
Occurrence of Fusarium langsethiae and T-2 and HT-2 Toxins in Italian Malting Barley

 ,
,  , ,
, ,  ,
, 
Abstract
:
1. Introduction
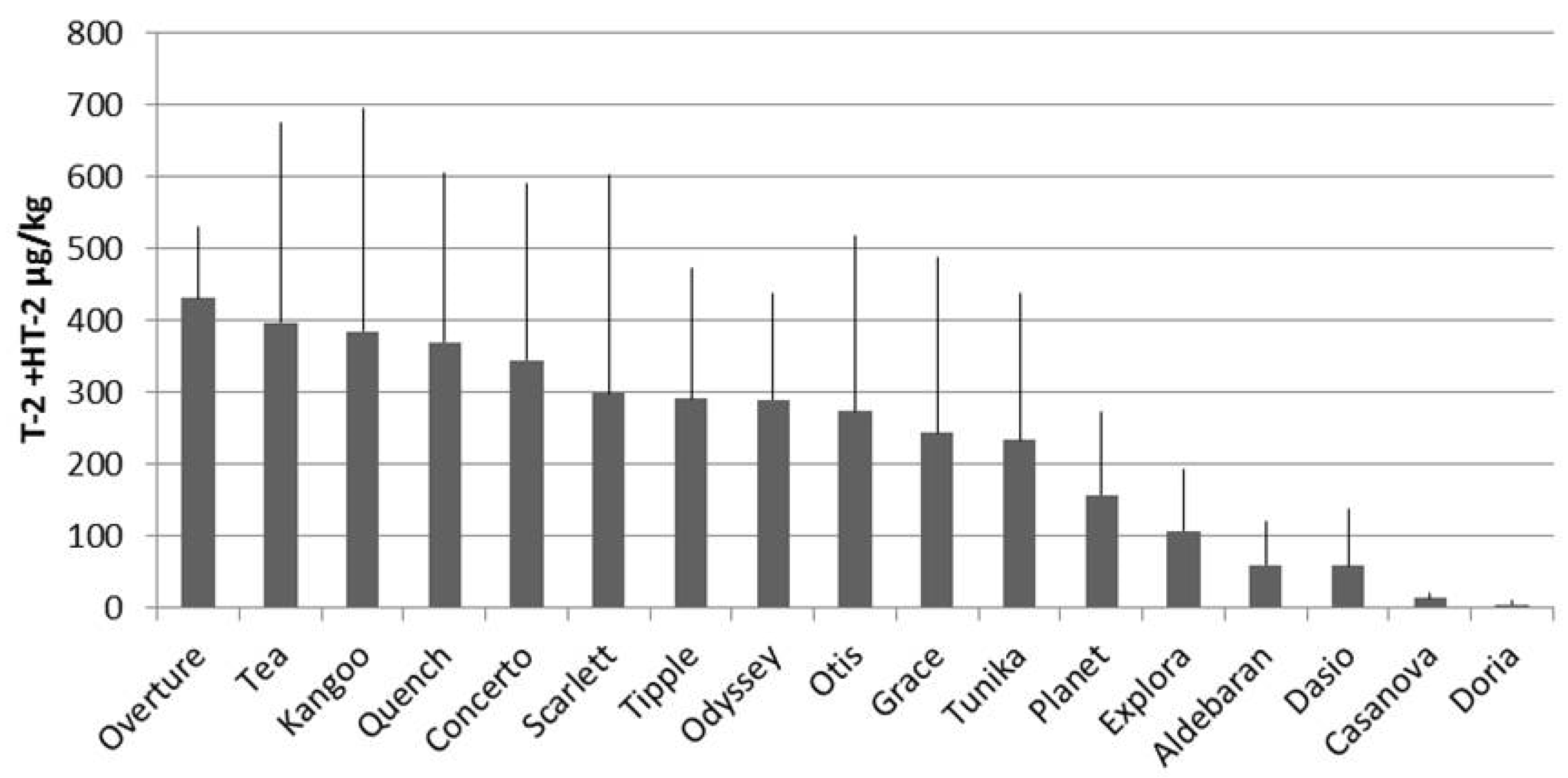
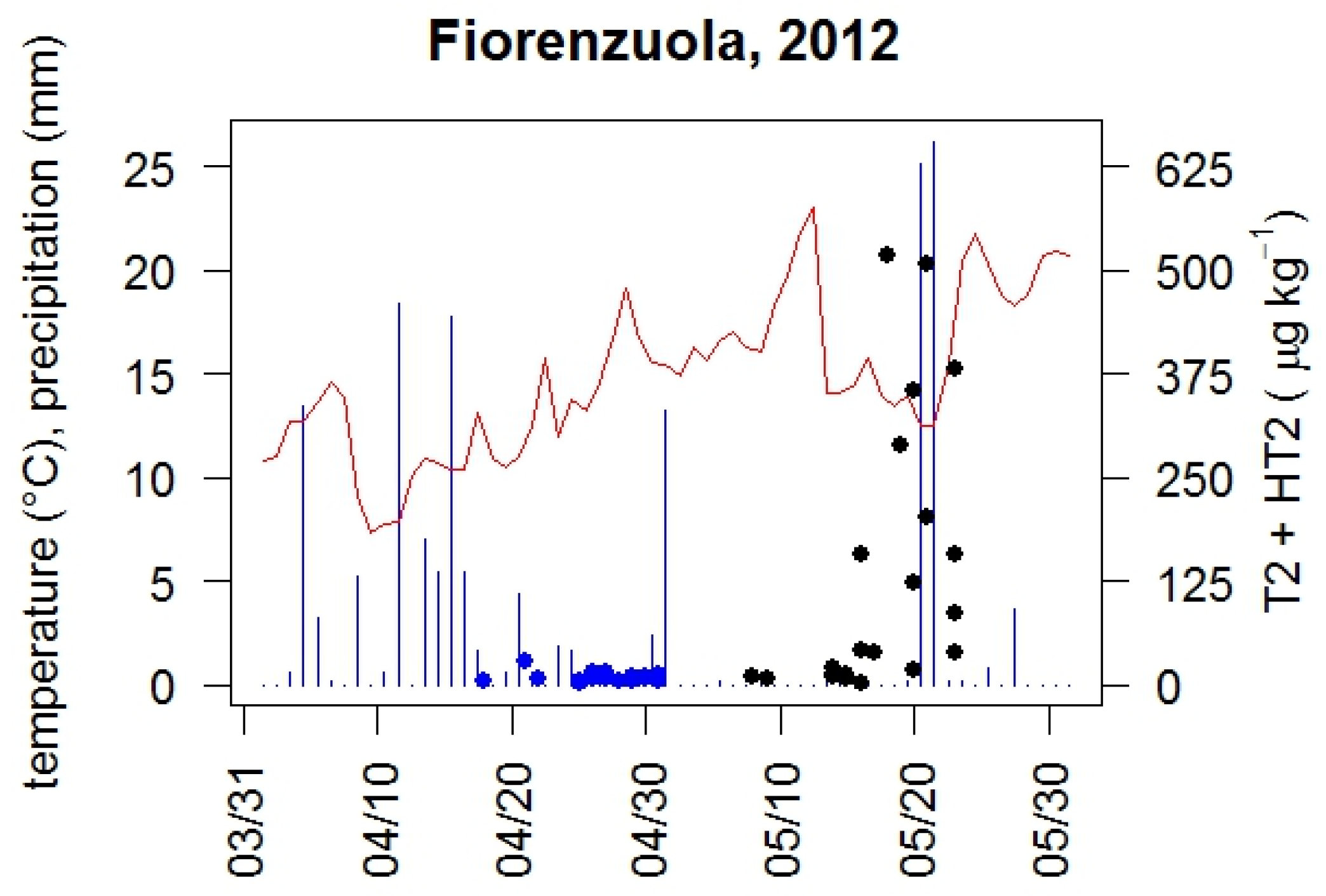
2. Results and Discussion
3. Conclusions
4. Material and Methods
4.1. Collection of Barley Samples
4.2. Fungal Strains
4.3. Mycotoxin Analysis
4.4. Determination and Quantification of Fusarium Species
4.4.1. DNA Extractions
4.4.2. Real-Time PCR
4.5. Meteorological Data
4.6. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stanca, A.M.; Gianinetti, A.; Rizza, F.; Terzi, V. Barley: An Overview of a Versatile Cereal Grain with Many Food and Feed Uses. In Encyclopedia of Food Grains, 2nd ed.; Wrigley, C., Corke, H., Seetharaman, K., Faubion, J., Eds.; Academic press: Waltham, MA, USA, 2016; pp. 147–152. [Google Scholar]
- Sarlin, T.; Nakari-Setälä, T.; Linder, M.; Penttilä, M.; Haikara, A. Fungal hydrophobins as predictors of the gushing activity of malt. J. Inst. Brew. 2005, 111, 105–111. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Bamforth, C.W. The microbiology of malting and brewing. MMBR 2013, 77, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Postulkova, M.; Riveros-Galan, D.; Cordova-Agiular, K.; Zitkova, K.; Verachtert, H.; Derdelinckx, G.; Dostalek, P.; Ruzicka, M.C.; Branyik, T. Technological possibilities to prevent and suppress primary gushing of beer. Trends Food Sci. Technol. 2016, 49, 64–73. [Google Scholar] [CrossRef]
- Terzi, V.; Tumino, G.; Stanca, A.M.; Morcia, C. Reducing the incidence of cereal head infection and mycotoxins in small grain cereal species. J. Cereal Sci. 2014, 59, 284–293. [Google Scholar] [CrossRef]
- Kirinčič, S.; Škrjanc, B.; Kos, N.; Kozolc, B.; Pirnat, N.; Tavčar-Kalcher, G. Mycotoxins in cereals and cereal products in Slovenia—Official control of foods in the years 2008–2012. Food Control 2015, 50, 157–165. [Google Scholar] [CrossRef]
- Pleadin, J.; Vahčić, N.; Perši, N.; Ševelj, D.; Markov, K.; Frece, J. Fusarium mycotoxins‘ occurrence in cereals harvested from Croatian fields. Food Control 2013, 32, 49–54. [Google Scholar] [CrossRef]
- Bĕláková, S.; Benešová, K.; Čáslavský, J.; Svoboda, Z. The occurrence of the selected Fusarium mycotoxins in Czech malting barley. Food Control 2014, 37, 93–98. [Google Scholar]
- Nielsen, L.K.; Cook, D.J.; Edwards, S.G.; Ray, R.V. The prevalence and impact of Fusarium head blight pathogens and mycotoxins on malting barley quality in UK. Int. J. Food Microbiol. 2014, 179, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Vea, M.; Lizarraga, E.; González-Peñas, E.; López de Cerain, A. Co-occurrence of type-A and type-B trichothecenes in barley from a northern region of Spain. Food Control 2012, 25, 81–88. [Google Scholar] [CrossRef]
- Infantino, A.; Santori, A.; Shah, D.A. Community structure of the Fusarium complex on wheat seed in Italy. Eur. J. Plant Pathol. 2012, 132, 499–510. [Google Scholar] [CrossRef]
- Infantino, A.; Santori, A.; Aureli, G.; Belocchi, A.; De Felice, S.; Tizzani, L.; Lattanzio, V.M.T.; Haidukowski, M.; Pascale, M. Occurrence of Fusarium langsethiae strains isolated from Durum Wheat in Italy. J. Phytophatol. 2015, 163, 612–619. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Hohn, T.M.; McCormick, S.P. Trichothecene biosynthesis in Fusarium. Microbiol. Mol. Biol. Rev. 1993, 57, 595–604. [Google Scholar]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Slaiding, I. T-2, HT-2 and deoxynivalenol (DON) in malting barley and malt. In Proceedings of the 6th EC Fusarium Toxin Forum, Brussels, Belgium, 9–10 February 2009.
- Occurrence Data of Trichothecene Mycotoxins T-2 Toxin and HAT-2 Toxin in Food and Feed. Available online: http://onlinelibrary.wiley.com/doi/10.2903/sp.efsa.2010.EN-66/abstract (accessed on 19 August 2016).
- On the Presence of T-2 Toxin and HT-2 Toxin in Cereals and Cereal Products. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32013H0165 (accessed on 19 August 2016).
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional barley. FAD Part B 2009, 26, 1185–1190. [Google Scholar]
- Hietaniemi, V.; Rämö, S.; Yli-Mattila, T.; Jestoi, M.; Peltonen, S.; Kartio, M.; Sieviläinen, E.; Koivisto, T.; Parikka, P. Updated survey of Fusarium species and toxins in Finnish cereal grains. FAD Part A 2016. [Google Scholar] [CrossRef]
- Opoku, N.; Back, M.; Edwards, S.G. Development of Fusarium langsethiae in commercial cereal production. Eur. J. Plant Pathol. 2013, 136, 159–170. [Google Scholar] [CrossRef]
- Linkmeyer, A.; Hofer, K.; Rychlik, M.; Herz, M.; Hausladen, H.; Hückelhoven, R.; Hess, M. Influence of inoculum and climatic factors on the severity of Fusarium head blight in German spring and winter barley. FAD Part A 2016, 33, 489–499. [Google Scholar]
- Rossi, V.; Languasco, L.; Pattori, E.; Giosuè, S. Dynamics of airborne Fusarium macroconidia in wheat fields naturally affected by Head Blight. J. Plant Pathol. 2002, 84, 53–64. [Google Scholar]
- Orlando, B.; Barrier-Guillot, B.; Gourdain, E.; Maumene, C. Identification of agronomic factors that influence the levels of T-2 and HT-2 toxins in barley grown in France. World Mycotox. J. 2010, 3, 169–174. [Google Scholar] [CrossRef]
- Mankevičienė, A.; Butkutė, B.; Gaurilčikienė, I.; Dabkevičius, Z. Risk assessment of Fusarium mycotoxins in Lithuanian small grain cereal grains. Food Control 2011, 22, 970–976. [Google Scholar] [CrossRef]
- Yli-Mattila, T.; Paavanen-Huhtala, S.; Parikka, P.; Hietaniemi, V.; Jestoi, M.; Rizzo, A. Toxigenic fungi and mycotoxins in Finnish cereals. In An Overview on Toxigenic Fungi and Mycotoxins in Europe; Logrieco, A., Visconti, A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 83–100. [Google Scholar]
- Yli-Mattila, T.; Paavanen-Huhtala, S.; Jestoi, M.; Parikka, P.; Hietaniemi, V.; Gagkaeva, T.; Sarlin, T.; Haikara, A.; Laaksonen, S.; Rizzo, A. Real-time PCR detection and quantification of Fusarium poae, F. graminearum, F. sporotichioides and F. langsethiae in cereal grains in Finland and Russia. Arch. Phytopathol. Plant Prot. 2008, 41, 243–260. [Google Scholar] [CrossRef]
- Yli-Mattila, T.; Parikka, P.; Lahtinen, T.; Ramo, S.; Kokkonen, M.; Rizzo, A.; Jestoi, M.; Hietaniemi, V. Fusarium DNA levels in Finnish cereal grains. In Current Advances in Molecular Mycology; Gherbawy, Y., Mach, L., Rai, M., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2009; pp. 107–138. [Google Scholar]
- Fredlund, E.; Gidlund, A.; Pettersson, H.; Olsen, M.; Börjesson, T. Real-time PCR detection of Fusarium species in Swedish oats and correlation to T-2 and HT-2 toxin content. World Mycot. J. 2010, 3, 77–88. [Google Scholar] [CrossRef]
- Wilson, A.; Simpson, D.; Chandler, E.; Jennings, P.; Nicholson, P. Development of PCR assays for the detection and differentiation of Fusarium sporotrichioides and Fusarium langsethiae. FEMS Microbiol. Lett. 2004, 233, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.G.; Imathiu, S.M.; Ray, R.V.; Back, M.; Hare, M.C. Molecular studies to identify the Fusarium species responsible for HT-2 and T-2 mycotoxins in UK oats. Int. J. Food Microbiol. 2012, 156, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Jestoi, M.; Rokka, M.; Yli-Mattila, T.; Parikka, P.; Rizzo, A.; Peltonen, K. Presence and concentrations of the Fusarium-related mycotoxins beauvericin, enniatins and moniliformin in Finnish grain samples. Food Addit. Contam. 2004, 21, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Jestoi, M.N.; Paavanen-Huhtala, S.; Parikka, P.; Yli-Mattila, T. In vitro and in vivo mycotoxin production of Fusarium species isolated from Finnish grains. Arch. Phytopathol. Plant Prot. 2008, 41, 545–558. [Google Scholar] [CrossRef]
- Kokkonen, M.; Ojala, L.; Parikka, P.; Jestoi, M. Mycotoxin production of selected Fusarium species at different culture conditions. Int. J. Food Microbiol. 2010, 143, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Vogelgsang, S.; Sulyok, M.; Banziger, I.; Krska, R.; Schuhmacher, R.; Forrer, H.R. Effect of fungal strain and cereal substrate on in vitro mycotoxin production by Fusarium poae and Fusarium avenaceum. FAD 2008, 25, 745–757. [Google Scholar]
- Infantino, A.; Pucci, N.; Conca, G.; Santori, A. First report of Fusarium langsethiae on durum wheat kernels in Italy. Plant Dis. 2007, 91, 1362–1362. [Google Scholar] [CrossRef]
- Hofgaard, I.S.; Aamot, H.U.; Torp, T.; Jestoi, M.; Lattanzio, V.M.T.; Klemsdal, S.S.; Waalwijk, C.; Van der Lee, T.; Brodal, G. Associations between Fusarium species and mycotoxins in oats and spring wheat from farmers’ fields in Norway over a six-year period. World Mycot. J. 2016, 9, 365–378. [Google Scholar] [CrossRef]
- Torp, M.; Langseth, W. Production of T-2 toxin by a Fusarium resembling Fusarium poae. Mycopathologia 1999, 147, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Torp, M.; Nirenberg, H.I. Fusarium langsethiae sp. nov. on cereals in Europe. Int. J. Food Microbiol. 2004, 95, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Lysøe, E.; Frandsen, R.J.; Divon, H.H.; Terzi, V.; Orrù, L.; Lamontanara, A.; Kolseth, A.K.; Nielsen, K.F.; Thrane, U. Draft genome sequence and chemical profiling of Fusarium langsethiae, an emerging producer of type A trichothecenes. Int. J. Food Microbiol. 2016, 221, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torp, M.; Adler, A. The European Sporotrichiella project: A polyphasic approach to the biology of a new Fusarium species. Int. J. Food Microbiol. 2004, 95, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Divon, H.H.; Razzaghian, J.; Udnes-Aamot, H.; Klemsdal, S.S. Fusarium langsethiae (Torp and Nirenberg), investigation of alternative infection routes in oats. Eur. J. Plant Pathol. 2011, 132, 147–161. [Google Scholar] [CrossRef]
- Nazari, L.; Pattori, E.; Terzi, V.; Morcia, C.; Rossi, V. Influence of temperature on infection, growth, and mycotoxin production by Fusarium langsethiae and F. sporotrichioides in durum wheat. Food Microbiol. 2014, 39, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Imathiu, S.M.; Edwards, S.G.; Ray, R.V.; Back, M.A. Fusarium langsethiae—A HT-2 and T-2 toxins producer that needs more attention. J. Phytopathol. 2013, 161, 1–10. [Google Scholar] [CrossRef]
- Imathiu, S.M.; Ray, R.V.; Back, M.I.; Hare, M.C.; Edwards, S.G. A survey investigating the infection of Fusarium langsethiae and production of HT-2 and T-2 mycotoxins in UK oat fields. J. Phytopathol. 2013, 161, 1–10. [Google Scholar] [CrossRef]
- Rocha, O.; Ansari, K.; Doohan, F.M. Effects of trichothecene mycotoxins on eukaryotic cells: A review. FAD 2005, 22, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Escrivá, L.; Font, G.; Manyes, L. In vivo toxicity studies of Fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef] [PubMed]
- De Ruyck, K.; De Boevre, M.; Huybrechts, I.; De Saeger, S. Dietary mycotoxins, co-exposure, and carcinogenesis in humans: Short review. Mut. Res. Rev. Mut. Res. 2015, 766, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA Panel on Contaminants in the Food Chain. Scientific opinion on the risk for animal and public health related to the presence of T-2 and HAT-2 toxin in food and feed. EFSA J. 2011, 9, 2481. [Google Scholar] [CrossRef]
- Ueno, Y. The toxicology of mycotoxins. CRC Crit. Rev. Toxicol. 1985, 14, 99–132. [Google Scholar] [CrossRef] [PubMed]
- Kokkonen, M.; Medina, A.; Magan, N. Comparative study of water and temperature relations of growth and T-2/HT-2 toxin production by strains of Fusarium sporotrichioides and Fusarium langsethiae. World Mycotoxin J. 2012, 5, 365–372. [Google Scholar] [CrossRef]
- Strub, C.; Pocaznoi, D.; Lebrihi, A.; Fournier, R.; Mathieu, F. Influence of barley malting operating parameters on T-2 and HT-2 toxinogenesis of Fusarium langsethiae, a worring contaminant of malting barley in Europe. FAD Part A 2010, 27, 1247–1252. [Google Scholar]
- Hofgaard, I.S.; Seehusen, T.; Aamot, H.U.; Riley, H.; Razzaghian, J.; Le, V.H.; Hjelkrem, A.-G.R.; Dill-Macky, R.; Brodal, G. Inoculum Potential of Fusarium spp. Relates to Tillage and Straw Management in Norwegian Fields of Spring Oats. Front. Microbiol. 2016, 7, 556. [Google Scholar] [CrossRef] [PubMed]
- Bernhoft, A.; Clasen, P.-E.; Kristoffersen, A.B.; Torp, M. Less Fusarium infestation and mycotoxin contamination in organic than in conventional cereals. Food Addit. Contam. 2010, 27, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Malachova, A.; Cerkal, R.; Ehrenbergerova, J.; Dzuman, Z.; Vaculova, K.; Hajslova, J. Fusarium mycotoxins in various barley cultivars and their transfer into malt. J. Sci. Food Agric. 2010, 90, 2495–2505. [Google Scholar] [CrossRef] [PubMed]
- Mankevičienė, A.; Gaurilčikienė, I.; Dabkevičius, Z.; Semaškienė, R.; Mačkinaitė, R.; Supronienė, S. Mycotoxin contamination of Lithuanian-grown cereal grains and factors determining it. Ekologija 2006, 3, 21–27. [Google Scholar]
- Barreau, C. Mycotoxins in French cereal and regulation of trichothecene biosynthesis in grain by Fusarium. In Proceedings of the 2nd Mediterranean MycoRed Workshop, Istanbul, Turkey, 14 October 2011.
- Brera, C.; De Santis, B.; Prantera, E.; Debegnach, F.; Pannunzi, E.; Fasano, F.; Berdini, C.; Slate, A.B.; Miraglia, M.; Whitaker, T.B. Effect of sample size in the evaluation of “in field” sampling plans for Aflatoxin B1 determination in corn. J. Agric. Food Chem. 2010, 58, 8481–8489. [Google Scholar] [CrossRef] [PubMed]
- Morcia, C.; Malnati, M.; Terzi, V. In vitro antifungal activity of terpinen-4-ol, eugenol, carvone, 1,8-cineole (eucalyptol) and thymol against mycotoxigenic plant pathogens. Food Addit. Contam. 2012, 29, 415–422. [Google Scholar]
- Rossi, V.; Terzi, V.; Moggi, F.; Morcia, C.; Faccioli, P.; Haidukowski, M.; Pascale, M. Assessment of Fusarium infection in wheat heads using a quantitative polymerase chain reaction (qPCR) assay. FAD 2007, 24, 1121–1130. [Google Scholar]
- Terzi, V.; Morcia, C.; Faccioli, P.; Faccini, N.; Rossi, V.; Cigolini, M.; Corbellini, M.; Scudellari, D.; Delogu, G. Fusarium DNA traceability along the bread production chain. Int. J. Food Sci. Technol. 2007, 42, 1390–1396. [Google Scholar] [CrossRef]
- Studies on Fusarium poæ, F. sporotrichioides and F. langsethiæ, Responsible for Production of T2/HT2 and Nivalenol in Wheat. Available online: http://hdl.handle.net/10280/1748 (accessed on 19 August 2016). (In Italian)
- Brera, C.; De Santis, B.; Trifirò, G.; Debegnach, F. Studio interlaboratorio per la validazione di un metodo immunoenzimatico per la determinazione delle tossine T-2 e HT-2 in campioni di frumento. In Atti Progetto Micoprincem Micotossine Principali ed Emergenti nei Cereali; Consiglio per la Ricerca e la Sperimentazione in Agricoltura: Rome, Italy, 2014; Volume 1, pp. 8–13. (In Italian) [Google Scholar]
- Busman, M.; Poling, S.M.; Maragos, C.M. Observation of T-2 Toxin and HT-2 Toxin Glucosides from Fusarium sporotrichioides by Liquid Chromatography Coupled to Tandem Mass Spectrometry (LC-MS/MS). Toxins 2011, 3, 1554–1568. [Google Scholar] [CrossRef] [PubMed]
- Sarlin, T.; Yli-Mattila, T.; Jestoi, M.; Rizzo, A.; Paavanen-Huhtala, S.; Haikara, A. Real-time PCR for quantification of toxigenic Fusarium species in barley and malt. Eur. J. Plant Pathol. 2006, 114, 371–380. [Google Scholar] [CrossRef]
- Al-Samarrai, T.H.; Schmid, J. A simple method for extraction of fungal genomic DNA. Lett. Appl. Microbiol. 2000, 30, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Faccioli, P.; Ciceri, G.P.; Provero, P.; Stanca, A.M.; Morcia, C.; Terzi, V.A. combined strategy of “in silico” transcriptome analysis and web search engine optimization allows an agile identification of reference genes suitable for normalization in gene expression studies. Plant Mol. Biol. 2007, 63, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Nicolaisen, M.; Suproniene, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Iustesen, A.F. Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Methods 2009, 76, 234–240. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growing Season | Number of Analyzed Samples | Positive Samples a | ||||
|---|---|---|---|---|---|---|
| % | Mean (µg/kg) | Median (µg/kg) | Range (µg/kg) | % Positives > 200 µg/kg b | ||
| 2011 | 72 | 28 | 69 | 54 | 30–467 | 2.7 |
| 2012 | 303 | 22 | 72 | 34 | 26–518 | 2 |
| 2013 | 158 | 23 | 264 | 212 | 26–787 | 11.4 |
| 2014 | 158 | 53 | 106 | 27 | 26–724 | 19.6 |
| Growing Season | Number of Analyzed Samples | Positive Samples a | |||
|---|---|---|---|---|---|
| % | Mean | Median | Range | ||
| 2011 | 28 | 100 | 95 | 66 | 37–467 |
| 2012 | 48 | 59 | 70 | 40 | 39–518 |
| 2013 | 18 | 94 | 358 | 285 | 90–787 |
| 2014 | 13 | 92 | 271 | 276 | 76–466 |
| Growing Season | Number of Analyzed Samples | Positive Samples a | |||
|---|---|---|---|---|---|
| % | Mean | Median | Range | ||
| 2012 | 13 | 0 | - | - | - |
| 2013, autumn sowing | 11 | 27 | 25 | 22 | 29–64 |
| 2013, spring sowing | 9 | 73 | 312 | 296 | 57–877 |
| 2014, autumn sowing | 15 | 71 | 30 | 29 | 26–36 |
| 2014, spring sowing | 13 | 70 | 109 | 76 | 43–391 |
| Toxin | Mean Value (μg/kg) | Max Value (μg/kg) | Country | Period | Reference |
|---|---|---|---|---|---|
| T-2 + HT-2 | 36 | 277 | Finland | 2005–2006 | [19] |
| T-2 + HT-2 | 37 | 626 | Finland | 2005–2014 | [19] |
| T-2 + HT-2 | ≤20 | - | Norway | 2002–2004 | [54] |
| T-2 | 30 | - | Czech Republic | 2005–2008 | [55] |
| HT-2 | 110 | 716 | Czech Republic | 2005–2008 | [55] |
| T-2 + HT-2 | 8.9 | 145 | Czech Republic | 2008–2011 | [8] |
| T-2 + HT-2 | 29.2 (in positive samples) | 532.5 | Spain | 2007 | [10] |
| T-2 +HT-2 | ≤20 | 138 | UK | 2002–2005 | [18] |
| T-2 | 19.7 | 319 | Lithuania | 2003–2005 | [56] |
| T-2 + HT-2 | 45.72 | 755.7 | France (spring barley) | 2007–2010 | [57] |
| T-2 + HT-2 | 10.4 | 340.1 | France (winter barley) | 2007–2010 | [57] |
| T-2 + HT-2 | 127 (in positive samples) | 787 | Italy | 2011–2014 | this work |
| Species | Strain | Source of Isolation | Obtained from | Repository | References |
|---|---|---|---|---|---|
| F. sporotrichioides | ITEM 194 | Zea mays kernel (Italy) | Institute of Sciences of Food Production, CNR, Bari, Italy | Institute of Sciences of Food Production, CNR, Bari, Italy | [59] |
| F. langsethiae | 11020 | Durum wheat kernels (Italy) | Università del Sacro Cuore, Di.Pro.Ve.S., Piacenza, Italy | Università del Sacro Cuore, Di.Pro.Ve.S, Piacenza, Italy | [42] |
| F. graminearum | ITEM 6477 | Triticum sp. (Italy) | Institute of Sciences of Food Production, CNR, Bari, Italy | Institute of Sciences of Food Production, CNR, Bari, Italy | [60] |
| F. culmorum | MPVP 70 | Bread wheat plants (Italy) | Università del Sacro Cuore, Di.Pro.Ve.S, Piacenza, Italy | Università del Sacro Cuore, Di.Pro.Ve.S, Piacenza, Italy | [61] |
| F. poae | ITEM 10402 | Wheat kernels (Italy) | Università del Sacro Cuore, Di.Pro.Ve.S, Piacenza, Italy | Institute of Sciences of Food Production, CNR, Bari, Italy | [62] |
| Primer Names and Sequences | Use | Reference |
|---|---|---|
| TC130707F (TCGGCTACAGCATTGAAGACG) | Barley DNA quantification | [67] |
| TC130707R (CCAAAAACGATATCAGGATGGC) | ||
| F.langA29F (CAAGTCGACCACTGTGAGTACCTCT) | F. langsethiae DNA quantification | [68] |
| F.langA95R (TGTCAAAGCATGTCAGTAAAGATGAC) | ||
| F.spoA18F (CGAAGTCGACCACTGTGAGTACA) | F. sporotrichioides DNA quantification | [68] |
| F.spoA18R (CTGTCAAAGCATGTCACTAAAAATGAT) | ||
| F.poaeA51F (ACCGAATCTCAACTCCGCTTT) | F. poae DNA quantification | [68] |
| F.poaeA98R (GTCTGTCAAGCATGTTAGCACAAGT) | ||
| 22F (AATATGGAAAACGGAGTTCATCTACA) | F. graminearum and F. culmorum DNA quantification | [68] |
| 122 R (ATTGCCGGTGCCTGAAAGT) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morcia, C.; Tumino, G.; Ghizzoni, R.; Badeck, F.W.; Lattanzio, V.M.T.; Pascale, M.; Terzi, V. Occurrence of Fusarium langsethiae and T-2 and HT-2 Toxins in Italian Malting Barley. Toxins 2016, 8, 247. https://doi.org/10.3390/toxins8080247
Morcia C, Tumino G, Ghizzoni R, Badeck FW, Lattanzio VMT, Pascale M, Terzi V. Occurrence of Fusarium langsethiae and T-2 and HT-2 Toxins in Italian Malting Barley. Toxins. 2016; 8(8):247. https://doi.org/10.3390/toxins8080247
Chicago/Turabian StyleMorcia, Caterina, Giorgio Tumino, Roberta Ghizzoni, Franz W. Badeck, Veronica M.T. Lattanzio, Michelangelo Pascale, and Valeria Terzi. 2016. "Occurrence of Fusarium langsethiae and T-2 and HT-2 Toxins in Italian Malting Barley" Toxins 8, no. 8: 247. https://doi.org/10.3390/toxins8080247
APA StyleMorcia, C., Tumino, G., Ghizzoni, R., Badeck, F. W., Lattanzio, V. M. T., Pascale, M., & Terzi, V. (2016). Occurrence of Fusarium langsethiae and T-2 and HT-2 Toxins in Italian Malting Barley. Toxins, 8(8), 247. https://doi.org/10.3390/toxins8080247