
Comparative Study of Biological Activities of Venom from Colubrid Snakes Rhabdophis tigrinus (Yamakagashi) and Rhabdophis lateralis


 ,
,
Abstract
:1. Introduction
2. Results and Discussion
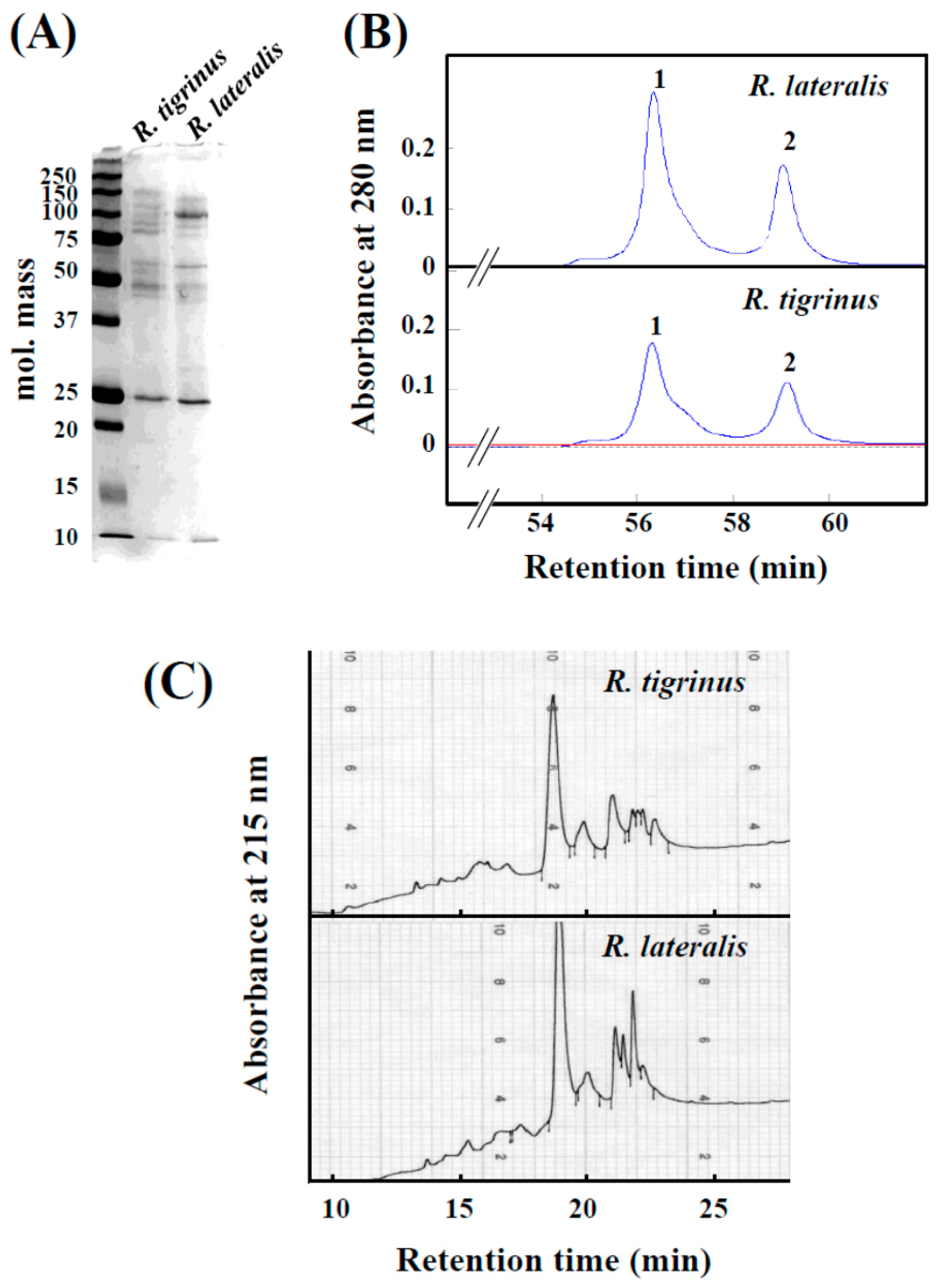
2.1. Comparison of the Venom Components
2.2. Effect on the Blood Coagulation System
2.2.1. Clotting Time of Human Plasma
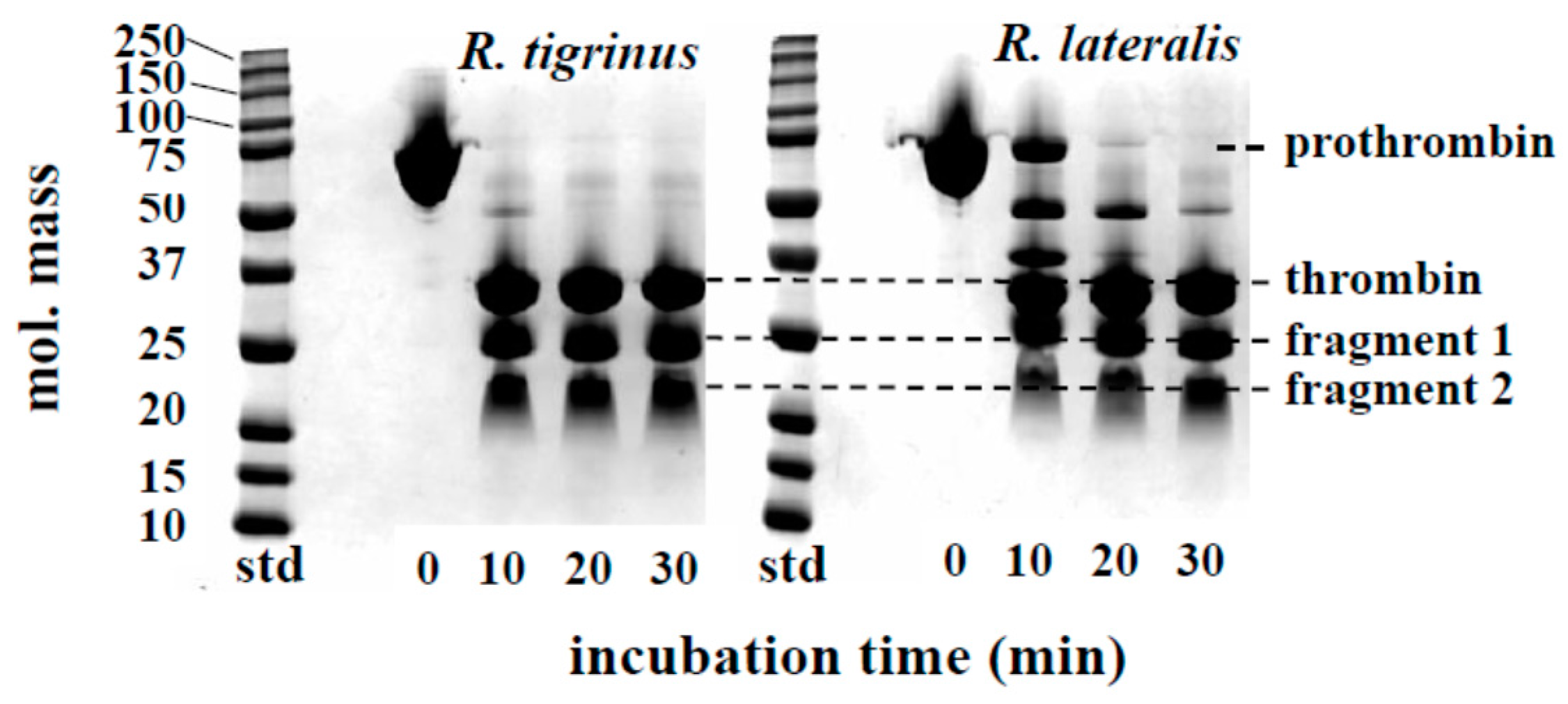
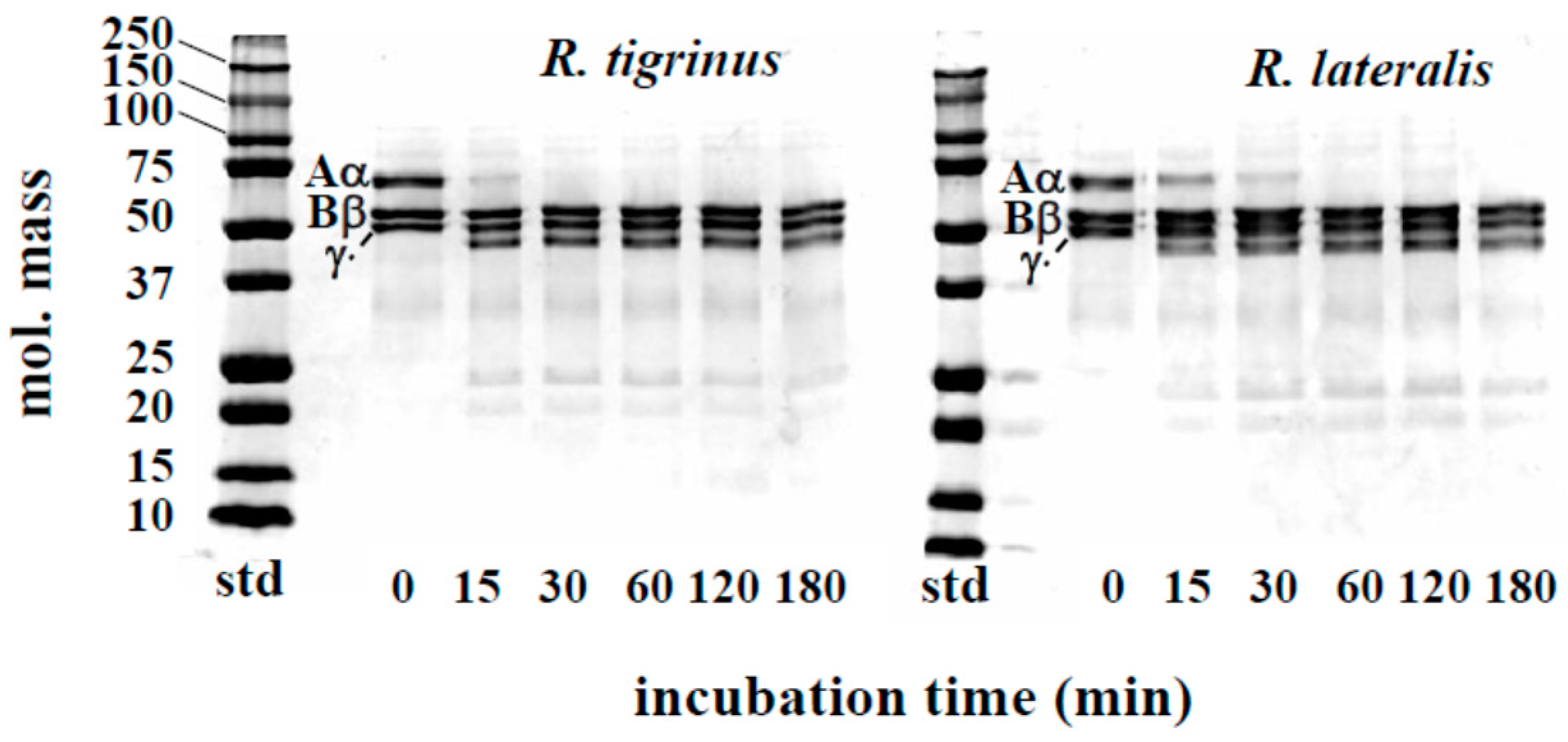
2.2.2. Effect of Venom on Degradation of Prothrombin and Fibrinogen
2.3. Proteolytic Activity
2.3.1. Effects of Venom Fractions on Synthetic Substrates
2.3.2. Effects of Venom Fractions on Extracellular Matrix Proteins
2.3.3. Comparison of Proteolytic Specificity
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Fractionation of Crude Venom
4.3. Blood Coagulation Assays
4.4. Proteolytic Activities
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Takeuchi, H.; Ota, H.; Oh, H.-S.; Hikida, T. Extensive genetic divergence in the East Asian natricine snake, Rhabdophis tigrinus (Serpentes: Colubridae), with special reference to prominent geographical differentiation of the mitochondrial cytochrome b gene in Japanese populations. Biol. J. Linn. Soc. Lond. 2012, 105, 395–408. [Google Scholar] [CrossRef]
- Uetz, P.; Hallermann, J. The Reptile Database. Zoological Museum Hamburg. Available online: https://reptile-database.reptarium.cz/species?genus=Rhabdophis&species=tigrinus (accessed on 19 July 2017).
- Morokuma, K.; Kobori, N.; Fukuda, T.; Uchida, T.; Sakai, A.; Toriba, M.; Ohkuma, K.; Nakai, K.; Kurata, T.; Takahashi, M. Experimental manufacture of equine antivenom against Yamakagashi (Rhabdophis tigrinus). Jpn. J. Infect. Dis. 2011, 64, 397–402. [Google Scholar] [PubMed]
- Hifumi, T.; Sakai, A.; Yamamoto, A.; Murakawa, M.; Ato, M.; Shibayama, K.; Ginnaga, A.; Kato, H.; Koido, Y.; Inoue, J.; et al. Clinical characteristics of yamakagashi (Rhabdophis tigrinus) bites: A national survey in Japan, 2000–2013. J. Intensiv. Care 2014, 2, 19. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A. Diagnosis and treatment of snakebite by Mamushi and Yamakagashi. Chudoku Kenkyu 2013, 26, 193–199. [Google Scholar] [PubMed]
- World Health Organization. Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; Replacement of Annex 2 of WHO Technical Report Series, No. 964; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Sakai, A.; Honma, M.; Sawai, Y. Study on the toxicity of venoms extracted from Duvernoy’s gland of certain Asian Colubrid snakes. Snake 1984, 16, 16–20. [Google Scholar]
- Morita, T.; Matsumoto, H.; Iwanaga, S.; Sakai, A. A prothrombin activator found in Rhabdophis tigrinus tigrinus (Yamakagashi Snake) venom. In Hemostasis and Animal Venoms; Pirkle, H., Markland, F.S., Jr., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1988; pp. 55–66. [Google Scholar]
- Komori, K.; Konishi, M.; Maruta, Y.; Toriba, M.; Sakai, A.; Matsuda, A.; Hori, T.; Nakatani, M.; Minamino, N.; Akizawa, T. Characterization of a novel metalloproteinase in Duvernoy’s gland of Rhabdophis tigrinus tigrinus. J. Toxicol. Sci. 2006, 31, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Koike, H.; Sugiyama, Y.; Motoyoshi, K.; Wada, T.; Hishinuma, S.; Mita, M.; Morita, T. Cloning and characterization of novel snake venom proteins that block smooth muscle contraction. Eur. J. Biochem. 2002, 269, 2708–2715. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Morita, T. Structure and function of snake venom cysteine-rich secretory proteins. Toxicon 2004, 44, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Juárez, P.; Sanz, L.; Calvete, J.J. Snake venomics: Characterization of protein families in Sistrurus barbouri venom by cysteine mapping, N-terminal sequencing, and tandem mass spectrometry analysis. Proteomics 2004, 4, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Zhou, X.; Chong, M.Z.; D’hoedt, D.; Foo, C.S.; Rajagopalan, N.; Nirthanan, S.; Bertrand, D.; Sivaraman, J.; Kini, R.M. Structural and functional characterization of a novel homodimeric three-finger neurotoxin from the venom of Ophiophagus hannah (king cobra). J. Biol. Chem. 2010, 285, 8302–8315. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.N.; Timoniuk, A.; Maruñak, S.; Teibler, P.; Acosta, O.; Peichoto, M.E. Biochemical and biological analysis of Philodryas baroni (Baron’s green racer; Dipsadidae) venom: Relevance to the findings of human risk assessment. Hum. Exp. Toxicol. 2014, 33, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, S.A.; Kardong, K.V. Properties of Duvernoy’s secretions from opisthoglyphous and aglyphous colubrid snakes. Toxicon 1994, 32, 1161–1185. [Google Scholar] [CrossRef]
- Peichoto, M.E.; Leiva, L.C.; Guaimás Moya, L.E.; Rey, L.; Acosta, O. Duvernoy’s gland secretion of Philodryas patagoniensis from the northeast of Argentina: Its effects on blood coagulation. Toxicon 2005, 45, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, N.G.; Ventura, S.; Dauer, R.; Hodgson, W.C. A biochemical and pharmacological examination of Rhamphiophis oxyrhynchus (Rufous beaked snake) venom. Toxicon 2005, 45, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Kamiguti, A.S.; Theakstonk, R.D.; Shermanm, N.; Fox, J.W. Mass spectrophotometric evidence for P-III/P-IV metalloproteinases in the venom of the Boomslang (Dispholidus typus). Toxicon 2000, 38, 1613–1620. [Google Scholar] [CrossRef]
- Tsai, I.H.; Chen, Y.H.; Wang, Y.M. Comparative proteomics and subtyping of venom phospholipases A2 and disintegrins of Protobothrops pit vipers. Biochim. Biophys. Acta 2004, 1702, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Aurell, L.; Friberger, P.; Karlsson, G.; Claeson, G. A new sensitive and highly specific chromogenic peptide substrate for factor Xa. Thromb. Res. 1977, 11, 595–609. [Google Scholar] [CrossRef]
- Axelsson, G.; Korsan-Bengtsen, K.; Waldenström, J. Prothrombin determination by means of a chromogenic peptide substrate. Thromb. Haemost. 1976, 36, 517–524. [Google Scholar] [PubMed]
- Nagase, H.; Fields, C.G.; Fields, G.B. Design and characterization of a fluorogenic substrate selectively hydrolyzed by stromelysin 1 (matrix metalloproteinase-3). J. Biol. Chem. 1994, 269, 20952–20957. [Google Scholar] [PubMed]
- Knigh, C.G.; Willenbrock, F.; Murphy, G. A novel coumarin-labelled peptide for sensitive continuous assays of the matrix metalloproteinases. FEBS Lett. 1992, 296, 263–266. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Assay Condition | Clotting Time (s) * | |
|---|---|---|
| Control (10× normal plasma) | ||
| Prothrombin time † | 33 ± 1.2 | |
| Activated partial thromboplastin time ‡ | 109 ± 0.8 | |
| Plasma (10×) + 5 μg of Venom | R. tigrinus | R. lateralis |
| Clotting time without reagents § | 155 ± 3.7 | 168 ± 11.1 |
| Prothrombin time † | 25 ± 0.5 | 25 ± 0.5 |
| Activated partial thromboplastin time ‡ | 48 ± 4.5 | 46 ± 3.3 |
| Substrate | Activity (ΔA405 nm/min/mg Protein) | |
|---|---|---|
| R. tigrinus | R. lateralis | |
| Fraction 1 | Fraction 1 | |
| normal plasma | ||
| S-2222 * | 3.2 | 4.2 |
| S-2238 † | 1001.7 | 1493.4 |
| factor II-deficient plasma ‡ | ||
| S-2238 | 170.0 | 214.0 |
| Substrate | Proteolytic Activity (nmole Substrate Hydrolyzed/min/mg Protein) | |||
|---|---|---|---|---|
| R. tigrinus | R. lateralis | |||
| Fraction 1 | Fraction 2 | Fraction 1 | Fraction 2 | |
| NFF-2 * | 0.043 | 0 | 0.327 | 0 |
| NFF-3 † | 0.441 | 0 | 0.283 | 0 |
| 3163v ‡ | 0.431 | 0.010 | 1.287 | 0 |
| Digested Fragment | By R. tigrinus Fraction 1 | By R. lateralis Fraction 1 | ||
|---|---|---|---|---|
| m/z† | N-terminal Sequence ‡ | m/z† | N-terminal Sequence ‡ | |
| fragment 1 | n.d. # | – | 1189.5 | F V N |
| fragment 2 | 1601.9 | F V N | 1601.8 | F V N |
| fragment 3 | n.d. # | – | 1522.7 | V C G |
| 1423.7 | C G E | |||
| fragment 4 | 1635.8 | L V C | 1635.7 | L V C |
| fragment 5 | 1878.1 | F V N | 1877.9 | F V N |
| fragment 6 | 1912.0 | L Y L | 1911.9 | L Y L |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komori, Y.; Hifumi, T.; Yamamoto, A.; Sakai, A.; Ato, M.; Sawabe, K.; Nikai, T. Comparative Study of Biological Activities of Venom from Colubrid Snakes Rhabdophis tigrinus (Yamakagashi) and Rhabdophis lateralis. Toxins 2017, 9, 373. https://doi.org/10.3390/toxins9110373
Komori Y, Hifumi T, Yamamoto A, Sakai A, Ato M, Sawabe K, Nikai T. Comparative Study of Biological Activities of Venom from Colubrid Snakes Rhabdophis tigrinus (Yamakagashi) and Rhabdophis lateralis. Toxins. 2017; 9(11):373. https://doi.org/10.3390/toxins9110373
Chicago/Turabian StyleKomori, Yumiko, Toru Hifumi, Akihiko Yamamoto, Atsushi Sakai, Manabu Ato, Kyoko Sawabe, and Toshiaki Nikai. 2017. "Comparative Study of Biological Activities of Venom from Colubrid Snakes Rhabdophis tigrinus (Yamakagashi) and Rhabdophis lateralis" Toxins 9, no. 11: 373. https://doi.org/10.3390/toxins9110373
APA StyleKomori, Y., Hifumi, T., Yamamoto, A., Sakai, A., Ato, M., Sawabe, K., & Nikai, T. (2017). Comparative Study of Biological Activities of Venom from Colubrid Snakes Rhabdophis tigrinus (Yamakagashi) and Rhabdophis lateralis. Toxins, 9(11), 373. https://doi.org/10.3390/toxins9110373