
New Insights into VacA Intoxication Mediated through Its Cell Surface Receptors

Abstract
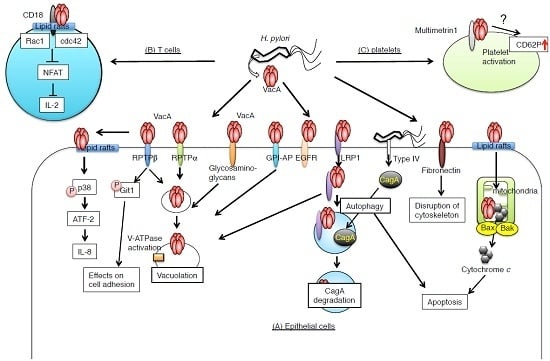
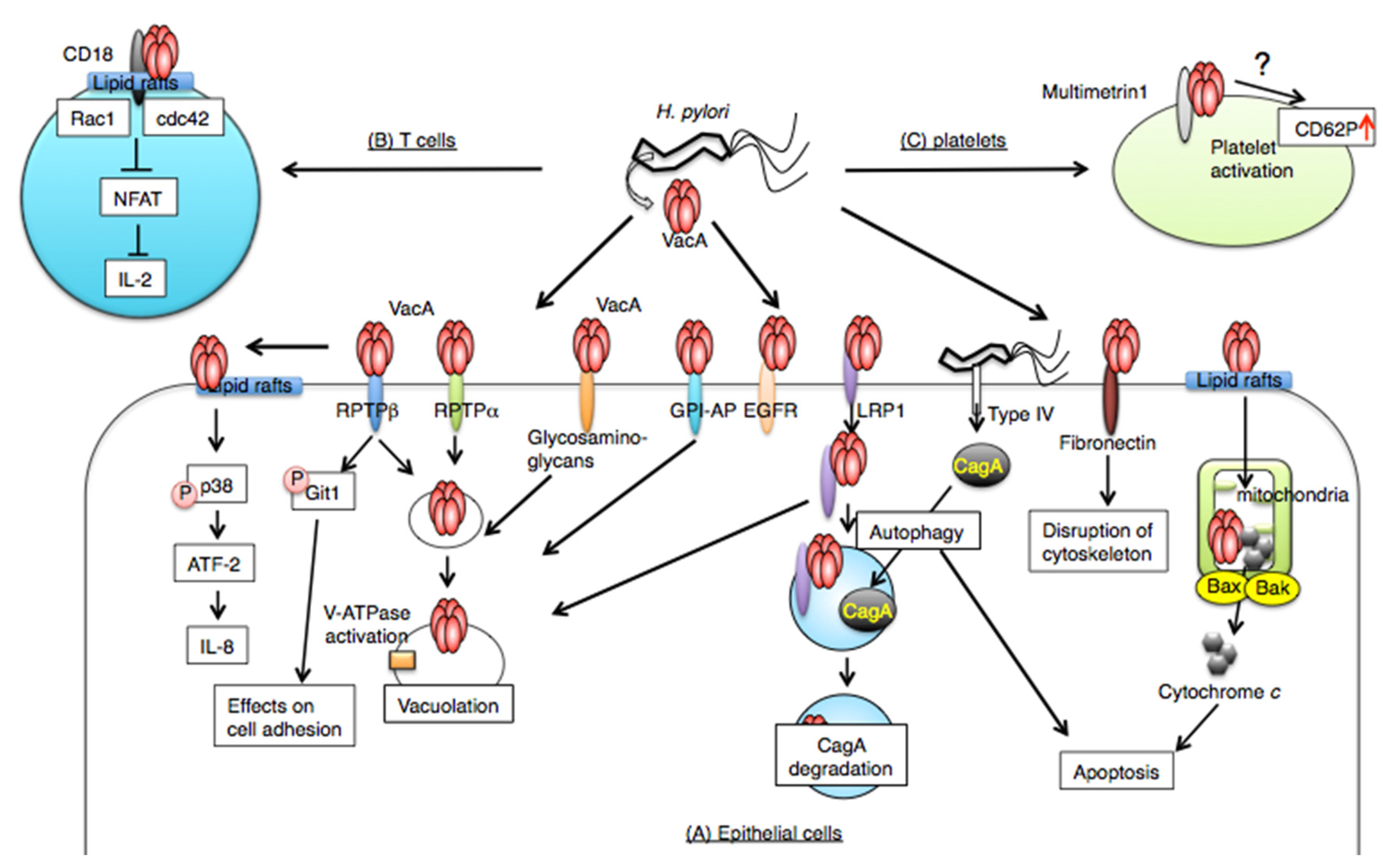
:
{kind=link}
{kind=link}
1. Introduction
2. VacA Receptors on Target Cells
2.1. Receptor-Like Protein Tyrosine Phosphatase β (RPTPβ)
2.2. Receptor-Like Protein Tyrosine Phosphatase α (RPTPα)
2.3. Low-Density Lipoprotein Receptor-Related Protein-1 (LRP1)
2.3.1. EGFR
2.3.2. Fibronectin
2.4. The Role of Lipid Rafts, Heparan Sulfate, and Glycosaminoglycans in VacA Binding
2.5. CD18 on T Lymphocytes
2.6. Multimerin 1 on Platelets
3. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Montecucco, C.; Rappuoli, R. Living dangerously: How Helicobacter pylori survives in the human stomach. Nat. Rev. Mol. Cell Biol. 2001, 2, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Matsushima, M.; Shirakura, K.; Koike, J.; Masui, A.; Takagi, A.; Shirasugi, Y.; Ogawa, Y.; Shirai, T.; Mine, T. Association of inflammatory cytokine gene polymorphisms with platelet recovery in idiopathic thrombocytopenic purpura patients after the eradication of Helicobacter pylori. Digestion 2008, 77, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Blaser, M.J.; Atherton, J.C. Helicobacter pylori persistence: Biology and disease. J. Clin. Investig. 2004, 113, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Algood, H.M.; Cover, T.L. Helicobacter pylori persistence: An overview of interactions between H. pylori and host immune defenses. Clin. Microbiol. Rev. 2006, 19, 597–613. [Google Scholar] [CrossRef] [PubMed]
- Suerbaum, S.; Michetti, P. Helicobacter pylori infection. N. Engl. J. Med. 2002, 347, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Pagliaccia, C.; de Bernard, M.; Lupetti, P.; Ji, X.; Burroni, D.; Cover, T.L.; Papini, E.; Rappuoli, R.; Telford, J.L.; Reyrat, J.M. The m2 form of the Helicobacter pylori cytotoxin has cell type-specific vacuolating activity. Proc. Natl. Acad. Sci. USA 1998, 95, 10212–10217. [Google Scholar] [CrossRef] [PubMed]
- Reyrat, J.M.; Lanzavecchia, S.; Lupetti, P.; de Bernard, M.; Pagliaccia, C.; Pelicic, V.; Charrel, M.; Ulivieri, C.; Norais, N.; Ji, X.; et al. 3D imaging of the 58 kDa cell binding subunit of the Helicobacter pylori cytotoxin. J. Mol. Biol. 1999, 290, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.C.; Wang, H.J.; Kuo, C.H. Two distinctive cell binding patterns by vacuolating toxin fused with glutathione S-transferase: One high-affinity m1-specific binding and the other lower-affinity binding for variant m forms. Biochemistry 2001, 40, 11887–11896. [Google Scholar] [CrossRef] [PubMed]
- De Bernard, M.; Burroni, D.; Papini, E.; Rappuoli, R.; Telford, J.; Montecucco, C. Identification of the Helicobacter pylori VacA toxin domain active in the cell cytosol. Infect. Immun. 1998, 66, 6014–6016. [Google Scholar] [PubMed]
- Ye, D.; Willhite, D.C.; Blanke, S.R. Identification of the minimal intracellular vacuolating domain of the Helicobacter pylori vacuolating toxin. J. Biol. Chem. 1999, 274, 9277–9282. [Google Scholar] [CrossRef] [PubMed]
- Ghose, C.; Perez-Perez, G.I.; Torres, V.J.; Crosatti, M.; Nomura, A.; Peek, R.M., Jr.; Cover, T.L.; Francois, F.; Blaser, M.J. Serological assays for identification of human gastric colonization by Helicobacter pylori strains expressing VacA m1 or m2. Clin. Vaccine Immunol. CVI 2007, 14, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Skibinski, D.A.; Genisset, C.; Barone, S.; Telford, J.L. The cell-specific phenotype of the polymorphic VacA midregion is independent of the appearance of the cell surface receptor protein tyrosine phosphatase beta. Infect. Immun. 2006, 74, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Gangwer, K.A.; Mushrush, D.J.; Stauff, D.L.; Spiller, B.; McClain, M.S.; Cover, T.L.; Lacy, D.B. Crystal structure of the Helicobacter pylori vacuolating toxin p55 domain. Proc. Natl. Acad. Sci. USA 2007, 104, 16293–16298. [Google Scholar] [CrossRef] [PubMed]
- De Guzman, B.B.; Hisatsune, J.; Nakayama, M.; Yahiro, K.; Wada, A.; Yamasaki, E.; Nishi, Y.; Yamazaki, S.; Azuma, T.; Ito, Y.; et al. Cytotoxicity and recognition of receptor-like protein tyrosine phosphatases, rptpalpha and rptpbeta, by Helicobacter pylori m2 VacA. Cell. Microbiol. 2005, 7, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Kuo, C.H.; Yeh, A.A.; Chang, P.C.; Wang, W.C. Vacuolating toxin production in clinical isolates of Helicobacter pylori with different VacA genotypes. J. Infect. Dis. 1998, 178, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Blaser, M.J. Purification and characterization of the vacuolating toxin from Helicobacter pylori. J. Biol. Chem. 1992, 267, 10570–10575. [Google Scholar] [PubMed]
- Cover, T.L.; Blanke, S.R. Helicobacter pylori VacA, a paradigm for toxin multifunctionality. Nat. Rev. Microbiol. 2005, 3, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Palframan, S.L.; Kwok, T.; Gabriel, K. Vacuolating cytotoxin A (VacA), a key toxin for Helicobacter pylori pathogenesis. Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Cid, T.P.; Fernandez, M.C.; Benito Martinez, S.; Jones, N.L. Pathogenesis of Helicobacter pylori infection. Helicobacter 2013, 18 (Suppl. 1), 12–17. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, L.K.; Jones, N.L. Modulation of autophagy by Helicobacter pylori and its role in gastric carcinogenesis. Trends Microbiol. 2013, 21, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Boquet, P.; Ricci, V. Intoxication strategy of Helicobacter pylori VacA toxin. Trends Microbiol. 2012, 20, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Rassow, J. Helicobacter pylori vacuolating toxin A and apoptosis. Cell Commun. Signal. CCS 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.J.; Blanke, S.R. Remodeling the host environment: Modulation of the gastric epithelium by the Helicobacter pylori vacuolating toxin (VacA). Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Terebiznik, M.R.; Raju, D.; Vazquez, C.L.; Torbricki, K.; Kulkarni, R.; Blanke, S.R.; Yoshimori, T.; Colombo, M.I.; Jones, N.L. Effect of Helicobacter pylori’s vacuolating cytotoxin on the autophagy pathway in gastric epithelial cells. Autophagy 2009, 5, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Telford, J.L.; Ghiara, P.; Dell’Orco, M.; Comanducci, M.; Burroni, D.; Bugnoli, M.; Tecce, M.F.; Censini, S.; Covacci, A.; Xiang, Z.; et al. Gene structure of the Helicobacter pylori cytotoxin and evidence of its key role in gastric disease. J. Exp. Med. 1994, 179, 1653–1658. [Google Scholar] [CrossRef] [PubMed]
- Tombola, F.; Carlesso, C.; Szabo, I.; de Bernard, M.; Reyrat, J.M.; Telford, J.L.; Rappuoli, R.; Montecucco, C.; Papini, E.; Zoratti, M. Helicobacter pylori vacuolating toxin forms anion-selective channels in planar lipid bilayers: Possible implications for the mechanism of cellular vacuolation. Biophys. J. 1999, 76, 1401–1409. [Google Scholar] [CrossRef]
- Szabo, I.; Brutsche, S.; Tombola, F.; Moschioni, M.; Satin, B.; Telford, J.L.; Rappuoli, R.; Montecucco, C.; Papini, E.; Zoratti, M. Formation of anion-selective channels in the cell plasma membrane by the toxin VacA of Helicobacter pylori is required for its biological activity. EMBO J. 1999, 18, 5517–5527. [Google Scholar] [CrossRef] [PubMed]
- Yahiro, K.; Satoh, M.; Nakano, M.; Hisatsune, J.; Isomoto, H.; Sap, J.; Suzuki, H.; Nomura, F.; Noda, M.; Moss, J.; et al. Low-density lipoprotein receptor-related protein-1 (LRP1) mediates autophagy and apoptosis caused by Helicobacter pylori VacA. J. Biol. Chem. 2012, 287, 31104–31115. [Google Scholar] [CrossRef] [PubMed]
- Yahiro, K.; Niidome, T.; Kimura, M.; Hatakeyama, T.; Aoyagi, H.; Kurazono, H.; Imagawa, K.; Wada, A.; Moss, J.; Hirayama, T. Activation of Helicobacter pylori VacA toxin by alkaline or acid conditions increases its binding to a 250-kDa receptor protein-tyrosine phosphatase beta. J. Biol. Chem. 1999, 274, 36693–36699. [Google Scholar] [CrossRef] [PubMed]
- Haunso, A.; Celio, M.R.; Margolis, R.K.; Menoud, P.A. Phosphacan immunoreactivity is associated with perineuronal nets around parvalbumin-expressing neurones. Brain Res. 1999, 834, 219–222. [Google Scholar] [CrossRef]
- Maeda, N.; Nishiwaki, T.; Shintani, T.; Hamanaka, H.; Noda, M. 6B4 proteoglycan/phosphacan, an extracellular variant of receptor-like protein-tyrosine phosphatase zeta/RPTPbeta, binds pleiotrophin/heparin-binding growth-associated molecule (HB-GAM). J. Biol. Chem. 1996, 271, 21446–21452. [Google Scholar] [PubMed]
- Meyer-Puttlitz, B.; Junker, E.; Margolis, R.U.; Margolis, R.K. Chondroitin sulfate proteoglycans in the developing central nervous system. II. Immunocytochemical localization of neurocan and phosphacan. J. Comp. Neurol. 1996, 366, 44–54. [Google Scholar] [CrossRef]
- Yuki, T.; Ishihara, S.; Rumi, M.; Ortega-Cava Cesar, F.; Kadowaki, Y.; Kazumori, H.; Yuki, M.; Wada, T.; Miyaoka, Y.; Yoshino, N.; et al. Expression of midkine and receptor-like protein tyrosine phosphatase (RPTP)-beta genes in the rat stomach and the influence of rebamipide. Aliment. Pharmacol. Ther. 2003, 18 (Suppl. 1), 106–112. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, A.; Shirasaka, D.; Yamamoto, S.; Ota, H.; Yahiro, K.; Fukada, M.; Shintani, T.; Wada, A.; Aoyama, N.; Hirayama, T.; et al. Mice deficient in protein tyrosine phosphatase receptor type Z are resistant to gastric ulcer induction by VacA of Helicobacter pylori. Nat. Genet. 2003, 33, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.N.; Davis, M.G.; Eby-Wilkens, E.; Howard, B.W.; Towne, B.A.; Dufresne, T.E.; Peters, K.G. Tyrosine phosphatase inhibition augments collateral blood flow in a rat model of peripheral vascular disease. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H268–H276. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, M.G.; Hughes, V.C.; Pan, L.; Simmons, M.; Daly, C.; Anderson, K.; Noguera-Troise, I.; Murphy, A.J.; Valenzuela, D.M.; Davis, S.; et al. Vascular endothelial tyrosine phosphatase (VE-PTP)-null mice undergo vasculogenesis but die embryonically because of defects in angiogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 3243–3248. [Google Scholar] [CrossRef] [PubMed]
- Baumer, S.; Keller, L.; Holtmann, A.; Funke, R.; August, B.; Gamp, A.; Wolburg, H.; Wolburg-Buchholz, K.; Deutsch, U.; Vestweber, D. Vascular endothelial cell-specific phosphotyrosine phosphatase (VE-PTP) activity is required for blood vessel development. Blood 2006, 107, 4754–4762. [Google Scholar] [CrossRef] [PubMed]
- De Bernard, M.; Moschioni, M.; Papini, E.; Telford, J.L.; Rappuoli, R.; Montecucco, C. Tpa and butyrate increase cell sensitivity to the vacuolating toxin of Helicobacter pylori. FEBS Lett. 1998, 436, 218–222. [Google Scholar] [CrossRef]
- Padilla, P.I.; Wada, A.; Yahiro, K.; Kimura, M.; Niidome, T.; Aoyagi, H.; Kumatori, A.; Anami, M.; Hayashi, T.; Fujisawa, J.; et al. Morphologic differentiation of HL-60 cells is associated with appearance of RPTPbeta and induction of Helicobacter pylori VacA sensitivity. J. Biol. Chem. 2000, 275, 15200–15206. [Google Scholar] [CrossRef] [PubMed]
- Yahiro, K.; Wada, A.; Yamasaki, E.; Nakayama, M.; Nishi, Y.; Hisatsune, J.; Morinaga, N.; Sap, J.; Noda, M.; Moss, J.; et al. Essential domain of receptor tyrosine phosphatase beta (RPTPbeta) for interaction with Helicobacter pylori vacuolating cytotoxin. J. Biol. Chem. 2004, 279, 51013–51021. [Google Scholar] [CrossRef] [PubMed]
- Hammer, A.; Oladimeji, P.; De Las Casas, L.E.; Diakonova, M. Phosphorylation of tyrosine 285 of PAK1 facilitates βPIX/GIT1 binding and adhesion turnover. FASEB J. 2015, 29, 943–959. [Google Scholar] [CrossRef] [PubMed]
- Matthews, R.J.; Cahir, E.D.; Thomas, M.L. Identification of an additional member of the protein-tyrosine-phosphatase family: Evidence for alternative splicing in the tyrosine phosphatase domain. Proc. Natl. Acad. Sci. USA 1990, 87, 4444–4448. [Google Scholar] [CrossRef] [PubMed]
- Sap, J.; D'Eustachio, P.; Givol, D.; Schlessinger, J. Cloning and expression of a widely expressed receptor tyrosine phosphatase. Proc. Natl. Acad. Sci. USA 1990, 87, 6112–6116. [Google Scholar] [CrossRef] [PubMed]
- Yahiro, K.; Wada, A.; Nakayama, M.; Kimura, T.; Ogushi, K.; Niidome, T.; Aoyagi, H.; Yoshino, K.; Yonezawa, K.; Moss, J.; et al. Protein-tyrosine phosphatase alpha, RPTP alpha, is a Helicobacter pylori VacA receptor. J. Biol. Chem. 2003, 278, 19183–19189. [Google Scholar] [CrossRef] [PubMed]
- Truffi, M.; Dubreuil, V.; Liang, X.; Vacaresse, N.; Nigon, F.; Han, S.P.; Yap, A.S.; Gomez, G.A.; Sap, J. RPTPalpha controls epithelial adherens junctions, linking E-cadherin engagement to c-Src-mediated phosphorylation of cortactin. J. Cell Sci. 2014, 127, 2420–2432. [Google Scholar] [CrossRef] [PubMed]
- Gomez, G.A.; McLachlan, R.W.; Wu, S.K.; Caldwell, B.J.; Moussa, E.; Verma, S.; Bastiani, M.; Priya, R.; Parton, R.G.; Gaus, K.; et al. An RPTPalpha/Src family kinase/RAP1 signaling module recruits myosin IIB to support contractile tension at apical E-cadherin junctions. Mol. Biol. Cell 2015, 26, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.M.; Strickland, D.K.; Bakillah, A. The mammalian low-density lipoprotein receptor family. Ann. Rev. Nutr. 1999, 19, 141–172. [Google Scholar] [CrossRef] [PubMed]
- Raju, D.; Hussey, S.; Ang, M.; Terebiznik, M.R.; Sibony, M.; Galindo-Mata, E.; Gupta, V.; Blanke, S.R.; Delgado, A.; Romero-Gallo, J.; et al. Vacuolating cytotoxin and variants in Atg16L1 that disrupt autophagy promote Helicobacter pylori infection in humans. Gastroenterology 2012, 142, 1160–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cover, T.L.; Krishna, U.S.; Israel, D.A.; Peek, R.M., Jr. Induction of gastric epithelial cell apoptosis by Helicobacter pylori vacuolating cytotoxin. Cancer Res. 2003, 63, 951–957. [Google Scholar] [PubMed]
- Willhite, D.C.; Blanke, S.R. Helicobacter pylori vacuolating cytotoxin enters cells, localizes to the mitochondria, and induces mitochondrial membrane permeability changes correlated to toxin channel activity. Cell. Microbiol. 2004, 6, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, E.; Wada, A.; Kumatori, A.; Nakagawa, I.; Funao, J.; Nakayama, M.; Hisatsune, J.; Kimura, M.; Moss, J.; Hirayama, T. Helicobacter pylori vacuolating cytotoxin induces activation of the proapoptotic proteins bax and bak, leading to cytochrome c release and cell death, independent of vacuolation. J. Biol. Chem. 2006, 281, 11250–11259. [Google Scholar] [CrossRef] [PubMed]
- Kuck, D.; Kolmerer, B.; Iking-Konert, C.; Krammer, P.H.; Stremmel, W.; Rudi, J. Vacuolating cytotoxin of Helicobacter pylori induces apoptosis in the human gastric epithelial cell line ags. Infect. Immun. 2001, 69, 5080–5087. [Google Scholar] [CrossRef] [PubMed]
- Galmiche, A.; Rassow, J.; Doye, A.; Cagnol, S.; Chambard, J.C.; Contamin, S.; de Thillot, V.; Just, I.; Ricci, V.; Solcia, E.; et al. The N-terminal 34 kDa fragment of Helicobacter pylori vacuolating cytotoxin targets mitochondria and induces cytochrome c release. EMBO J. 2000, 19, 6361–6370. [Google Scholar] [CrossRef] [PubMed]
- Tombola, F.; Oregna, F.; Brutsche, S.; Szabo, I.; Del Giudice, G.; Rappuoli, R.; Montecucco, C.; Papini, E.; Zoratti, M. Inhibition of the vacuolating and anion channel activities of the VacA toxin of Helicobacter pylori. FEBS Lett. 1999, 460, 221–225. [Google Scholar] [CrossRef]
- Jain, P.; Luo, Z.Q.; Blanke, S.R. Helicobacter pylori vacuolating cytotoxin A (VacA) engages the mitochondrial fission machinery to induce host cell death. Proc. Natl. Acad. Sci. USA 2011, 108, 16032–16037. [Google Scholar] [CrossRef] [PubMed]
- Radin, J.N.; Gonzalez-Rivera, C.; Ivie, S.E.; McClain, M.S.; Cover, T.L. Helicobacter pylori VacA induces programmed necrosis in gastric epithelial cells. Infect. Immun. 2011, 79, 2535–2543. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, M. Helicobacter pylori caga and gastric cancer: A paradigm for hit-and-run carcinogenesis. Cell Host Microbe 2014, 15, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Suzuki, H.; Saya, H.; Hatakeyama, M.; Hirayama, T.; Hirata, K.; Nagano, O.; Matsuzaki, J.; Hibi, T. Reactive oxygen species-induced autophagic degradation of Helicobacter pylori CagA is specifically suppressed in cancer stem-like cells. Cell Host Microbe 2012, 12, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Seto, K.; Hayashi-Kuwabara, Y.; Yoneta, T.; Suda, H.; Tamaki, H. Vacuolation induced by cytotoxin from Helicobacter pylori is mediated by the EGF receptor in hela cells. FEBS Lett. 1998, 431, 347–350. [Google Scholar] [CrossRef]
- Tabel, G.; Hoa, N.T.; Tarnawski, A.; Chen, J.; Domek, M.; Ma, T.Y. Helicobacter pylori infection inhibits healing of the wounded duodenal epithelium in vitro. J. Lab. Clin. Med. 2003, 142, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Tegtmeyer, N.; Zabler, D.; Schmidt, D.; Hartig, R.; Brandt, S.; Backert, S. Importance of EGF receptor, HER2/Neu and Erk1/2 kinase signalling for host cell elongation and scattering induced by the Helicobacter pylori caga protein: Antagonistic effects of the vacuolating cytotoxin VacA. Cell. Microbiol. 2009, 11, 488–505. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, M.J.; Smith, M.L. Multiscale relationships between fibronectin structure and functional properties. Acta Biomater. 2014, 10, 1524–1531. [Google Scholar] [PubMed]
- Schwarz-Linek, U.; Werner, J.M.; Pickford, A.R.; Gurusiddappa, S.; Kim, J.H.; Pilka, E.S.; Briggs, J.A.; Gough, T.S.; Hook, M.; Campbell, I.D.; et al. Pathogenic bacteria attach to human fibronectin through a tandem beta-zipper. Nature 2003, 423, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Gruenheid, S.; Finlay, B.B. Microbial pathogenesis and cytoskeletal function. Nature 2003, 422, 775–781. [Google Scholar] [PubMed]
- Hennig, E.E.; Godlewski, M.M.; Butruk, E.; Ostrowski, J. Helicobacter pylori VacA cytotoxin interacts with fibronectin and alters hela cell adhesion and cytoskeletal organization in vitro. FEMS Immunol. Med. Microbiol. 2005, 44, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Pai, R.; Cover, T.L.; Tarnawski, A.S. Helicobacter pylori vacuolating cytotoxin (VacA) disorganizes the cytoskeletal architecture of gastric epithelial cells. Biochem. Biophys. Res. Commun. 1999, 262, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Utt, M.; Danielsson, B.; Wadstrom, T. Helicobacter pylori vacuolating cytotoxin binding to a putative cell surface receptor, heparan sulfate, studied by surface plasmon resonance. FEMS Immunol. Med. Microbiol. 2001, 30, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Dedieu, S.; Langlois, B. Lrp-1: A new modulator of cytoskeleton dynamics and adhesive complex turnover in cancer cells. Cell Adhes. Migr. 2008, 2, 77–80. [Google Scholar] [CrossRef]
- Geisse, N.A.; Cover, T.L.; Henderson, R.M.; Edwardson, J.M. Targeting of Helicobacter pylori vacuolating toxin to lipid raft membrane domains analysed by atomic force microscopy. Biochem. J. 2004, 381, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Schraw, W.; Li, Y.; McClain, M.S.; van der Goot, F.G.; Cover, T.L. Association of Helicobacter pylori vacuolating toxin (VacA) with lipid rafts. J. Biol. Chem. 2002, 277, 34642–34650. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.K.; Willhite, D.C.; Patel, R.M.; Ye, D.; Williams, C.L.; Torres, E.M.; Marty, K.B.; MacDonald, R.A.; Blanke, S.R. Plasma membrane cholesterol modulates cellular vacuolation induced by the Helicobacter pylori vacuolating cytotoxin. Infect. Immun. 2002, 70, 4112–4123. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.H.; Wang, W.C. Binding and internalization of Helicobacter pylori VacA via cellular lipid rafts in epithelial cells. Biochem. Biophys. Res. Commun. 2003, 303, 640–644. [Google Scholar] [CrossRef]
- Nakayama, M.; Hisatsune, J.; Yamasaki, E.; Nishi, Y.; Wada, A.; Kurazono, H.; Sap, J.; Yahiro, K.; Moss, J.; Hirayama, T. Clustering of Helicobacter pylori VacA in lipid rafts, mediated by its receptor, receptor-like protein tyrosine phosphatase beta, is required for intoxication in AZ-521 cells. Infect. Immun. 2006, 74, 6571–6580. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.J. Lipid rafts: Bringing order to chaos. J. Lipid Res. 2003, 44, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Ricci, V.; Galmiche, A.; Doye, A.; Necchi, V.; Solcia, E.; Boquet, P. High cell sensitivity to Helicobacter pylori VacA toxin depends on a GPI-anchored protein and is not blocked by inhibition of the clathrin-mediated pathway of endocytosis. Mol. Biol. Cell 2000, 11, 3897–3909. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.R.; Patel, H.K.; Kostolansky, S.S.; Ballivian, R.A.; Eichberg, J.; Blanke, S.R. Sphingomyelin functions as a novel receptor for Helicobacter pylori VacA. PLoS Pathog. 2008, 4, e1000073. [Google Scholar] [CrossRef] [PubMed]
- Czajkowsky, D.M.; Iwamoto, H.; Cover, T.L.; Shao, Z. The vacuolating toxin from Helicobacter pylori forms hexameric pores in lipid bilayers at low ph. Proc. Natl. Acad. Sci. USA 1999, 96, 2001–2006. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.R.; Wilson, B.A.; Blanke, S.R. Sphingomyelin is important for the cellular entry and intracellular localization of Helicobacter pylori VacA. Cell. Microbiol. 2010, 12, 1517–1533. [Google Scholar] [CrossRef] [PubMed]
- Sasisekharan, R.; Venkataraman, G. Heparin and heparan sulfate: Biosynthesis, structure and function. Curr. Opin. Chem. Biol. 2000, 4, 626–631. [Google Scholar] [CrossRef]
- Sommi, P.; Ricci, V.; Romano, M.; Fiocca, R.; Ivey, K.J.; Solcia, E.; Ventura, U. H-pylori-induced cell vacuolation in vitro: Inhibitory action of heparin. Gastroenterology 1997, 112, A294–A294. [Google Scholar]
- Maeda, N.; Ichihara-Tanaka, K.; Kimura, T.; Kadomatsu, K.; Muramatsu, T.; Noda, M. A receptor-like protein-tyrosine phosphatase ptpzeta/rptpbeta binds a heparin-binding growth factor midkine. Involvement of arginine 78 of midkine in the high affinity binding to ptpzeta. J. Biol. Chem. 1999, 274, 12474–12479. [Google Scholar] [CrossRef] [PubMed]
- Molinari, M.; Salio, M.; Galli, C.; Norais, N.; Rappuoli, R.; Lanzavecchia, A.; Montecucco, C. Selective inhibition of ii-dependent antigen presentation by Helicobacter pylori toxin VacA. J. Exp. Med. 1998, 187, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Torres, V.J.; vanCompernolle, S.E.; Sundrud, M.S.; Unutmaz, D.; Cover, T.L. Helicobacter pylori vacuolating cytotoxin inhibits activation-induced proliferation of human T and B lymphocyte subsets. J. Immunol. 2007, 179, 5433–5440. [Google Scholar] [CrossRef] [PubMed]
- Gebert, B.; Fischer, W.; Weiss, E.; Hoffmann, R.; Haas, R. Helicobacter pylori vacuolating cytotoxin inhibits t lymphocyte activation. Science 2003, 301, 1099–1102. [Google Scholar] [CrossRef] [PubMed]
- Boncristiano, M.; Paccani, S.R.; Barone, S.; Ulivieri, C.; Patrussi, L.; Ilver, D.; Amedei, A.; D'Elios, M.M.; Telford, J.L.; Baldari, C.T. The Helicobacter pylori vacuolating toxin inhibits T cell activation by two independent mechanisms. J. Exp. Med. 2003, 198, 1887–1897. [Google Scholar] [CrossRef] [PubMed]
- Sundrud, M.S.; Torres, V.J.; Unutmaz, D.; Cover, T.L. Inhibition of primary human T cell proliferation by Helicobacter pylori vacuolating toxin (VacA) is independent of VacA effects on IL-2 secretion. Proc. Natl. Acad. Sci. USA 2004, 101, 7727–7732. [Google Scholar] [CrossRef] [PubMed]
- Sewald, X.; Gebert-Vogl, B.; Prassl, S.; Barwig, I.; Weiss, E.; Fabbri, M.; Osicka, R.; Schiemann, M.; Busch, D.H.; Semmrich, M.; et al. Integrin subunit CD18 is the T-lymphocyte receptor for the Helicobacter pylori vacuolating cytotoxin. Cell Host Microbe 2008, 3, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Sewald, X.; Jimenez-Soto, L.; Haas, R. Pkc-dependent endocytosis of the Helicobacter pylori vacuolating cytotoxin in primary T lymphocytes. Cell. Microbiol. 2011, 13, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rivera, C.; Algood, H.M.; Radin, J.N.; McClain, M.S.; Cover, T.L. The intermediate region of Helicobacter pylori VacA is a determinant of toxin potency in a jurkat T cell assay. Infect. Immun. 2012, 80, 2578–2588. [Google Scholar] [CrossRef] [PubMed]
- Rhead, J.L.; Letley, D.P.; Mohammadi, M.; Hussein, N.; Mohagheghi, M.A.; Eshagh Hosseini, M.; Atherton, J.C. A new Helicobacter pylori vacuolating cytotoxin determinant, the intermediate region, is associated with gastric cancer. Gastroenterology 2007, 133, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Kim, J.S.; Yoo, D.Y.; Ko, S.H.; Kim, N.; Kim, H.; Kim, Y.J. Stimulation of dendritic cells with Helicobacter pylori vacuolating cytotoxin negatively regulates their maturation via the restoration of E2F1. Clin. Exp. Immunol. 2011, 166, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Elizalde, J.I.; Gomez, J.; Panes, J.; Lozano, M.; Casadevall, M.; Ramirez, J.; Pizcueta, P.; Marco, F.; Rojas, F.D.; Granger, D.N.; et al. Platelet activation in mice and human Helicobacter pylori infection. J. Clin. Investig. 1997, 100, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Ahn, E.R.; Tiede, M.P.; Jy, W.; Bidot, C.J.; Fontana, V.; Ahn, Y.S. Platelet activation in Helicobacter pylori-associated idiopathic thrombocytopenic purpura: Eradication reduces platelet activation but seldom improves platelet counts. Acta Haematol. 2006, 116, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Massaguer, A.; Engel, P.; Perez-del-Pulgar, S.; Bosch, J.; Pizcueta, P. Production and characterization of monoclonal antibodies against conserved epitopes of P-selectin (CD62P). Tissue Antigens 2000, 56, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Satoh, K.; Hirayama, T.; Takano, K.; Suzuki-Inoue, K.; Sato, T.; Ohta, M.; Nakagomi, J.; Ozaki, Y. VacA, the vacuolating cytotoxin of Helicobacter pylori, binds to multimerin 1 on human platelets. Thromb. J. 2013, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Stanley, P. Golgi glycosylation. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lewandrowski, U.; Moebius, J.; Walter, U.; Sickmann, A. Elucidation of N-glycosylation sites on human platelet proteins: A glycoproteomic approach. Mol. Cell. Proteom. MCP 2006, 5, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Jeimy, S.B.; Fuller, N.; Tasneem, S.; Segers, K.; Stafford, A.R.; Weitz, J.I.; Camire, R.M.; Nicolaes, G.A.; Hayward, C.P. Multimerin 1 binds factor V and activated factor V with high affinity and inhibits thrombin generation. Thromb. Haemost. 2008, 100, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Tasneem, S.; Adam, F.; Minullina, I.; Pawlikowska, M.; Hui, S.K.; Zheng, S.; Miller, J.L.; Hayward, C.P. Platelet adhesion to multimerin 1 in vitro: Influences of platelet membrane receptors, von willebrand factor and shear. J. Thromb. Haemost. JTH 2009, 7, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, K.A. Microbial recognition of target-cell glycoconjugates. Curr. Opin. Struct. Biol. 1995, 5, 622–635. [Google Scholar] [CrossRef]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yahiro, K.; Hirayama, T.; Moss, J.; Noda, M. New Insights into VacA Intoxication Mediated through Its Cell Surface Receptors. Toxins 2016, 8, 152. https://doi.org/10.3390/toxins8050152
Yahiro K, Hirayama T, Moss J, Noda M. New Insights into VacA Intoxication Mediated through Its Cell Surface Receptors. Toxins. 2016; 8(5):152. https://doi.org/10.3390/toxins8050152
Chicago/Turabian StyleYahiro, Kinnosuke, Toshiya Hirayama, Joel Moss, and Masatoshi Noda. 2016. "New Insights into VacA Intoxication Mediated through Its Cell Surface Receptors" Toxins 8, no. 5: 152. https://doi.org/10.3390/toxins8050152
APA StyleYahiro, K., Hirayama, T., Moss, J., & Noda, M. (2016). New Insights into VacA Intoxication Mediated through Its Cell Surface Receptors. Toxins, 8(5), 152. https://doi.org/10.3390/toxins8050152