
Microcystin?LR Detected in a Low Molecular Weight Fraction from a Crude Extract of Zoanthus sociatus

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Gel Filtration in Sephadex G-50
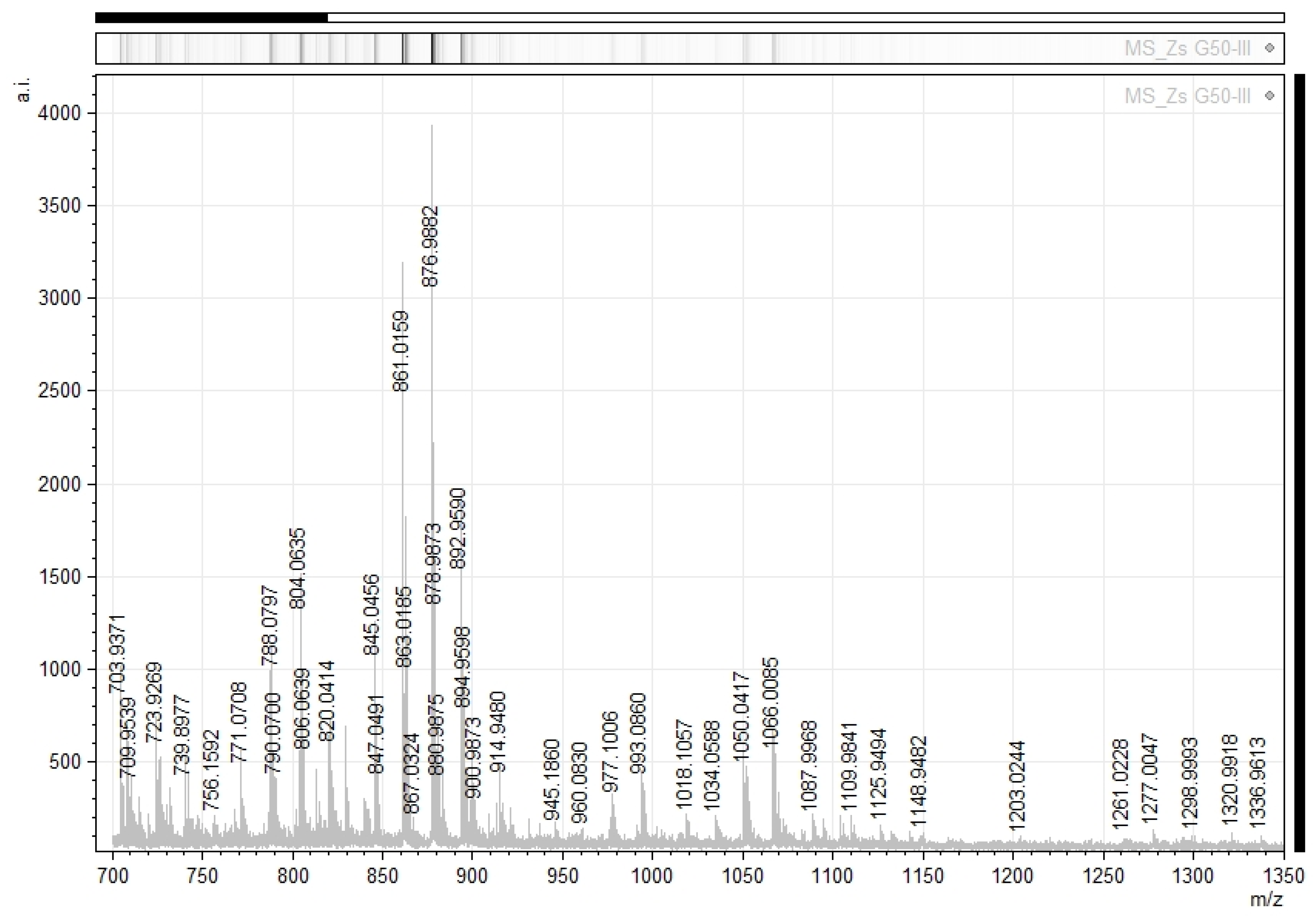
2.2. Zs G50-III MALDI-TOF/TOF Analysis
2.3. Zs G50-III Reserved-Phase Chromatographic Analysis
2.3.1. RP-HPLC Analytical Profile
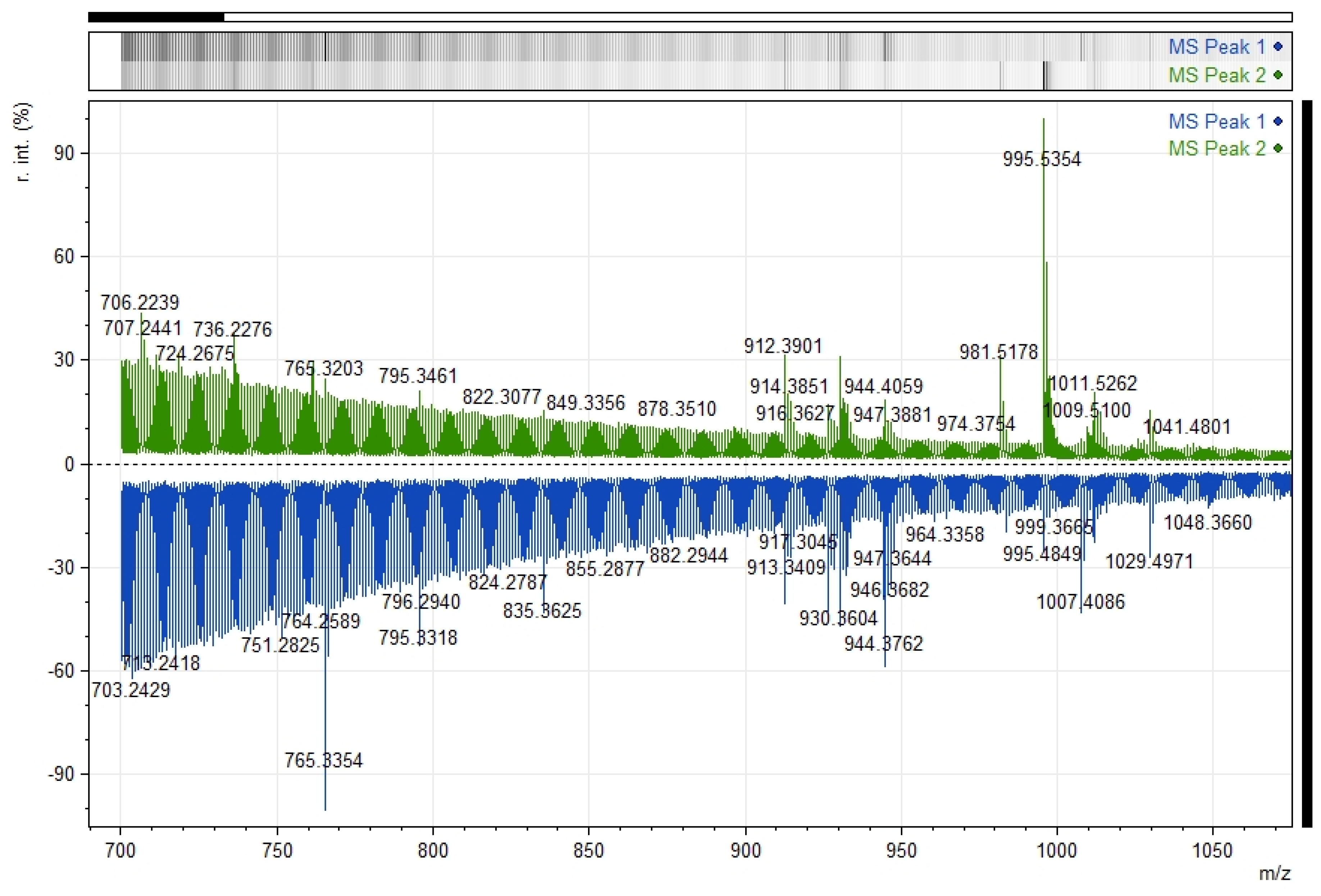
2.3.2. Zs G50-III RP-HPLC Semi-Preparative Assays
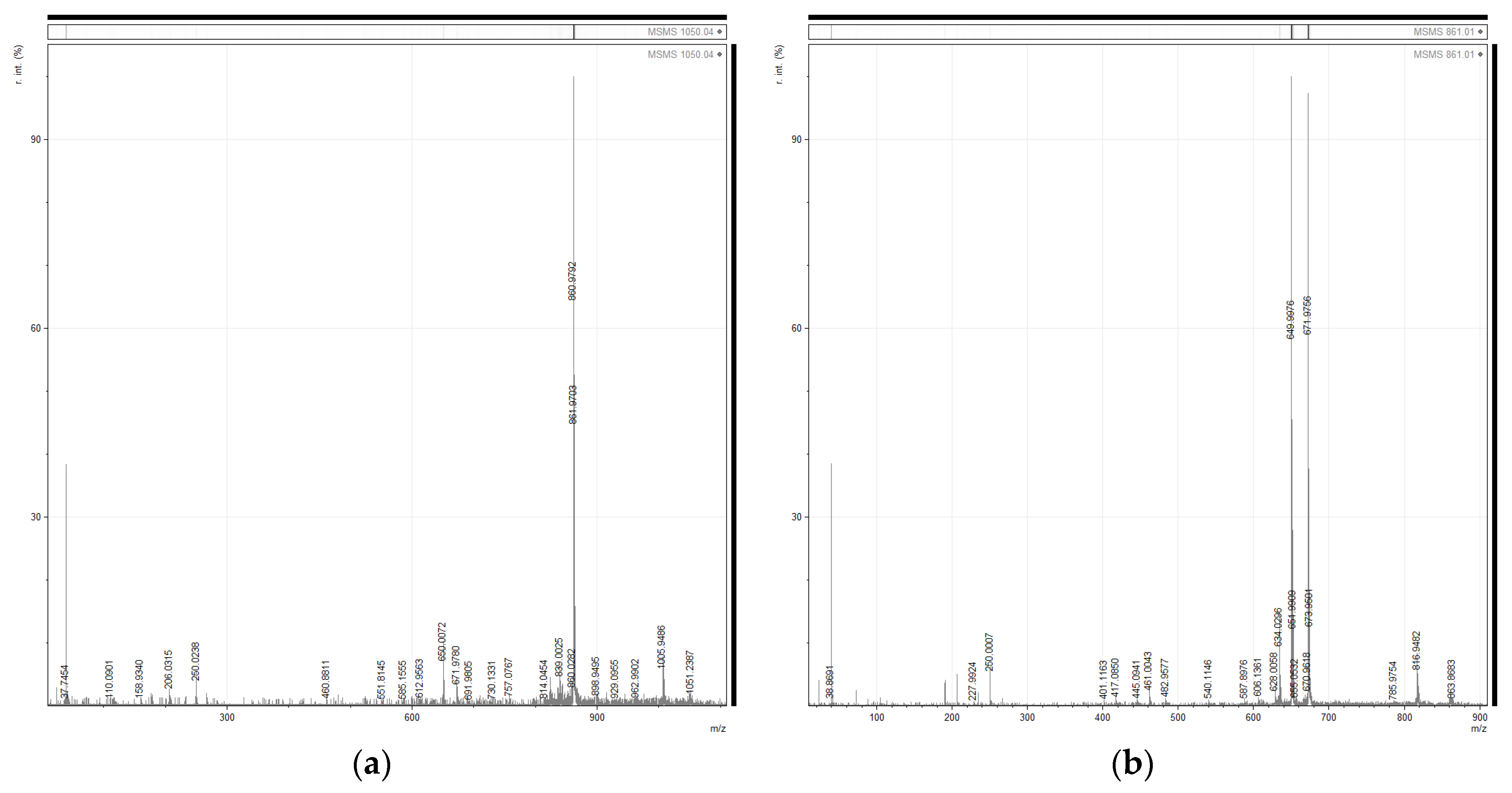
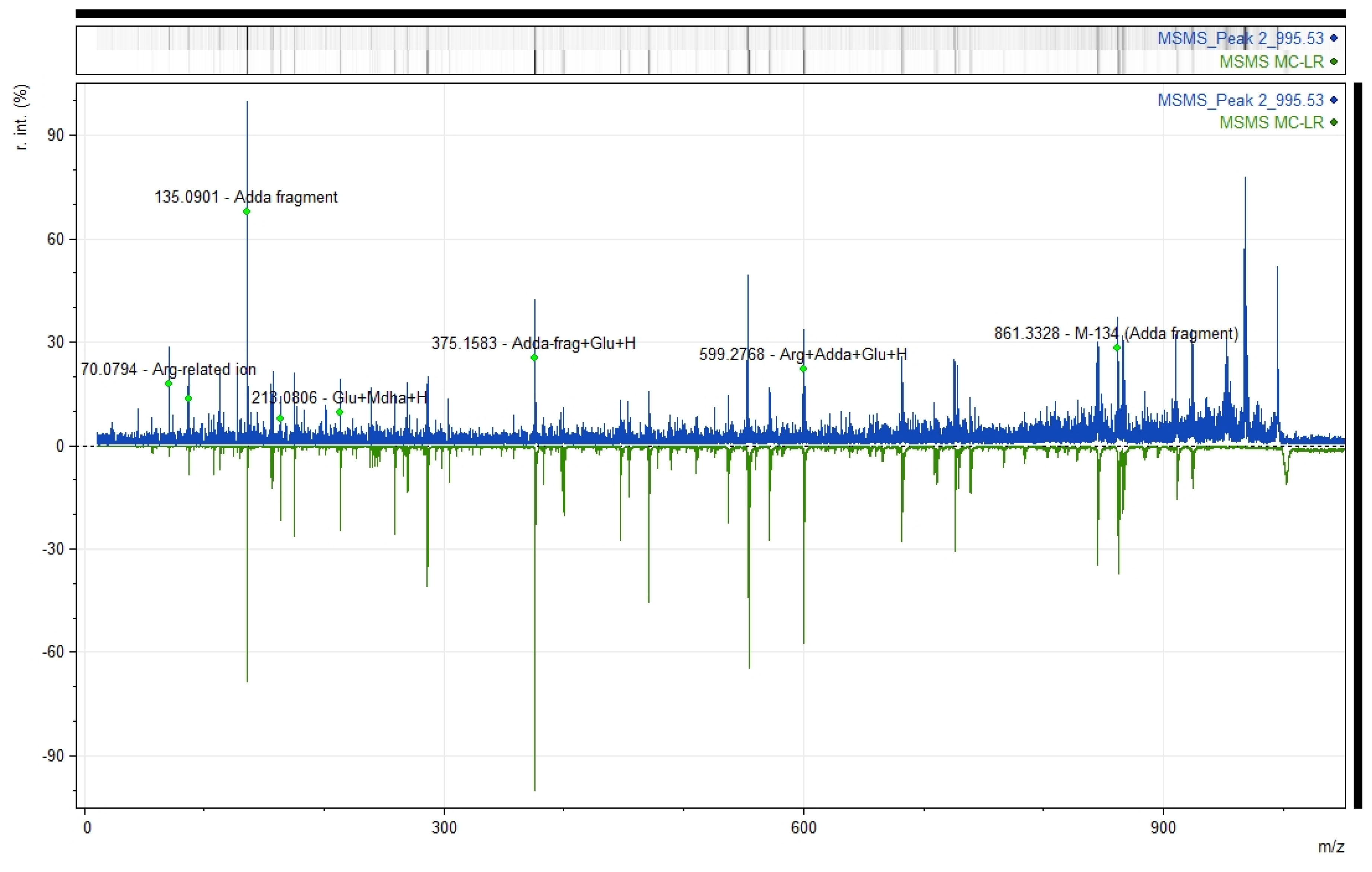
2.4. Identification of MC-LR by MALDI-TOF/TOF Analysis
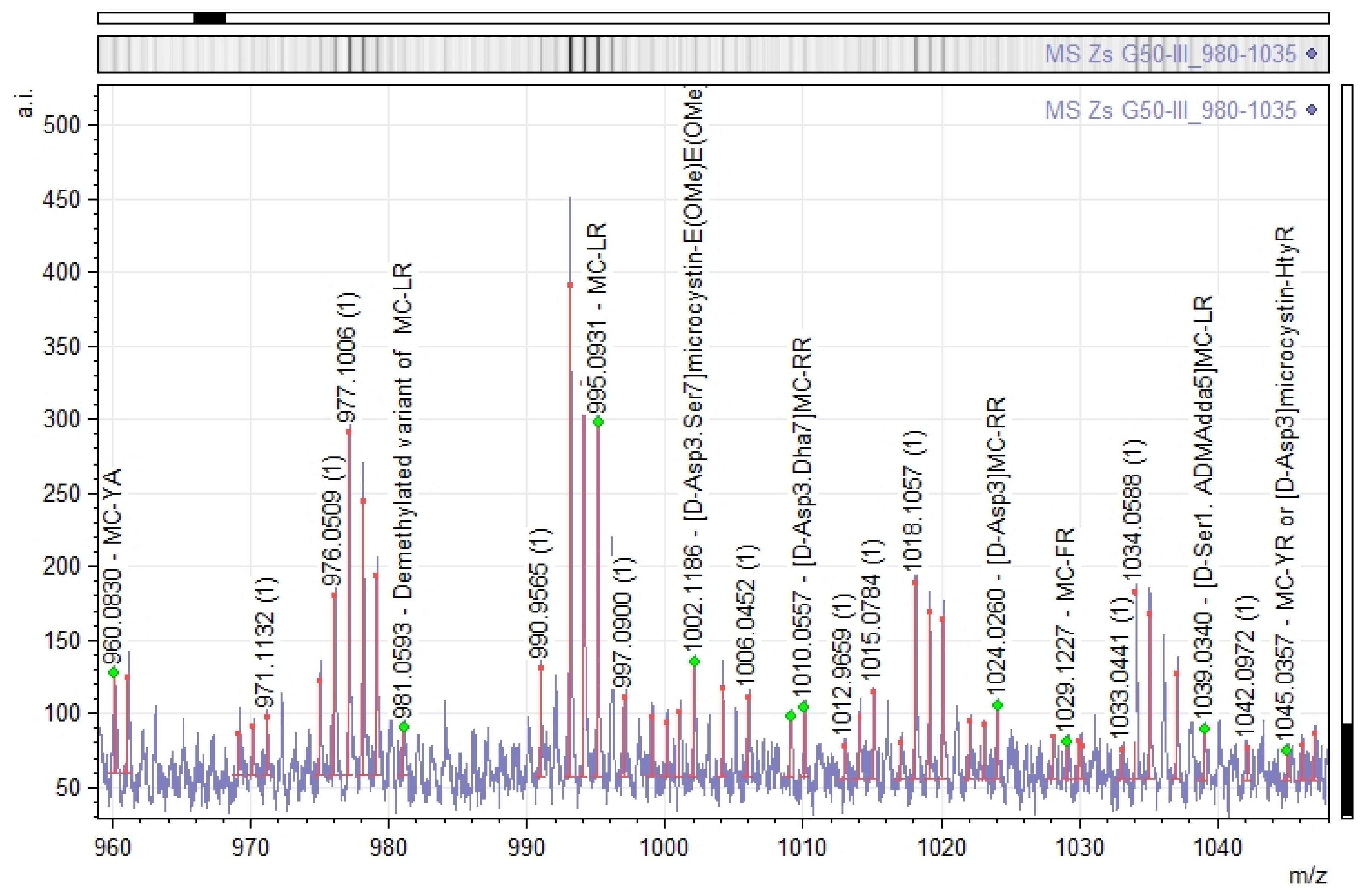
2.5. Putative Origins of Microcystins in the Zs G50-III Fraction
3. Conclusions
4. Materials and Methods
4.1. Preparation of Crude Extract and Gel Filtration on Sephadex G-50
4.2. Mass Spectrometry Analysis and Database Search
MALDI-TOF/TOF Raw Data Availability
4.3. Reserved-Phase Chromatographic Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| LMW | Low molecular weight |
| MALDI-TOF/TOF | Matrix assisted laser desorption/ionization time-of-fly/time-of-fly |
| MS | Mass spectra |
| m/z | Mass-to-charge ratio |
| a.i | Absolute intensity |
| int | Peak’s corrected intensity |
| r.int | Relative intensity |
| S/N | Peak’s signal-to-noise ratio |
| mass | Neutral mass |
| fwhm | Full width at half maximum |
| resol | resolution |
| NORINE | Non-ribosomal peptide database |
| HPLC | High Performance Liquid Chromatographic |
| RP | Reverse Phase |
| MC-LR | Microcystin LR |
| Z. sociatus | Zoanthus sociatus |
| TTX | tetrodotoxin |
| Palythoa caribaeorum | P. caribaeourum |
| LC/ESI-Q-ToF | Liquid Chromatography/Electrospray ionization coupled to Quadruple Time-of-flight |
| Adda | 3-amino-9-methoxy-2,6,8-trimethyl-10-phenyl-deca-4,6-dienoic acid |
| LD | Lethal Dose |
| UV | Ultraviolet |
| TFA | Trifluoroacetic acid |
| PDA | Photodiode Array |
References
- Yan, L.; Herrington, J.; Goldberg, E.; Dulski, P.M.; Bugianesi, R.M.; Slaughter, R.S.; Banerjee, P.; Brochu, R.M.; Priest, B.T.; Kaczorowski, G.J.; et al. Stichodactyla helianthus peptide, a pharmacological tool for studying Kv3.2 channels. Mol. Pharmacol. 2005, 67, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Tejuca, M.; Diaz, I.; Figueredo, R.; Roque, L.; Pazos, F.; Martinez, D.; Iznaga-Escobar, N.; Perez, R.; Alvarez, C.; Lanio, M.E. Construction of an immunotoxin with the pore forming protein StI and ior C5, a monoclonal antibody against a colon cancer cell line. Int. Immunopharmacol. 2004, 4, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Beeton, C.; Pennington, M.W.; Wulff, H.; Singh, S.; Nugent, D.; Crossley, G.; Khaytin, I.; Calabresi, P.A.; Chen, C.Y.; Gutman, G.A.; et al. Targeting effector memory T cells with a selective peptide inhibitor of Kv1.3 channels for therapy of autoimmune diseases. Mol. Pharmacol. 2005, 67, 1369–1381. [Google Scholar] [CrossRef] [PubMed]
- Chi, V.; Pennington, M.W.; Norton, R.S.; Tarcha, E.J.; Londono, L.M.; Sims-Fahey, B.; Upadhyay, S.K.; Lakey, J.T.; Iadonato, S.; Wulff, H.; et al. Development of a sea anemone toxin as an immunomodulator for therapy of autoimmune diseases. Toxicon 2012, 59, 529–546. [Google Scholar] [CrossRef] [PubMed]
- Honma, T.; Shiomi, K. Peptide toxins in sea anemones: Structural and functional aspects. Mar. Biotechnol. 2006, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.S.; Fuentes-Silva, D.; King, G.F. Development of a rational nomenclature for naming peptide and protein toxins from sea anemones. Toxicon 2012, 60, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Frazão, B.; Vasconcelos, V.; Antunes, A. Sea anemone (cnidaria, anthozoa, actiniaria) toxins: An overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef] [PubMed]
- Norton, R.S. Structures of sea anemone toxins. Toxicon 2009, 54, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Beress, L. Biologically active compounds from coelenterates. Pure Appl. Chem. 1982, 54, 1981–1994. [Google Scholar] [CrossRef]
- Wu, C.H. Palytoxin: Membrane mechanisms of action. Toxicon 2009, 54, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Deeds, J.R.; Handy, S.M.; White, K.D.; Reimer, J.D. Palytoxin found in Palythoa sp. zoanthids (anthozoa, hexacorallia) sold in the home aquarium trade. PLoS ONE 2011, 6, e18235. [Google Scholar] [CrossRef] [PubMed]
- Lazcano-Perez, F.; Vivas, O.; Roman-Gonzalez, S.A.; Rodriguez-Bustamante, E.; Castro, H.; Arenas, I.; Garcia, D.E.; Sanchez-Puig, N.; Arreguin-Espinosa, R. A purified Palythoa venom fraction delays sodium current inactivation in sympathetic neurons. Toxicon 2014, 82, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Chen Huang, J.-E.R.M.; Zhou, H.; Lima, E.P.; Gomes, P.B.; Perez, C.D.; Lou, I.; Lee, S.M.Y.; Radis-Baptista, G. Basal repertoire of toxin-related transcripts in the zoanthid Protopalythoa variabilis (cnidaria, anthozoa). CJPT 2015, 29, 104–105. [Google Scholar]
- Kelecom, A. Studies of Brazilian marine invertebrates. VIII. Zoanthosterol, a new sterol from the zoanthid Zoanthus sociatus (hexacorallia, zoanthidea). Bull. Soc. Chim. Belges 1981, 90, 971–976. [Google Scholar] [CrossRef]
- Fukuzawa, S.; Hayashi, Y.; Uemura, D. The isolation and structures of five new alkaloids, norzoanthamine, oxyzoanth amine, norzoanthaminone, cyclozoanthamine and epinorzoanthamine. Heterocycl. Commun. 1995, 1, 207–214. [Google Scholar] [CrossRef]
- Cen-Pacheco, F.; Martin, M.N.; Fernandez, J.J.; Hernandez Daranas, A. New oxidized zoanthamines from a Canary Islands Zoanthus sp. Mar. Drugs 2014, 12, 5188–5196. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, V.; Saxena, A.; Pandey, K.; Bajpai, P.; Misra-Bhattacharya, S. Antifilarial activity of Zoanthus species (phylum coelenterata, class anthzoa) against human lymphatic filaria, Brugia malayi. Parasitol. Res. 2004, 93, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Garcia, C.M.; Sanchez-Soto, C.; Fuentes-Silva, D.; Leon-Pinzon, C.; Dominguez-Perez, D.; Varela, C.; Rodriguez-Romero, A.; Castaneda, O.; Hiriart, M. Low molecular weight compounds from Zoanthus sociatus impair insulin secretion via Ca+2 influx blockade and cause glucose intolerance in vivo. Toxicon 2012, 59, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Perez, D.; Diaz-Garcia, C.M.; Garcia-Delgado, N.; Sierra-Gomez, Y.; Castaneda, O.; Antunes, A. Insights into the toxicological properties of a low molecular weight fraction from Zoanthus sociatus (cnidaria). Mar. Drugs 2013, 11, 2873–2881. [Google Scholar] [CrossRef] [PubMed]
- Perkins, D.N.; Pappin, D.J.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Caboche, S.; Pupin, M.; Leclère, V.; Fontaine, A.; Jacques, P.; Kucherov, G. Norine: A database of nonribosomal peptides. Nucleic Acids Res. 2008, 36, D326–D331. [Google Scholar] [CrossRef] [PubMed]
- Caboche, S.; Leclere, V.; Pupin, M.; Kucherov, G.; Jacques, P. Diversity of monomers in nonribosomal peptides: Towards the prediction of origin and biological activity. J. Bacteriol. 2010, 192, 5143–5150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C.; et al. Evidence for a novel marine harmful algal bloom: Cyanotoxin (microcystin) transfer from land to sea otters. PLoS ONE 2010, 5, e12576. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Z.X.; Boland, M.P.; Smillie, M.A.; Klix, H.; Ptak, C.; Andersen, R.J.; Holmes, C.F.B. Identification of protein phosphatase inhibitors of the microcystin class in the marine environment. Toxicon 1993, 31, 1407–1414. [Google Scholar] [CrossRef]
- Lehman, P.W.; Boyer, G.; Hall, C.; Waller, S.; Gehrts, K. Distribution and toxicity of a new colonial Microcystis aeruginosa bloom in the San Francisco Bay Estuary, California. Hydrobiologia 2005, 541, 87–99. [Google Scholar] [CrossRef]
- Matthiensen, A.; Beattie, K.A.; Yunes, J.S.; Kaya, K.; Codd, G.A. [d-leu1]Microcystin-LR, from the cyanobacterium Microcystis RST 9501 and from a Microcystis bloom in the Patos Lagoon estuary, Brazil. Phytochemistry 2000, 55, 383–387. [Google Scholar] [CrossRef]
- Tonk, L.; Bosch, K.; Visser, P.M.; Huisman, J. Salt tolerance of the harmful cyanobacterium Microcystis aeruginosa. Aquat. Microb. Ecol. 2007, 46, 117–123. [Google Scholar] [CrossRef]
- Richardson, L.L.; Sekar, R.; Myers, J.L.; Gantar, M.; Voss, J.D.; Kaczmarsky, L.; Remily, E.R.; Boyer, G.L.; Zimba, P.V. The presence of the cyanobacterial toxin microcystin in black band disease of corals. FEMS Microbiol. Lett. 2007, 272, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Boutet, E.; Lieberherr, D.; Tognolli, M.; Schneider, M.; Bansal, P.; Bridge, A.J.; Poux, S.; Bougueleret, L.; Xenarios, I. Uniprotkb/swiss-prot, the manually annotated section of the uniprot knowledgebase: How to use the entry view. Methods Mol. Biol. (Clifton, N.J.) 2016, 1374, 23–54. [Google Scholar]
- Ferranti, P.; Fabbrocino, S.; Nasi, A.; Caira, S.; Bruno, M.; Serpe, L.; Gallo, P. Liquid chromatography coupled to quadruple time-of-flight tandem mass spectrometry for microcystin analysis in freshwaters: Method performances and characterisation of a novel variant of Microcystin-RR. Rapid Commun. Mass Spectrom. 2009, 23, 1328–1336. [Google Scholar] [CrossRef] [PubMed]
- Mbukwa, E.A.; Msagati, T.A.M.; Mamba, B.B. Quantitative variations of intracellular Microcystin-LR, -RR and -YR in samples collected from four locations in Hartbeespoort Dam in North West province (South Africa) during the 2010/2011 summer season. Int. J. Environ. Res. Public Health 2012, 9, 3484–3505. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Sun, F.; Choi, B.W.; Rinehart, K.L.; Carmichael, W.W.; Evans, W.R.; Beasley, V.R. Seven more microcystins from homer lake cells: Application of the general method for structure assignment of peptides containing. Alpha.,.Beta.-dehydroamino acid unit(s). J. Org. Chem. 1995, 60, 3671–3679. [Google Scholar] [CrossRef]
- Vasas, G.; Szydlowska, D.; Gaspar, A.; Welker, M.; Trojanowicz, M.; Borbely, G. Determination of microcystins in environmental samples using capillary electrophoresis. J. Biochem. Biophys. Methods 2006, 66, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Bogialli, S.; Bruno, M.; Curini, R.; Di Corcia, A.; Lagana, A.; Mari, B. Simple assay for analyzing five microcystins and nodularin in fish muscle tissue: Hot water extraction followed by liquid chromatography-tandem mass spectrometry. J. Agric. Food Chem. 2005, 53, 6586–6592. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Yuan, M.; Sivonen, K.; Carmichael, W.W.; Rinehart, K.L.; Rouhiainen, L.; Sun, F.; Brittain, S.; Otsuki, A. Seven new microcystins possessing two L-glutamic acid units, isolated from Anabaena sp. Strain 186. Chem. Res. Toxicol. 1998, 11, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Stoner, R.D.; Adams, W.H.; Slatkin, D.N.; Siegelman, H.W. The effects of single L-amino acid substitutions on the lethal potencies of the microcystins. Toxicon 1989, 27, 825–828. [Google Scholar] [CrossRef]
- Sivonen, K.; Namikoshi, M.; Evans, W.R.; Carmichael, W.W.; Sun, F.; Rouhiainen, L.; Luukkainen, R.; Rinehart, K.L. Isolation and characterization of a variety of microcystins from seven strains of the cyanobacterial genus Anabaena. Appl. Environ. Microbiol. 1992, 58, 2495–2500. [Google Scholar] [PubMed]
- Meriluoto, J.A.; Sandstrom, A.; Eriksson, J.E.; Remaud, G.; Craig, A.G.; Chattopadhyaya, J. Structure and toxicity of a peptide hepatotoxin from the cyanobacterium Oscillatoria agardhii. Toxicon 1989, 27, 1021–1034. [Google Scholar] [CrossRef]
- Namikoshi, M.; Rinehart, K.L.; Sakai, R.; Stotts, R.R.; Dahlem, A.M.; Beasley, V.R.; Carmichael, W.W.; Evans, W.R. Identification of 12 hepatotoxins from a homer lake bloom of the cyanobacteria Microcystis aeruginosa, Microcystis viridis, and Microcystis wesenbergii: Nine new microcystins. J. Org. Chem. 1992, 57, 866–872. [Google Scholar] [CrossRef]
- Sivonen, K.; Namikoshi, M.; Evans, W.R.; Fardig, M.; Carmichael, W.W.; Rinehart, K.L. Three new microcystins, cyclic heptapeptide hepatotoxins, from Nostoc sp. Strain 152. Chem. Res. Toxicol. 1992, 5, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Sivonen, K.; Evans, W.R.; Carmichael, W.W.; Rouhiainen, L.; Luukkainen, R.; Rinehart, K.L. Structures of three new homotyrosine-containing microcystins and a new homophenylalanine variant from Anabaena sp. Strain 66. Chem. Res. Toxicol. 1992, 5, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, M.R.; Bakshi, K.; Volkin, D.B.; Middaugh, C.R. Ultraviolet Absorption Spectroscopy of Peptides. In Therapeutic Peptides: Methods and Protocols; Nixon, E.A., Ed.; Humana Press: Totowa, NJ, USA, 2014; pp. 225–236. [Google Scholar]
- Scopes, R. Measurement of protein by spectrophotometry at 205 nm. Anal. Biochem. 1974, 59, 277–282. [Google Scholar] [CrossRef]
- Anthis, N.J.; Clore, G.M. Sequence-specific determination of protein and peptide concentrations by absorbance at 205 nm. Protein Sci. Publ. Protein Soc. 2013, 22, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.C.; von Hippel, P.H. Calculation of protein extinction coefficients from amino acid sequence data. Anal. Biochem. 1989, 182, 319–326. [Google Scholar] [CrossRef]
- Zhang, T.; Kiyonami, R.; Wang, L.; Jiang, G. Identification and Quantitation of Microcystins by Targeted Full-Scan LC-MS/MS. In Application Note 569; Thermo-Fisher-Scientific: San Jose, CA, USA, 2016. [Google Scholar]
- Neumann, U.; Campos, V.; Cantarero, S.; Urrutia, H.; Heinze, R.; Weckesser, J.; Erhard, M. Co-occurrence of non-toxic (cyanopeptolin) and toxic (microcystin) peptides in a bloom of Microcystis sp. from a Chilean lake. Syst. Appl. Microbiol. 2000, 23, 191–197. [Google Scholar] [CrossRef]
- Tsuji, K.; Watanuki, T.; Kondo, F.; Watanabe, M.F.; Suzuki, S.; Nakazawa, H.; Suzuki, M.; Uchida, H.; Harada, K.-I. Stability of microcystins from cyanobacteria—II. Effect of UV light on decomposition and isomerization. Toxicon 1995, 33, 1619–1631. [Google Scholar] [CrossRef]
- Martin, C.; Oberer, L.; Ino, T.; Konig, W.A.; Busch, M.; Weckesser, J. Cyanopeptolins, new depsipeptides from the cyanobacterium Microcystis sp. PCC 7806. J. Antibiot. 1993, 46, 1550–1556. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, R.L.; Dörr, F.A.; Dörr, F.; Bortoli, S.; Delherbe, N.; Vásquez, M.; Pinto, E. Co-occurrence of microcystin and microginin congeners in Brazilian strains of Microcystis sp. FEMS Microbiol. Ecol. 2012, 82, 692–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.F.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from Microcystis CAWBG11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef] [PubMed]
- Taton, A.; Grubisic, S.; Balthasart, P.; Hodgson, D.A.; Laybourn-Parry, J.; Wilmotte, A. Biogeographical distribution and ecological ranges of benthic cyanobacteria in East Antarctic Lakes. FEMS Microbiol. Ecol. 2006, 57, 272–289. [Google Scholar] [CrossRef] [PubMed]
- Fumanti, B.; Cavacini, P.; Alfinito, S. Benthic algal mats of some lakes of Inexpressible Island (Northern Victoria Land, Antarctica). Polar Biol. 1997, 17, 25–30. [Google Scholar] [CrossRef]
- Timo, N. Microcystin Congeners Described in the Literature. Available online: https://figshare.Com/articles/_microcystin_congeners_described_in_the_literature/880756 (accessed on 26 January 2017).
- Tillett, D.; Dittmann, E.; Erhard, M.; von Dohren, H.; Borner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Carmichael, W.W. Cyanobacteria secondary metabolites—the cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Zohary, T. Hyperscums of the cyanobacterium Microcystis aeruginosa in a hypertrophic lake (Hartbeespoort Dam, South Africa). J. Plankton Res. 1985, 7, 399–409. [Google Scholar] [CrossRef]
- Ueno, Y.; Nagata, S.; Tsutsumi, T.; Hasegawa, A.; Yoshida, F.; Suttajit, M.; Mebs, D.; Pütsch, M.; Vasconcelos, V. Survey of microcystins in environmental water by a highly sensitive immunoassay based on monoclonal antibody. Nat. Toxins 1996, 4, 271–276. [Google Scholar] [CrossRef]
- Lopes, V.R.; Ramos, V.; Martins, A.; Sousa, M.; Welker, M.; Antunes, A.; Vasconcelos, V.M. Phylogenetic, chemical and morphological diversity of cyanobacteria from Portuguese temperate estuaries. Mar. Environ. Res. 2012, 73, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Frazao, B.; Martins, R.; Vasconcelos, V. Are known cyanotoxins involved in the toxicity of picoplanktonic and filamentous north atlantic marine cyanobacteria? Mar. Drugs 2010, 8, 1908–1919. [Google Scholar] [CrossRef] [PubMed]
- Landsberg, J.H. The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- DeMott, W.R.; Moxter, F. Foraging cyanobacteria by copepods: Responses to chemical defense and resource abundance. Ecology 1991, 72, 1820–1834. [Google Scholar] [CrossRef]
- Malbrouck, C.; Kestemont, P. Effects of microcystins on fish. Environ. Toxicol. Chem. 2006, 25, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Amorim, Á.; Vasconcelos, V.T. Dynamics of microcystins in the mussel Mytilus galloprovincialis. Toxicon 1999, 37, 1041–1052. [Google Scholar] [CrossRef]
- Williams, D.E.; Dawe, S.C.; Kent, M.L.; Andersen, R.J.; Craig, M.; Holmes, C.F.B. Bioaccumulation and clearance of microcystins from salt water mussels, Mytilus edulis, and in vivo evidence for covalently bound microcystins in mussel tissues. Toxicon 1997, 35, 1617–1625. [Google Scholar] [CrossRef]
- Vasconcelos, V.; Oliveira, S.; Teles, F.O. Impact of a toxic and a non-toxic strain of Microcystis aeruginosa on the crayfish Procambarus clarkii. Toxicon 2001, 39, 1461–1470. [Google Scholar] [CrossRef]
- Zimba, P.V.; Camus, A.; Allen, E.H.; Burkholder, J.M. Co-occurrence of white shrimp, Litopenaeus vannamei, mortalities and microcystin toxin in a southeastern USA shrimp facility. Aquaculture 2006, 261, 1048–1055. [Google Scholar] [CrossRef]
- Backer, L.; Carmichael, W.; Kirkpatrick, B.; Williams, C.; Irvin, M.; Zhou, Y.; Johnson, T.; Nierenberg, K.; Hill, V.; Kieszak, S.; et al. Recreational exposure to low concentrations of microcystins during an algal bloom in a small lake. Mar. Drugs 2008, 6, 389–406. [Google Scholar] [CrossRef] [PubMed]
- De Silva, E.D.; Williams, D.E.; Andersen, R.J.; Klix, H.; Holmes, C.F.; Allen, T.M. Motuporin, a potent protein phosphatase inhibitor isolated from the Papua New Guinea sponge Theonella swinhoei Gray. Tetrahedron Lett. 1992, 33, 1561–1564. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Namikoshi, M.; Choi, B.W. Structure and biosynthesis of toxins from blue-green algae (cyanobacteria). J. Appl. Phycol. 1994, 6, 159–176. [Google Scholar] [CrossRef]
- Krishnamurthy, T.; Carmichael, W.W.; Sarver, E.W. Toxic peptides from freshwater cyanobacteria (blue-green algae). I. Isolation, purification and characterization of peptides from Microcystis aeruginosa and Anabaena flos-aquae. Toxicon 1986, 24, 865–873. [Google Scholar] [CrossRef]
- Zhao, Y.; Shi, K.; Su, X.; Xie, L.; Yan, Y. Microcystin-lr induces dysfunction of insulin secretion in rat insulinoma (INS-1) cells: Implications for diabetes mellitus. J. Hazard. Mater. 2016, 314, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Strohalm, M.; Hassman, M.; Košata, B.; Kodíček, M. Mmass data miner: An open source alternative for mass spectrometric data analysis. Rapid Commun. Mass Spectrom. 2008, 22, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Strohalm, M.; Kavan, D.; Novák, P.; Volný, M.; Havlíček, V.R. Mmass 3: A cross-platform software environment for precise analysis of mass spectrometric data. Anal. Chem. 2010, 82, 4648–4651. [Google Scholar] [CrossRef] [PubMed]








© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez‐Pérez, D.; Rodríguez, A.A.; Osorio, H.; Azevedo, J.; Castañeda, O.; Vasconcelos, V.; Antunes, A. Microcystin?LR Detected in a Low Molecular Weight Fraction from a Crude Extract of Zoanthus sociatus. Toxins 2017, 9, 89. https://doi.org/10.3390/toxins9030089
Domínguez‐Pérez D, Rodríguez AA, Osorio H, Azevedo J, Castañeda O, Vasconcelos V, Antunes A. Microcystin?LR Detected in a Low Molecular Weight Fraction from a Crude Extract of Zoanthus sociatus. Toxins. 2017; 9(3):89. https://doi.org/10.3390/toxins9030089
Chicago/Turabian StyleDomínguez‐Pérez, Dany, Armando Alexei Rodríguez, Hugo Osorio, Joana Azevedo, Olga Castañeda, Vítor Vasconcelos, and Agostinho Antunes. 2017. "Microcystin?LR Detected in a Low Molecular Weight Fraction from a Crude Extract of Zoanthus sociatus" Toxins 9, no. 3: 89. https://doi.org/10.3390/toxins9030089
APA StyleDomínguez‐Pérez, D., Rodríguez, A. A., Osorio, H., Azevedo, J., Castañeda, O., Vasconcelos, V., & Antunes, A. (2017). Microcystin?LR Detected in a Low Molecular Weight Fraction from a Crude Extract of Zoanthus sociatus. Toxins, 9(3), 89. https://doi.org/10.3390/toxins9030089