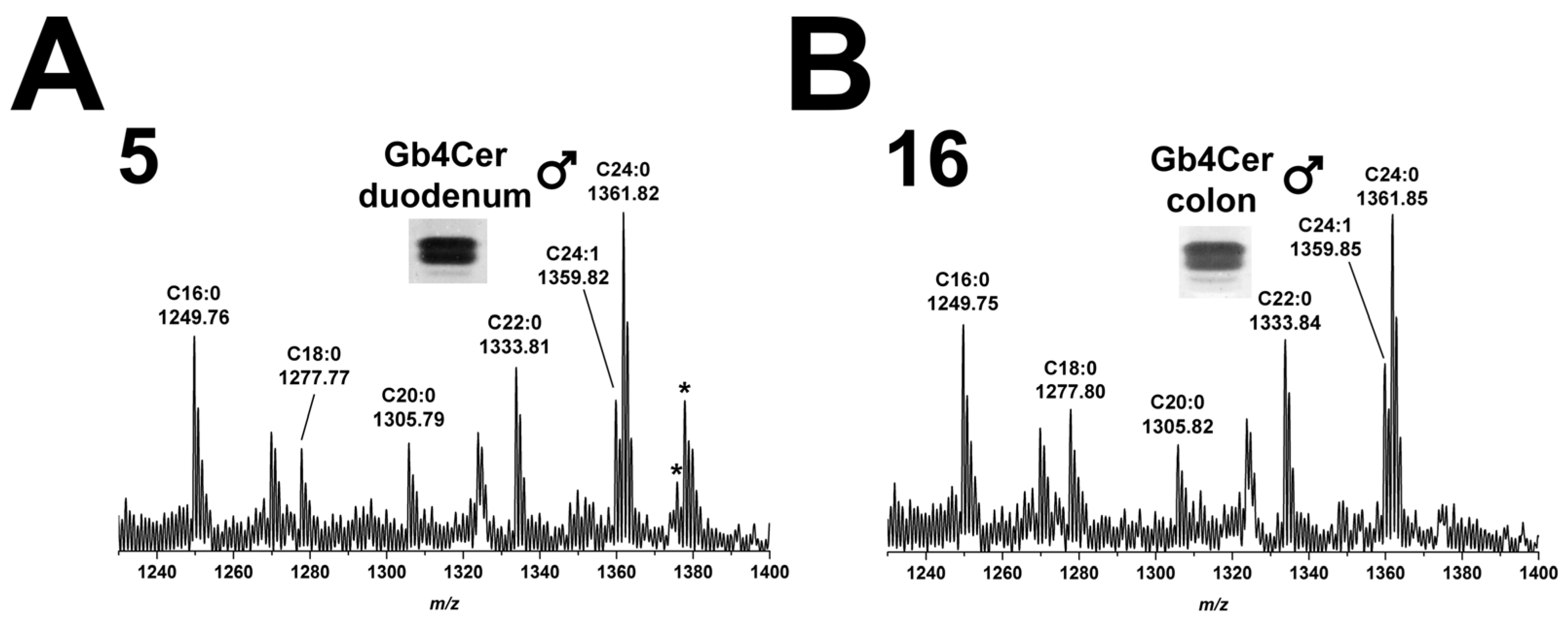
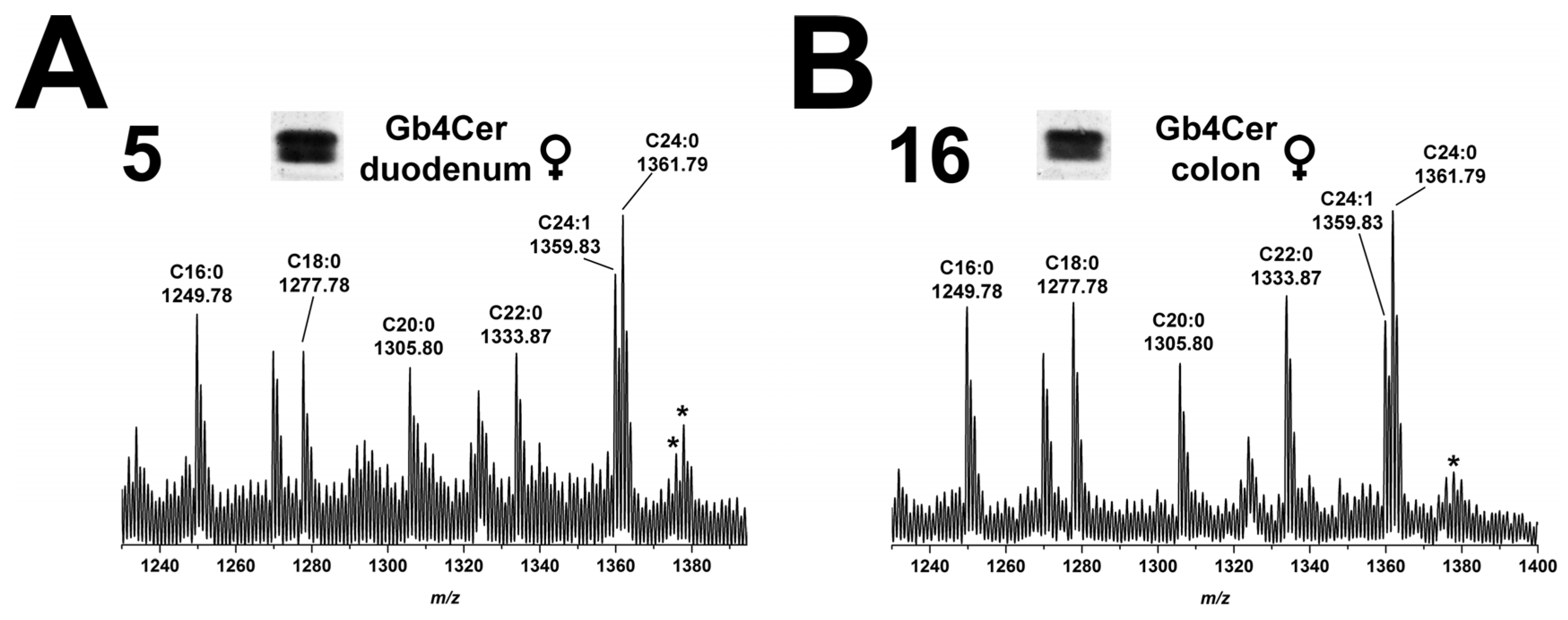
2.5.1. Gb4Cer Lipoforms of Duodenum and Colon
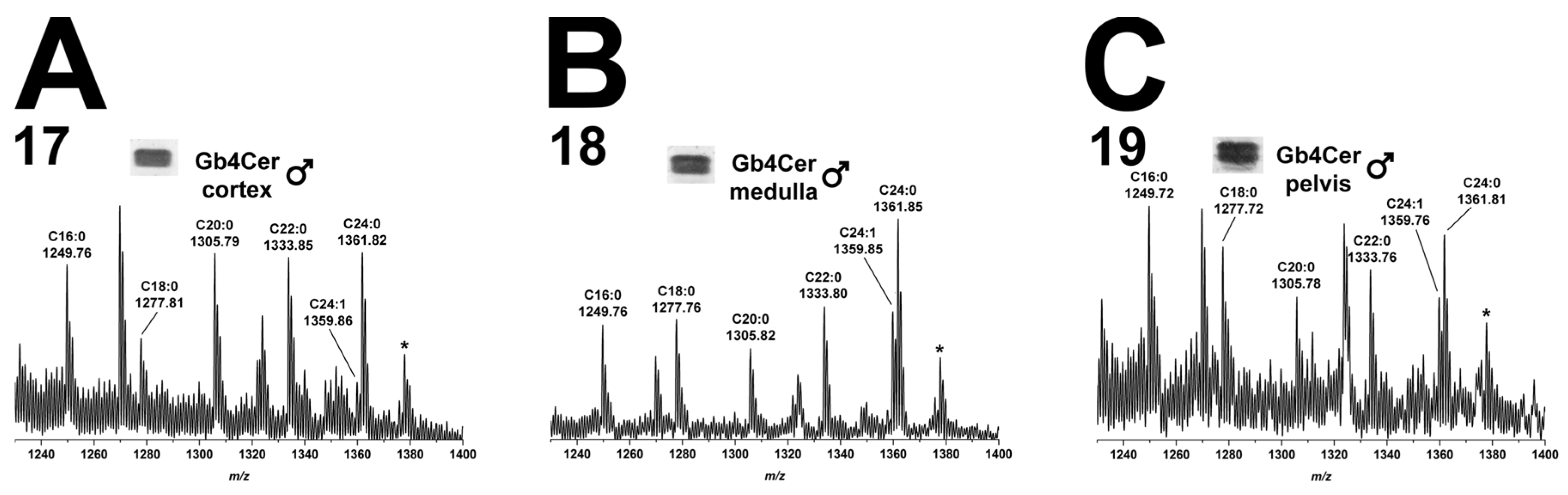
2.5.2. Gb4Cer Lipoforms in Kidney Cortex, Medulla, and Pelvis
2.6. Stx2e-Mediated Detection of Stx2e Receptors Gb3Cer and Gb4Cer in Tissues and Organs of Male Piglet
2.7. Antibody-Mediated Detection of Gb3Cer in Tissues and Organs of Female Piglet
2.8. Gb3Cer Lipoforms in Tissues and Organs of Female Piglet
2.8.1. Gb3Cer Lipoforms in Duodenum and Colon
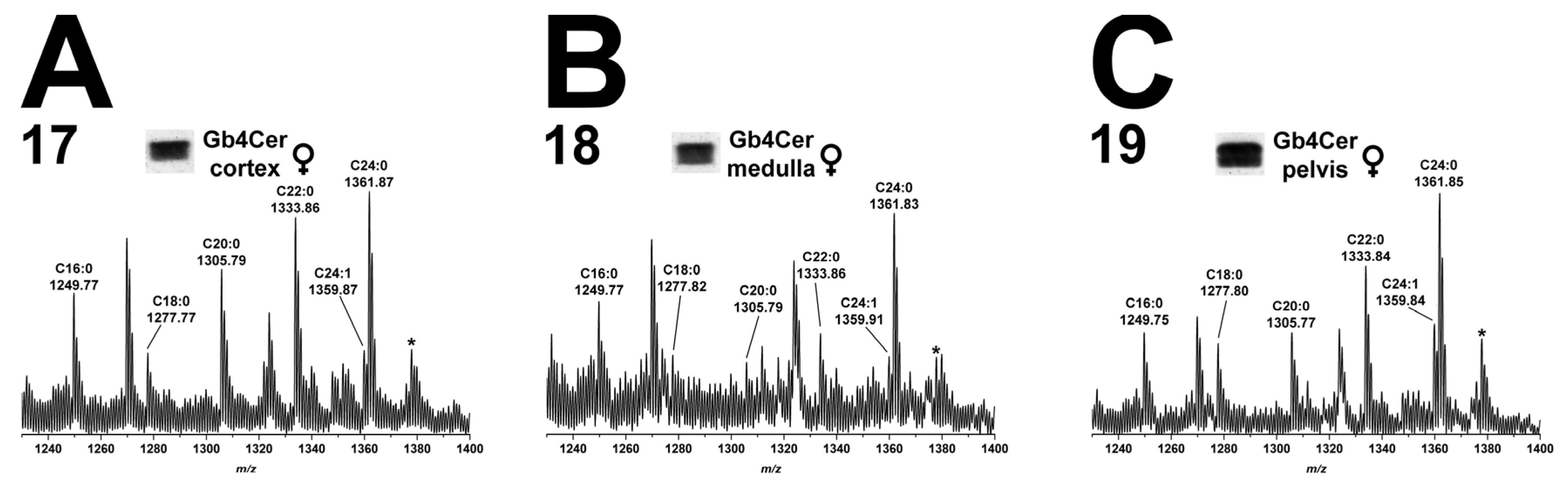
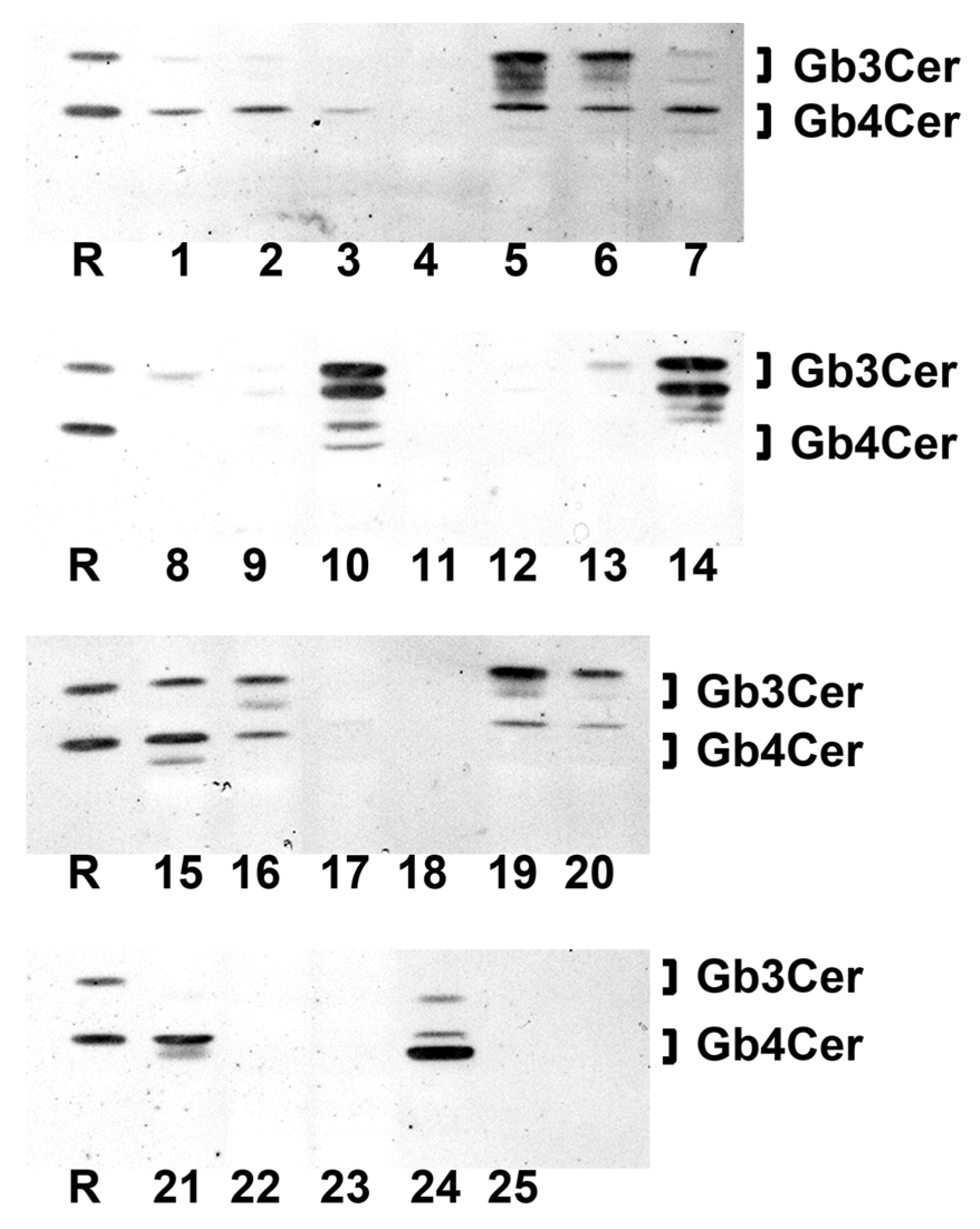
2.8.2. Gb3Cer Lipoforms in Kidney Cortex, Medulla, and Pelvis
2.9. Antibody-Mediated Detection of Gb4Cer in Tissues and Organs of Female Piglet
2.10. Gb4Cer Lipoforms in Tissues and Organs of Female Piglet
2.10.1. Gb4Cer Lipoforms of Duodenum and Colon
2.10.2. Gb4Cer Lipoforms in Kidney Cortex, Medulla, and Pelvis
2.11. Stx2e-Mediated Detection of Stx2e Receptors Gb3Cer and Gb4Cer in Tissues and Organs of Female Piglet
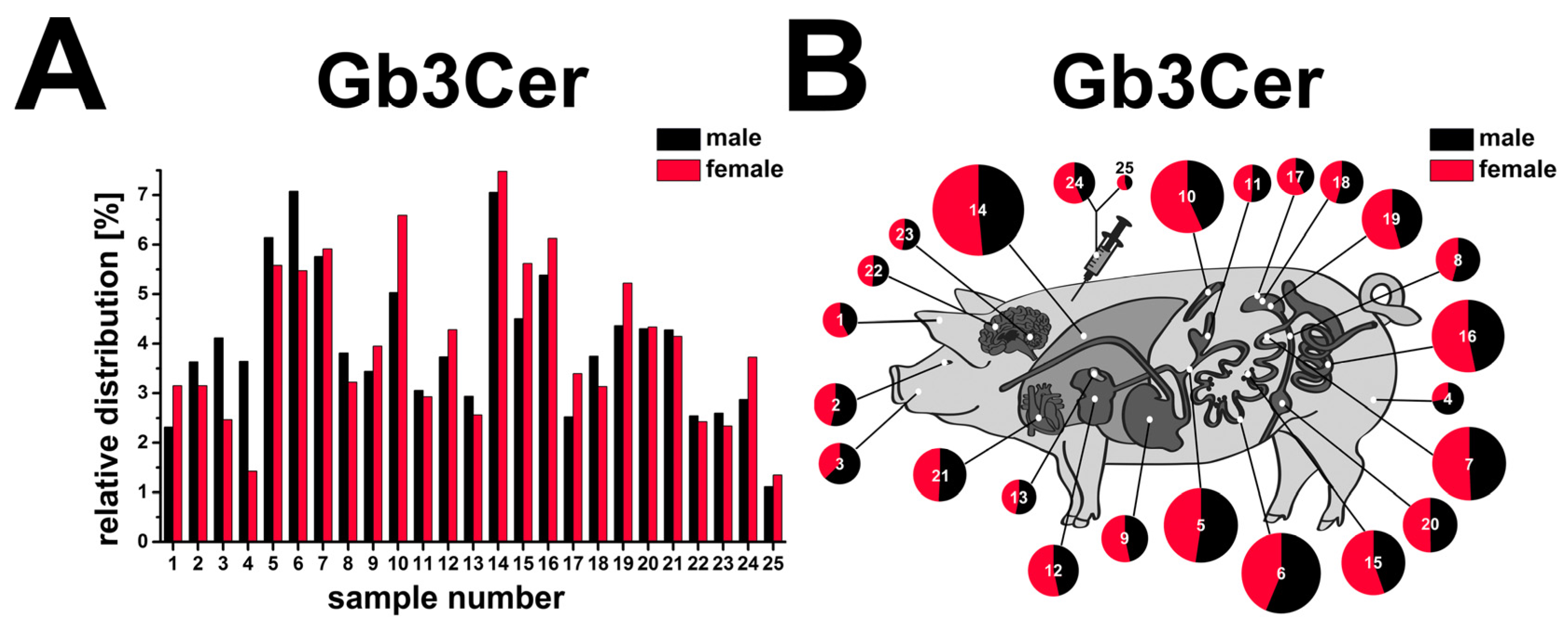
2.12. Compiled Distribution of Antibody-Detected Stx2e Receptor Gb3Cer in Tissues and Organs of Male and Female Piglet
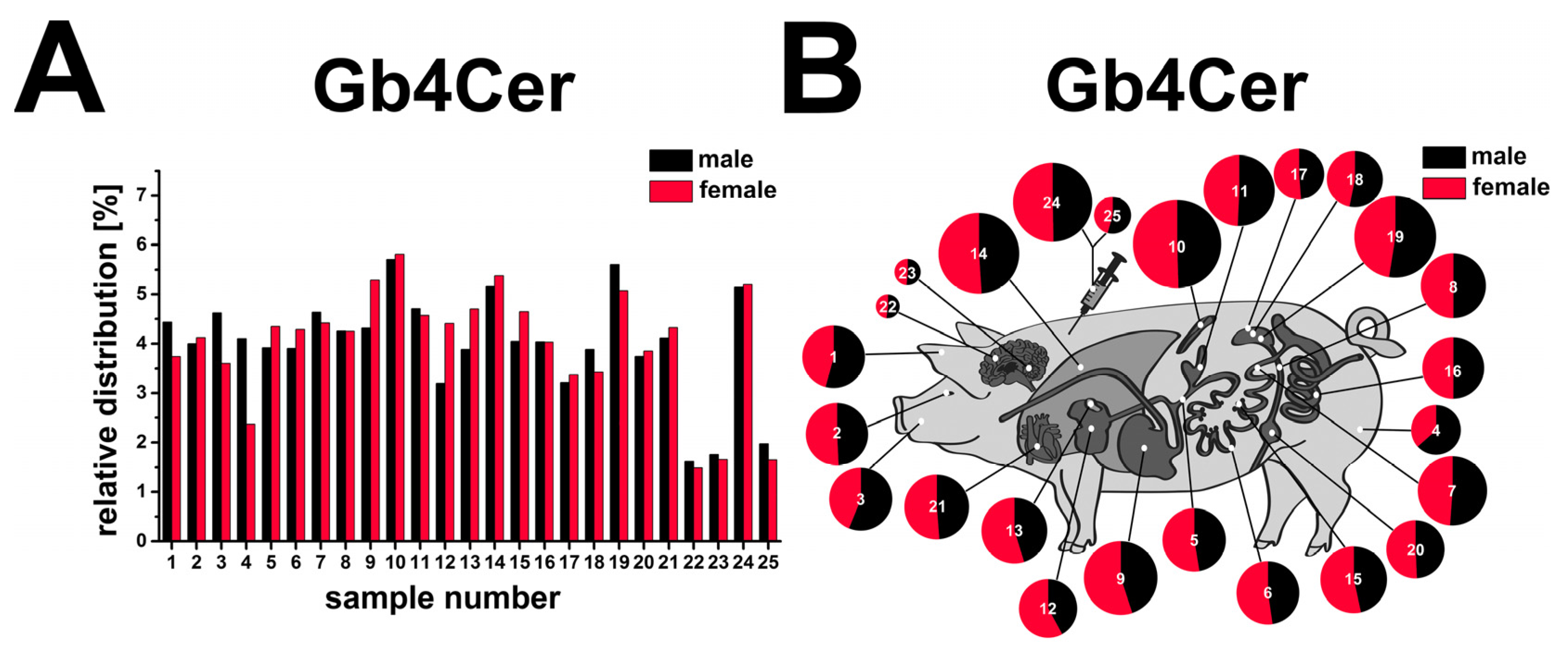
2.13. Compiled Distribution of Antibody-Detected Stx2e Receptor Gb4Cer in Tissues and Organs of Male and Female Piglet
3. Discussion
4. Materials and Methods
4.1. Pig Tissues and Organs
4.2. Preparation of GSL Extracts
4.3. Antibodies and Stx2e
4.4. Reference Glycosphingolipids (GSLs)
4.5. High-Performance Thin-Layer Chromatography (TLC) of GSLs
4.6. TLC Overlay Assay Detection of GSLs with Antibodies and Stx2e
4.7. Extraction of GSLs from Silica Gel of Immunostained TLC Bands
4.8. Mass Spectrometry
4.9. Statistics
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Scheutz, F.; Teel, L.D.; Beutin, L.; Piérard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [PubMed]
- Waddell, T.E.; Lingwood, C.A.; Gyles, C.L. Interaction of verotoxin 2e with pig intestine. Infect. Immun. 1996, 64, 1714–1719. [Google Scholar] [PubMed]
- Mainil, J. Shiga/verocytotoxins and Shiga/verotoxigenic Escherichia coli in animals. Vet. Res. 1999, 30, 235–257. [Google Scholar] [PubMed]
- Moxley, R.A. Edema disease. Vet. Clin. N. Am. Food Anim. Pract. 2000, 16, 175–185. [Google Scholar] [CrossRef]
- Tseng, M.; Fratamico, P.M.; Manning, S.D.; Funk, J.A. Shiga toxin-producing Escherichia coli in swine: The public health perspective. Anim. Health Res. Rev. 2014, 15, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Zoja, C.; Buelli, S.; Morigi, M. Shiga toxin-associated hemolytic uremic syndrome: Pathophysiology of endothelial dysfunction. Pediatr. Nephrol. 2010, 25, 2231–2240. [Google Scholar] [CrossRef] [PubMed]
- Trachtman, H.; Austin, C.; Lewinski, M.; Stahl, R.A. Renal and neurological involvement in typical Shiga toxin-associated HUS. Nat. Rev. Nephrol. 2012, 8, 658–669. [Google Scholar] [CrossRef] [PubMed]
- Orth-Höller, D.; Würzner, R. Role of complement in enterohemorrhagic Escherichia coli-induced hemolytic uremic syndrome. Semin. Thromb. Hemost. 2014, 40, 503–507. [Google Scholar] [PubMed]
- Caprioli, A.; Morabito, S.; Brugère, H.; Oswald, E. Enterohemorrhagic Escherichia coli: Emerging issues on virulence and modes of transmission. Vet. Res. 2005, 36, 289–311. [Google Scholar] [CrossRef] [PubMed]
- Piérard, D.; De Greve, H.; Haesebrouck, F.; Mainil, J. O157:H7 and O104:H4 Vero/Shiga toxin-producing Escherichia coli outbreaks: Respective role of cattle and humans. Vet. Res. 2012, 43, 13. [Google Scholar] [CrossRef] [PubMed]
- Moxley, R.A.; Acuff, G.R. Peri- and postharvest factors in the control of Shiga toxin-producing Escherichia coli in beef. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Menge, C.; Stamm, I.; Wuhrer, M.; Geyer, R.; Wieler, L.H.; Baljer, G. Globotriaosylceramide (Gb(3)/CD77) is synthesized and surface expressed by bovine lymphocytes upon activation in vitro. Vet. Immunol. Immunopathol. 2001, 83, 19–36. [Google Scholar] [CrossRef]
- Stamm, I.; Wuhrer, M.; Geyer, R.; Baljer, G.; Menge, C. Bovine lymphocytes express functional receptors for Escherichia coli Shiga toxin 1. Microb. Pathog. 2002, 33, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Menge, C.; Blessenohl, M.; Eisenberg, T.; Stamm, I.; Baljer, G. Bovine ileal intraepithelial lymphocytes represent target cells for Shiga toxin 1 from Escherichia coli. Infect. Immun. 2004, 72, 1896–1905. [Google Scholar] [CrossRef] [PubMed]
- Wieler, L.H.; Bauerfeind, R. STEC as a veterinary problem. Diagnostics and prophylaxis in animals. Methods Mol. Med. 2003, 73, 75–89. [Google Scholar] [PubMed]
- Meng, Q.; Bai, X.; Zhao, A.; Lan, R.; Du, H.; Wang, T.; Shi, C.; Yuan, X.; Bai, X.; Ji, S.; et al. Characterization of Shiga toxin-producing Escherichia coli isolated from healthy pigs in China. BMC Microbiol. 2014, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Winter, K.R.K.; Stoffregen, W.C.; Dean-Nystrom, E.A. Shiga toxin binding to isolated porcine tissues and peripheral blood leukocytes. Infect. Immun. 2004, 72, 6680–6684. [Google Scholar] [CrossRef] [PubMed]
- Pohlenz, J.F.; Winter, K.R.; Dean-Nystrom, E.A. Shiga-toxigenic Escherichia coli-inoculated neonatal piglets develop kidney lesions that are comparable to those in humans with hemolytic-uremic syndrome. Infect. Immun. 2005, 73, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Beddoe, T.; Paton, A.W.; Le Nours, J.; Rossjohn, J.; Paton, J.C. Structure, biological functions and applications of the AB5 toxins. Trends Biochem. Sci. 2010, 35, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Bergan, J.; Dyve Lingelem, A.B.; Simm, R.; Skotland, T.; Sandvig, K. Shiga toxins. Toxicon 2012, 60, 1085–1107. [Google Scholar] [CrossRef] [PubMed]
- Melton-Celsa, A.R. Shiga toxin (Stx) classification, structure, and function. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Bergan, J.; Dyve, A.B.; Skotland, T.; Torgersen, M.L. Endocytosis and retrograde transport of Shiga toxin. Toxicon 2010, 56, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Lingwood, C.A. Role of verotoxin receptors in pathogenesis. Trends Microbiol. 1996, 4, 147–153. [Google Scholar] [CrossRef]
- Nakajima, H.; Kiyokawa, N.; Katagiri, Y.U.; Taguchi, T.; Suzuki, T.; Sekino, T.; Mimori, K.; Ebata, T.; Saito, M.; Nakao, H.; et al. Kinetic analysis of binding between Shiga toxin and receptor glycolipid Gb3Cer by surface plasmon resonance. J. Biol. Chem. 2001, 276, 42915–42922. [Google Scholar] [CrossRef] [PubMed]
- Müthing, J.; Meisen, I.; Zhang, W.; Bielaszewska, M.; Mormann, M.; Bauerfeind, R.; Schmidt, M.A.; Friedrich, A.W.; Karch, H. Promiscuous Shiga toxin 2e and its intimate relationship to Forssman. Glycobiology 2012, 22, 849–862. [Google Scholar] [CrossRef] [PubMed]
- DeGrandis, S.; Law, H.; Brunton, J.; Gyles, C.; Lingwood, C.A. Globotetraosylceramide is recognized by the pig edema disease toxin. J. Biol. Chem. 1989, 264, 12520–12525. [Google Scholar] [PubMed]
- Keusch, G.T.; Jacewicz, M.; Acheson, D.W.K.; Donohue-Rolfe, A.; Kane, A.V.; McCluer, R.H. Globotriaosylceramide, Gb3, is an alternative functional receptor for Shiga-like toxin 2e. Infect. Immun. 1995, 63, 1138–1141. [Google Scholar] [PubMed]
- Lencer, W.I.; Saslowsky, D. Raft trafficking of AB5 subunit bacterial toxins. Biochim. Biophys. Acta 2005, 1746, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Römer, W.; Berland, L.; Chambon, V.; Gaus, K.; Windschiegl, B.; Tenza, D.; Aly, M.R.; Fraisier, V.; Florent, J.C.; Perrais, D.; et al. Shiga toxin induces tubular membrane invaginations for its uptake into cells. Nature 2007, 450, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Sens, P.; Johannes, L.; Bassereau, P. Biophysical approaches to protein-induced membrane deformations in trafficking. Curr. Opin. Cell Biol. 2008, 20, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Ewers, H.; Helenius, A. Lipid-mediated endocytosis. Cold Spring Harb. Perspect. Biol. 2011, 3, a004721. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Cherla, R.P.; Tesh, V.L. Shiga toxins: Intracellular trafficking to the ER leading to activation of host cell stress responses. Toxins 2010, 2, 1515–1535. [Google Scholar] [CrossRef] [PubMed]
- Brigotti, M.; Alfieri, R.; Sestili, P.; Bonelli, M.; Petronini, P.G.; Guidarelli, A.; Barbieri, L.; Stirpe, F.; Sperti, S. Damage to nuclear DNA induced by Shiga toxin 1 and ricin in human endothelial cells. FASEB J. 2002, 16, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Sestili, P.; Alfieri, R.; Carnicelli, D.; Martinelli, C.; Barbieri, L.; Stirpe, F.; Bonelli, M.; Petronini, P.G.; Brigotti, M. Shiga toxin 1 and ricin inhibit the repair of H2O2-induced DNA single strand breaks in cultured mammalian cells. DNA Repair 2005, 4, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Schweppe, C.H.; Bielaszewska, M.; Pohlentz, G.; Friedrich, A.W.; Büntemeyer, H.; Schmidt, M.A.; Kim, K.S.; Peter-Katalinić, J.; Karch, H.; Müthing, J. Glycosphingolipids in vascular endothelial cells: Relationship of heterogeneity in Gb3Cer/CD77 receptor expression with differential Shiga toxin 1 cytotoxicity. Glycoconj. J. 2008, 25, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Betz, J.; Bauwens, A.; Kunsmann, L.; Bielaszewska, M.; Mormann, M.; Humpf, H.U.; Karch, H.; Friedrich, A.W.; Müthing, J. Uncommon membrane distribution of Shiga toxin glycosphingolipid receptors in toxin-sensitivie human glomerular microvascular endothelial cells. Biol. Chem. 2012, 393, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Bauwens, A.; Betz, J.; Meisen, I.; Kemper, B.; Karch, H.; Müthing, J. Facing glycosphingolipid-Shiga toxin interaction: Dire straits for endothelial cells of the human vasculature. Cell. Mol. Life Sci. 2013, 70, 425–457. [Google Scholar] [CrossRef] [PubMed]
- Meisen, I.; Rosenbrück, R.; Galla, H.J.; Hüwel, S.; Kouzel, I.U.; Mormann, M.; Karch, H.; Müthing, J. Expression of Shiga toxin 2e glycosphingolipid receptors of primary porcine brain endothelial cells and toxin-mediated breakdown of the blood-brain barrier. Glycobiology 2013, 23, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Boyd, B.; Tyrrell, G.; Maloney, M.; Gyles, C.; Brunton, J.; Lingwood, C. Alteration of the glycolipid binding specificity of the pig edema toxin from globotetraosyl to globotriaosyl ceramide alters in vivo tissue targeting and results in verotoxin 1-like disease in pigs. J. Exp. Med. 1993, 177, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Waddell, T.E.; Coomber, B.L.; Gyles, C.L. Localization of potential binding sites for the edema disease Verotoxin (VT2e) in pigs. Can. J. Vet. Res. 1998, 62, 81–86. [Google Scholar] [PubMed]
- Bethke, U.; Müthing, J.; Schauder, B.; Conradt, P.; Mühlradt, P.F. An improved semi-quantitative nzyme immunostaining procedure for glycosphingolipid antigens on high performance thin layer chromatograms. J. Immunol. Methods 1986, 89, 111–116. [Google Scholar] [CrossRef]
- Distler, U.; Souady, J.; Hülsewig, M.; Drmić-Hofman, I.; Haier, J.; Friedrich, A.W.; Karch, H.; Senninger, N.; Dreisewerd, K.; Berkenkamp, S.; et al. Shiga toxin receptor Gb3Cer/CD77: Tumor association and promising therapeutic target in pancreas and colon cancer. PLoS ONE 2009, 4, e6813. [Google Scholar] [CrossRef] [PubMed]
- Kouzel, I.U.; Pohlentz, G.; Storck, W.; Radamm, L.; Hoffmann, P.; Bielaszewska, M.; Bauwens, A.; Cichon, C.; Schmidt, M.A.; Mormann, M.; et al. Association of Shiga toxin glycosphingolipid receptors with membrane microdomains of toxin-sensitive lymphoid and myeloid cells. J. Lipid Res. 2013, 51, 692–710. [Google Scholar] [CrossRef] [PubMed]
- Betz, J.; Dorn, I.; Kouzel, I.U.; Bauwens, A.; Meisen, I.; Kemper, B.; Bielaszewska, M.; Mormann, M.; Weymann, L.; Sibrowski, W.; et al. Shiga toxin of enterohemorrhagic Escherichia coli directly injures developing human erythrocytes. Cell. Microbiol. 2016, 18, 1339–1348. [Google Scholar] [CrossRef] [PubMed]
- Binnington, B.; Lingwood, D.; Nutikka, A.; Lingwood, C.A. Effect of globotriaosyl ceramide fatty acid α-hydroxylation on the binding of verotoxin 1 and verotoxin 2. Neurochem. Res. 2002, 27, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Coddens, A.; Valis, E.; Benktander, J.; Angström, J.; Breimer, M.E.; Cox, E.; Teneberg, S. Erythrocyte and porcine intestinal glycosphingolipids recognized by F4 fimbriae of enterotoxigenic Escherichia coli. PLoS ONE 2011, 6, e23309. [Google Scholar] [CrossRef] [PubMed]
- Madar Johannson, M.; Coddens, A.; Benktander, J.; Cox, E.; Teneberg, S. Porcine intestinal glycosphingolipids recognized by F6-fimbriated enterotoxigenic Escherichia coli. Microb. Pathog. 2014, 76, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Alsén, B.; Roche, N.; Angström, J.; von Euler, A.; Breimer, M.E.; Westerlund-Wikström, B.; Teneberg, S.; Richter-Dahlfors, A. Identification of target tissue glycosphingolipid receptors for uropathogenic, F1C-fimbriated Escherichia coli and its role in mucosal inflammation. J. Biol. Chem. 2002, 277, 18198–18205. [Google Scholar] [CrossRef] [PubMed]
- Breimer, M.E.; Hansson, G.C.; Karlsson, K.A.; Leffler, H. Blood group type glycosphingolipids from the small intestine of different animals analysed by mass spectrometry and thin-layer chromatography. A note on species diversity. J. Biochem. 1981, 90, 589–609. [Google Scholar] [PubMed]
- Yamamoto, M.; Cid, E.; Yamamoto, F. Molecular genetic basis of the human Forssman glycolipid antigen negativity. Sci. Rep. 2012, 2, 975. [Google Scholar] [CrossRef] [PubMed]
- Svensson, L.; Hult, A.K.; Stamps, R.; Ångström, J.; Teneberg, S.; Storry, J.R.; Jørgensen, R.; Rydberg, L.; Henry, S.M.; Olsson, M.L. Forssman expression on human erythrocytes: Biochemical and genetic evidence of a new histo-blood group system. Blood 2013, 121, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Breimer, M.E. Chemical and immunological identification of the Forssman pentaglycosylceramide in human kidney. Glycoconj. J. 1985, 2, 375–385. [Google Scholar]
- Matise, I.; Cornick, N.A.; Samuel, J.E.; Moon, H.W. Binding of Shiga toxin 2e to porcine erythrocytes in vivo and in vitro. Infect. Immun. 2003, 71, 5194–5201. [Google Scholar] [CrossRef] [PubMed]
- Obata, F. Influence of Escherichia coli Shiga toxin on the mammalian central nervous system. Adv. Appl. Microbiol. 2010, 7, 1–19. [Google Scholar]
- Matise, I.; Sirinarumitr, T.; Bosworth, B.T.; Moon, H.W. Vascular ultrastructure and DNA fragmentation in swine infected with Shiga toxin-producing Escherichia coli. Vet. Pathol. 2000, 37, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Obata, F.; Obrig, T. Role of Shiga/Vero toxins in pathogenesis. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Karch, H.; Friedrich, A.W.; Gerber, A.; Zimmerhackl, L.B.; Schmidt, M.A.; Bielaszewska, M. New aspects in the pathogenesis of enteropathic hemolytic uremic syndrome. Semin. Thromb. Hemost. 2006, 32, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Kunsmann, L.; Rüter, C.; Bauwens, A.; Greune, L.; Glüder, M.; Kemper, B.; Fruth, A.; Wai, S.N.; He, X.; Lloubes, R.; et al. Virulence from vesicles: Novel mechanisms of host cell injury by Escherichia coli O104:H4 outbreak strain. Sci. Rep. 2015, 5, 13252. [Google Scholar] [CrossRef] [PubMed]
- Piérard, D.; Huyghens, L.; Lauwers, S. Diarrhoea associated with Escherichia coli producing porcine oedema disease verotoxin. Lancet 1991, 338, 762. [Google Scholar] [CrossRef]
- Thomas, A.; Cheasty, T.; Chart, H.; Rowe, B. Isolation of Vero cytotoxin-producing Escherichia coli serotypes O9ab:H- and O101:H-carrying VT2 variant gene sequences from a patient with haemolytic uraemic syndrome. Eur. J. Clin. Microbiol. Infect. Dis. 1994, 13, 1074–1076. [Google Scholar] [CrossRef] [PubMed]
- Fasel, D.; Mellmann, A.; Cernela, N.; Hächler, H.; Fruth, A.; Khanna, N.; Egli, A.; Beckmann, C.; Hirsch, H.H.; Goldenberger, D.; et al. Hemolytic uremic syndrome in a 65-year old male linked to a very unusual type of stx2e- and eae-harboring O51:H49 Shiga toxin-producing Escherichia coli. J. Clin. Microbiol. 2014, 52, 1301–1303. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, A.W.; Bielaszewska, M.; Zhang, W.L.; Pulz, M.; Kuczius, T.; Ammon, A.; Karch, H. Escherichia coli harboring Shiga toxin 2 gene variants: Frequency and association with clinical symptoms. J. Infect. Dis. 2002, 185, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Beutin, L.; Krause, G.; Zimmermann, S.; Kaulfuss, S.; Gleier, K. Characterization of Shiga toxin-producing Escherichia coli strains isolated from human patients in Germany over a 3-year period. J. Clin. Microbiol. 2004, 42, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Sonntag, A.K.; Bielaszewska, M.; Mellmann, A.; Dierksen, N.; Schierack, P.; Wieler, L.H.; Schmidt, M.A.; Karch, H. Shiga toxin 2e-producing Escherichia coli isolates from humans and pigs differ in their virulence profiles and interactions with intestinal epithelial cells. Appl. Environ. Microbiol. 2005, 71, 8855–8863. [Google Scholar] [CrossRef] [PubMed]
- Ercoli, L.; Farneti, S.; Ranucci, D.; Scuota, S.; Branclari, R. Role of verocytotoxigenic Escherichia coli in the swine production chain. Int. J. Food Saf. 2015, 4, 5156. [Google Scholar] [CrossRef] [PubMed]
- Fratamico, P.M.; Bagi, L.K.; Bush, E.J.; Solow, B.T. Prevalence and characterization of Shiga toxin-producing Escherichia coli in swine feces recovered in the National Animal Health Monitoring System’s Swine 2000 study. Appl. Environ. Microbiol. 2004, 70, 7173–7178. [Google Scholar] [CrossRef] [PubMed]
- Tseng, M.; Fratamico, P.M.; Bagi, L.; Delannoy, S.; Fach, P.; Manning, S.D.; Funk, J.A. Diverse virulence gene content of Shiga toxin-producing Escherichia coli from finishing swine. Appl. Environ. Microbiol. 2014, 80, 6395–6402. [Google Scholar] [CrossRef] [PubMed]
- Tseng, M.; Fratamico, P.M.; Bagi, L.; Manzinger, D.; Funk, J.A. Shiga toxin-producing E. coli (STEC) in swine: Prevalence over the finishing period and characteristics of the STEC isolates. Epidemiol. Infect. 2015, 143, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Baranzoni, G.M.; Fratamico, P.M.; Gangiredla, J.; Patel, I.; Bagi, L.K.; Delannoy, S.; Fach, P.; Boccia, F.; Anastasio, A.; Pepe, T. Characterization of Shiga toxin subtypes and virulence genes in porcine Shiga toxin-producing Escherichia coli. Front. Microbiol. 2016, 7, 574. [Google Scholar] [CrossRef] [PubMed]
- Arimitsu, H.; Sasaki, K.; Kohda, T.; Shimizu, T.; Tsuji, T. Evaluation of Shiga toxin 2e-specific chicken egg yolk immunoglobulin: Production and neutralization activity. Microbiol. Immunol. 2014, 58, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Arimitsu, H.; Sasaki, K.; Tsuji, T. Development of a simple and rapid diagnosis method for swine edema disease to specifically detect Stx2e protein by immunochromatographic test. Microbiol. Immunol. 2016, 60, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Caprioli, A.; Scavia, G.; Morabito, S. Public health microbiology of Shiga toxin-producing Escherichia coli. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Mauro, S.A.; Koudelka, G.B. Shiga toxin: Expression, distribution, and its role in the environment. Toxins (Basel) 2011, 3, 608–625. [Google Scholar] [CrossRef] [PubMed]
- Schweppe, C.H.; Hoffmann, P.; Nofer, J.R.; Pohlentz, G.; Mormann, M.; Karch, H.; Friedrich, A.W.; Müthing, J. Neutral glycosphingolipids in human blood: A precise mass spectrometry analysis with special reference to lipoprotein-associated Shiga toxin receptors. J. Lipid Res. 2010, 51, 2282–2294. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, P.; Hülsewig, M.; Duvar, S.; Ziehr, H.; Mormann, M.; Peter-Katalinić, J.; Friedrich, A.W.; Karch, H.; Müthing, J. On the structural diversity of Shiga toxin glycosphingolipid receptors in lymphoid and myeloid cells determined by nanoelectrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2295–2304. [Google Scholar] [CrossRef] [PubMed]
- Bethke, U.; Kniep, B.; Mühlradt, P.F. Forssman glycolipid, an antigenic marker for a major subpopulation of macrophages from murine spleen and peripheral lymph nodes. J. Immunol. 1987, 138, 4329–4335. [Google Scholar] [PubMed]
- Chester, M.A. IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). Nomenclature of glycolipids-recommendations 1997. Eur. J. Biochem. 1998, 257, 293–298. [Google Scholar] [PubMed]
- Distler, U.; Hülsewig, M.; Souady, J.; Dreisewerd, K.; Haier, J.; Senninger, N.; Friedrich, A.W.; Karch, H.; Hillenkamp, F.; Berkenkamp, S.; et al. Matching IR-MALDI-o-TOF mass spectrometry with the TLC overlay binding assay and its clinical application for tracing tumor-associated glycosphingolipids in hepatocellular and pancreatic cancer. Anal. Chem. 2008, 80, 1835–1846. [Google Scholar] [CrossRef] [PubMed]
- Storck, W.; Meisen, I.; Gianmoena, K.; Pläger, I.; Kouzel, I.U.; Bielaszewska, M.; Haier, J.; Mormann, M.; Humpf, H.U.; Karch, H.; et al. Shiga toxin glycosphingolipid receptor expression and toxin susceptibility of human pancreatic ductal adenocarcinomas of differing origin and differentiation. Biol. Chem. 2012, 393, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Kouzel, I.U.; Pirkl, A.; Pohlentz, G.; Soltwisch, J.; Dreisewerd, K.; Karch, H.; Müthing, J. Progress in detection and structural characterization of glycosphingolipids in crude lipid extracts by enzymatic phospholipid disintegration combined with thin-layer chromatography immunodetection and IR-MALDI mass spectrometry. Anal. Chem. 2014, 86, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Meisen, I.; Peter-Katalinić, J.; Müthing, J. Direct analysis of silica gel extracts from immunostained glycosphingolipids by nanoelectrospray ionization quadrupole time-of-flight mass spectrometry. Anal. Chem. 2004, 76, 2248–2255. [Google Scholar] [CrossRef] [PubMed]
- Domon, B.; Costello, C.E. A systematic nomenclature for carbohydrate fragmentations in FAB-MS/MS spectra of glycoconjugates. Glycoconj. J. 1988, 5, 397–440. [Google Scholar] [CrossRef]
- Domon, B.; Costello, C.E. Structure elucidation of glycosphingolipids and gangliosides using high-performance tandem mass spectrometry. Biochemistry 1988, 27, 1534–1543. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org/ (accessed on 14 July 2016).















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gb3Cer | Gb4Cer | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Rank a | No. b | Male | No. c | Female | No. b | Male | No. c | Female | |
| 1 | 6 | intestine jejunum | 14 | lung | 10 | spleen | 10 | spleen | I d |
| 2 | 14 | lung | 10 | spleen | 19 | kidney pelvis | 14 | lung | |
| 3 | 5 | intestine duodenum | 16 | intestine colon | 14 | lung | 9 | stomach | |
| 4 | 7 | intestine ileum | 7 | intestine ileum | 24 | whole blood f | 24 | whole blood f | |
| 5 | 16 | intestine colon | 15 | lymph nodes e | 11 | pancreas | 19 | kidney pelvis | |
| 6 | 10 | spleen | 5 | intestine duodenum | 7 | intestine ileum | 13 | gall bladder | |
| 7 | 15 | lymph nodes e | 6 | intestine jejunum | 3 | nasal bridge | 15 | lymph nodes e | |
| 8 | 19 | kidney pelvis | 19 | kidney pelvis | 1 | earlobe | 11 | pancreas | |
| 9 | 20 | urinary bladder | 20 | urinary bladder | 9 | stomach | 7 | intestine ileum | |
| 10 | 21 | heart | 12 | liver | 8 | ureter | 12 | liver | |
| 11 | 3 | nasal bridge | 21 | heart | 21 | heart | 5 | intestine duodenum | II d |
| 12 | 8 | ureter | 9 | stomach | 4 | quadriceps muscle | 21 | heart | |
| 13 | 18 | kidney medulla | 24 | whole blood f | 15 | Lymphnodes e | 6 | intestine jejunum | |
| 14 | 12 | liver | 17 | kidney cortex | 16 | intestine colon | 8 | ureter | |
| 15 | 4 | quadriceps muscle | 8 | ureter | 2 | eyelid | 2 | eyelid | |
| 16 | 2 | eyelid | 1 | earlobe | 5 | intestine duodenum | 16 | intestine colon | |
| 17 | 9 | stomach | 2 | eyelid | 6 | intestine jejunum | 20 | urinary bladder | |
| 18 | 11 | pancreas | 18 | kidney medulla | 13 | gall bladder | 1 | earlobe | |
| 19 | 13 | gall bladder | 11 | pancreas | 18 | kidney medulla | 3 | nasal bridge | |
| 20 | 24 | whole blood f | 13 | gall bladder | 20 | urinary bladder | 18 | kidney medulla | III d |
| 21 | 23 | cerebellum | 3 | nasal bridge | 17 | kidney cortex | 17 | kidney cortex | |
| 22 | 22 | cerebrum | 22 | cerebrum | 12 | liver | 4 | quadriceps muscle | |
| 23 | 17 | kidney cortex | 23 | cerebellum | 25 | serum | 23 | cerebellum | |
| 24 | 1 | earlobe | 4 | quadriceps muscle | 23 | cerebellum | 25 | serum | |
| 25 | 25 | serum | 25 | serum | 22 | cerebrum | 22 | cerebrum | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steil, D.; Bonse, R.; Meisen, I.; Pohlentz, G.; Vallejo, G.; Karch, H.; Müthing, J. A Topographical Atlas of Shiga Toxin 2e Receptor Distribution in the Tissues of Weaned Piglets. Toxins 2016, 8, 357. https://doi.org/10.3390/toxins8120357
Steil D, Bonse R, Meisen I, Pohlentz G, Vallejo G, Karch H, Müthing J. A Topographical Atlas of Shiga Toxin 2e Receptor Distribution in the Tissues of Weaned Piglets. Toxins. 2016; 8(12):357. https://doi.org/10.3390/toxins8120357
Chicago/Turabian StyleSteil, Daniel, Robert Bonse, Iris Meisen, Gottfried Pohlentz, German Vallejo, Helge Karch, and Johannes Müthing. 2016. "A Topographical Atlas of Shiga Toxin 2e Receptor Distribution in the Tissues of Weaned Piglets" Toxins 8, no. 12: 357. https://doi.org/10.3390/toxins8120357
APA StyleSteil, D., Bonse, R., Meisen, I., Pohlentz, G., Vallejo, G., Karch, H., & Müthing, J. (2016). A Topographical Atlas of Shiga Toxin 2e Receptor Distribution in the Tissues of Weaned Piglets. Toxins, 8(12), 357. https://doi.org/10.3390/toxins8120357