
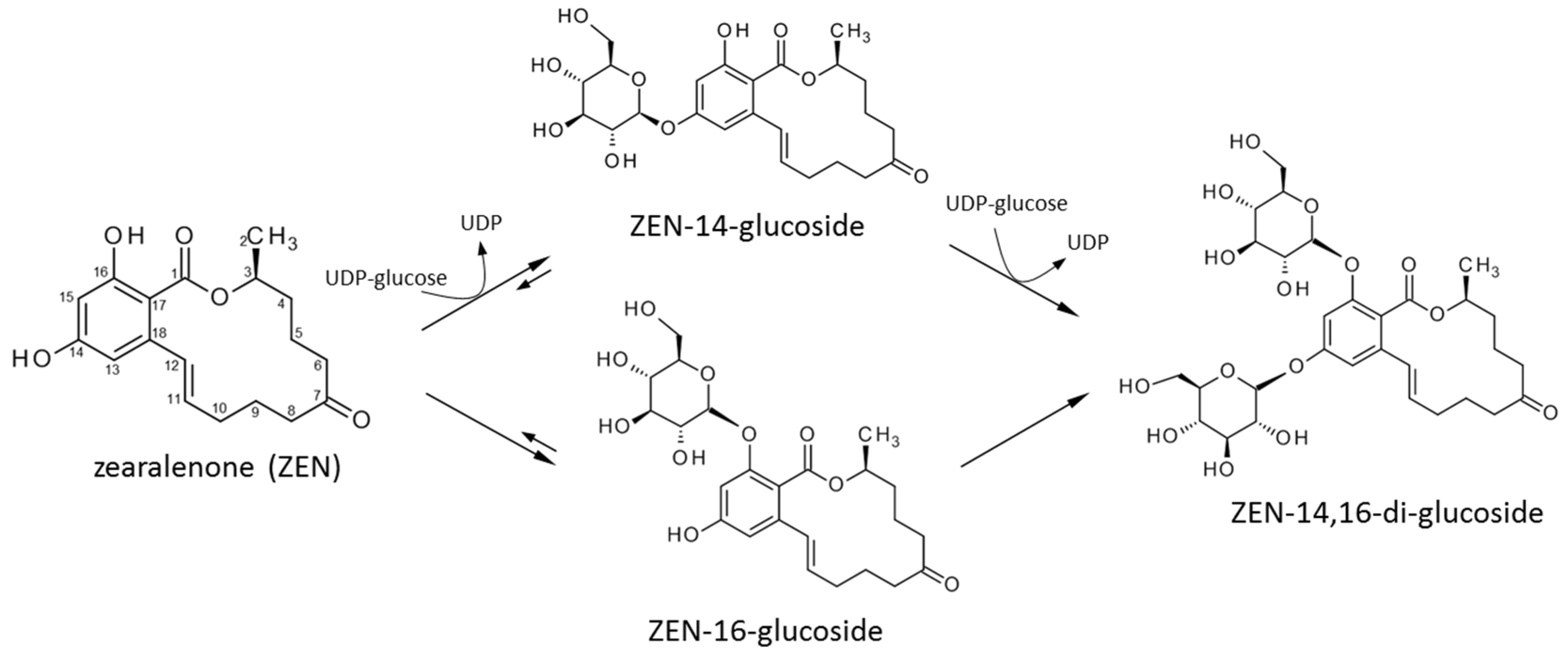
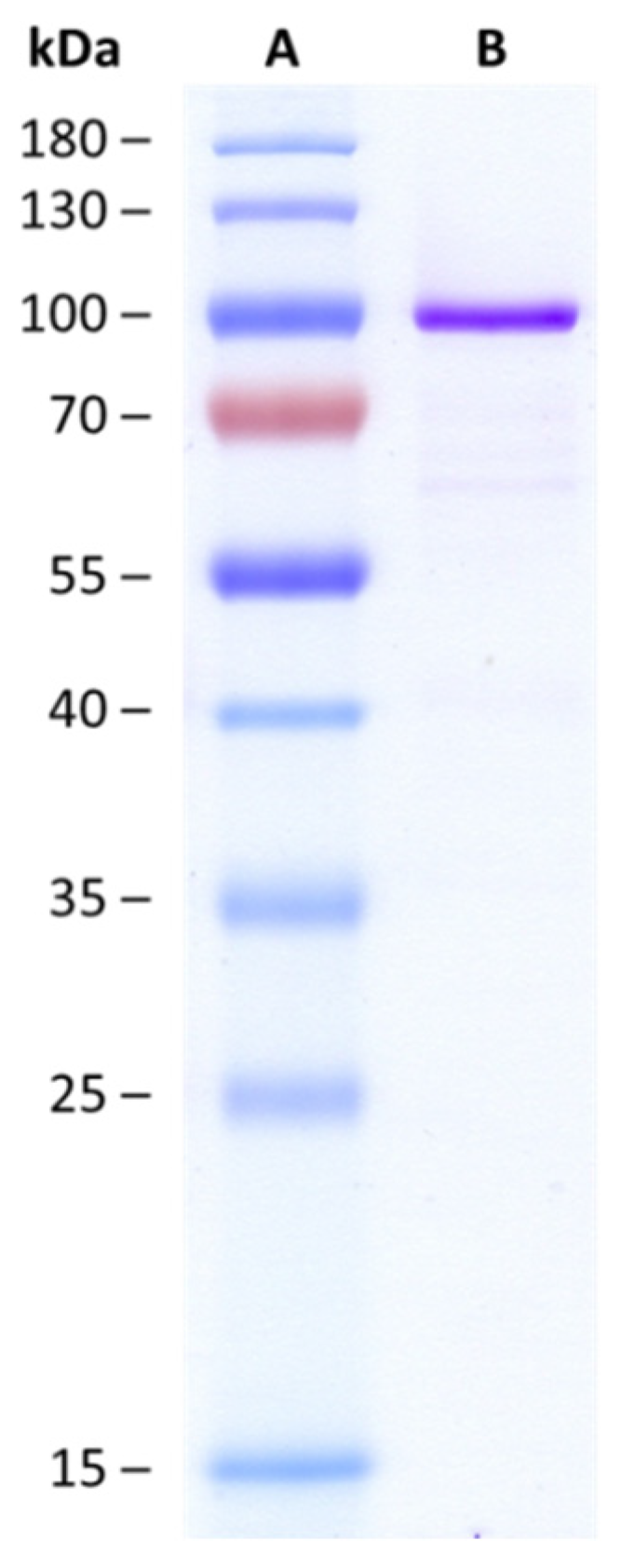
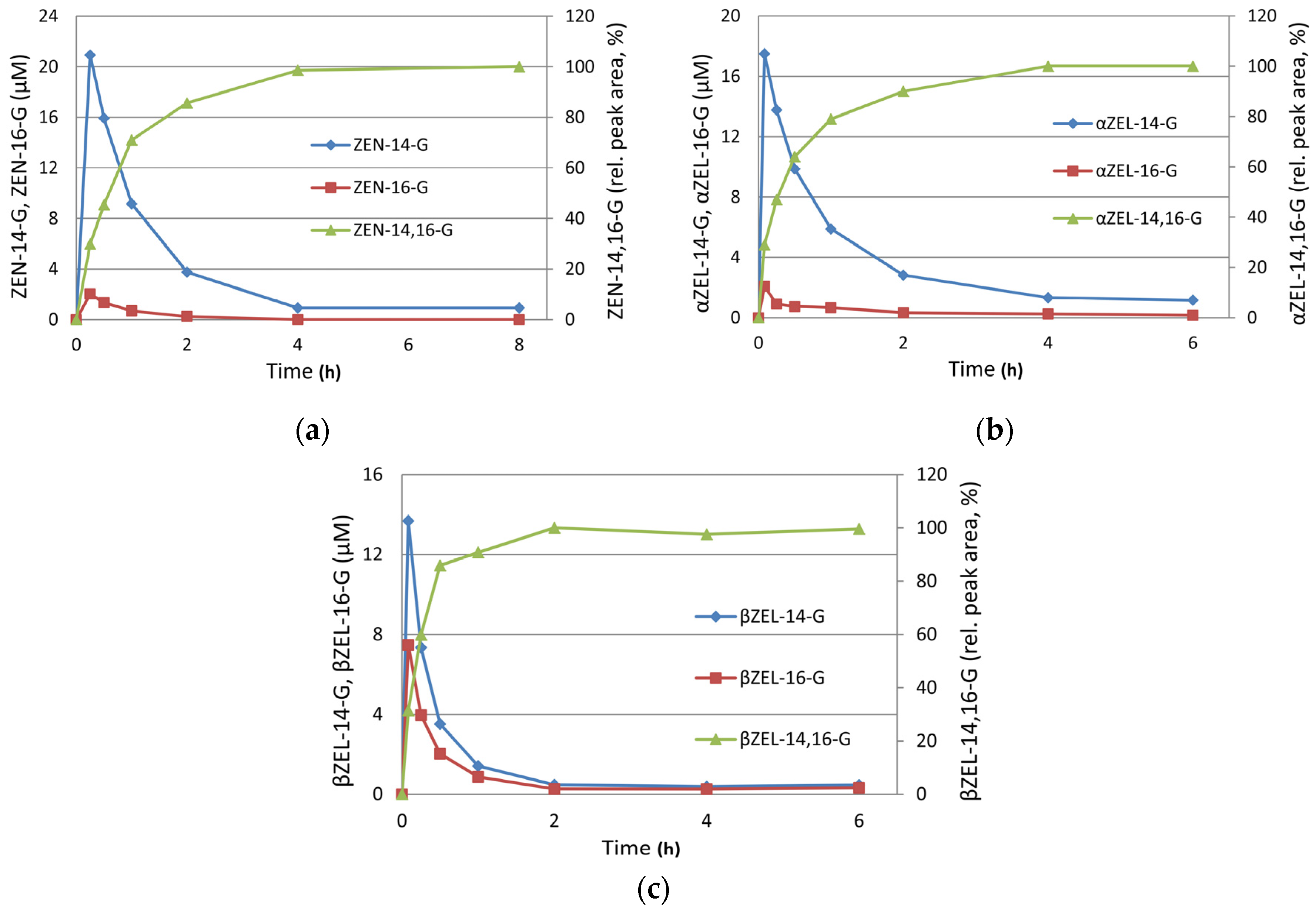
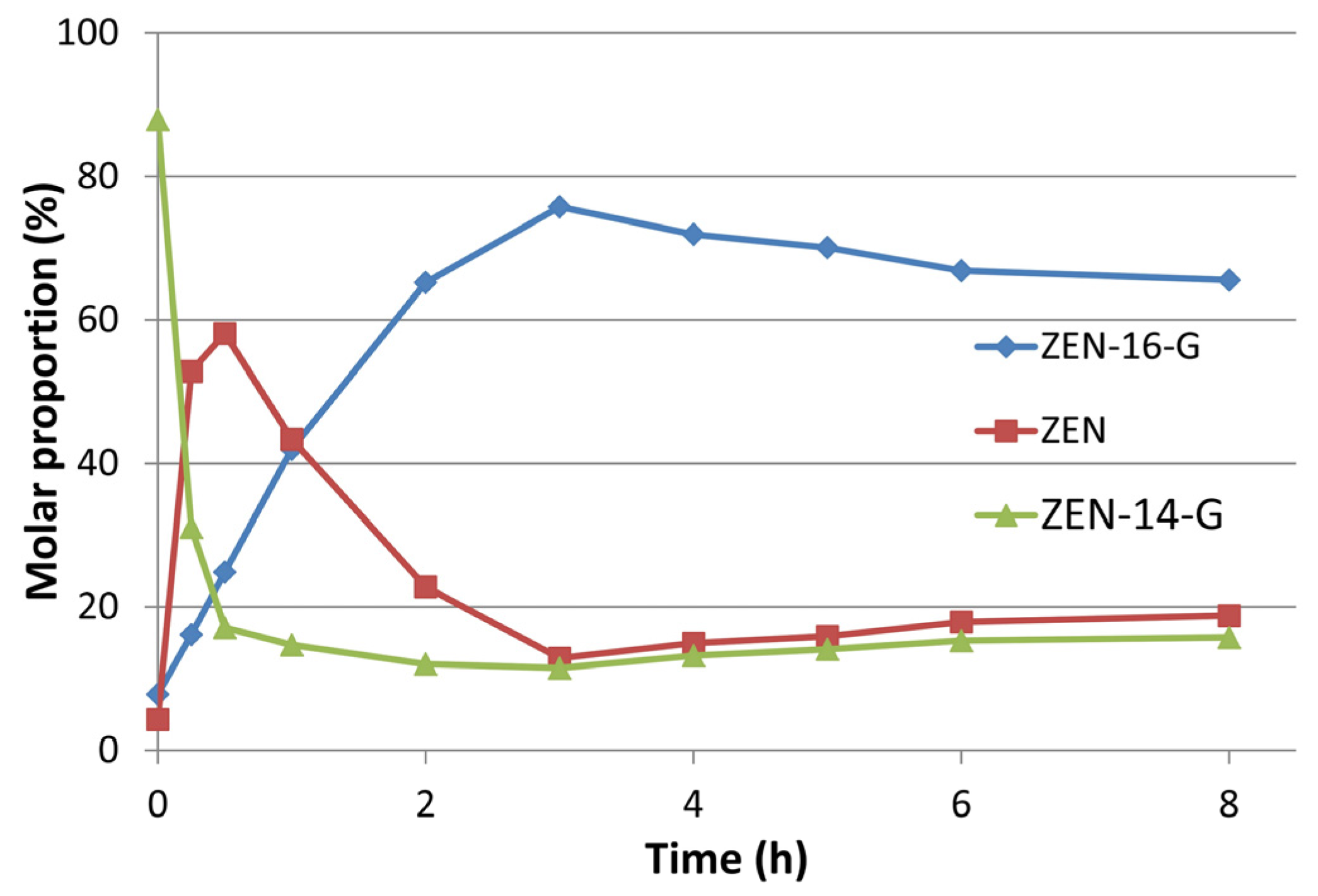
Synthesis of Mono- and Di-Glucosides of Zearalenone and α-/β-Zearalenol by Recombinant Barley Glucosyltransferase HvUGT14077

 ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Michlmayr, H.; Varga, E.; Lupi, F.; Malachová, A.; Hametner, C.; Berthiller, F.; Adam, G. Synthesis of Mono- and Di-Glucosides of Zearalenone and α-/β-Zearalenol by Recombinant Barley Glucosyltransferase HvUGT14077. Toxins 2017, 9, 58. https://doi.org/10.3390/toxins9020058
Michlmayr H, Varga E, Lupi F, Malachová A, Hametner C, Berthiller F, Adam G. Synthesis of Mono- and Di-Glucosides of Zearalenone and α-/β-Zearalenol by Recombinant Barley Glucosyltransferase HvUGT14077. Toxins. 2017; 9(2):58. https://doi.org/10.3390/toxins9020058
Chicago/Turabian StyleMichlmayr, Herbert, Elisabeth Varga, Francesca Lupi, Alexandra Malachová, Christian Hametner, Franz Berthiller, and Gerhard Adam. 2017. "Synthesis of Mono- and Di-Glucosides of Zearalenone and α-/β-Zearalenol by Recombinant Barley Glucosyltransferase HvUGT14077" Toxins 9, no. 2: 58. https://doi.org/10.3390/toxins9020058
APA StyleMichlmayr, H., Varga, E., Lupi, F., Malachová, A., Hametner, C., Berthiller, F., & Adam, G. (2017). Synthesis of Mono- and Di-Glucosides of Zearalenone and α-/β-Zearalenol by Recombinant Barley Glucosyltransferase HvUGT14077. Toxins, 9(2), 58. https://doi.org/10.3390/toxins9020058