

Crystal Packing Differences as a Key Factor for Stabilization of the N-Terminal Fragment of the Human HINT1 Protein

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning, Expression, and Purification of hHINT1
2.2. Crystallization of hHINT1
2.3. Data Collection, Structure Determination, and Refinement
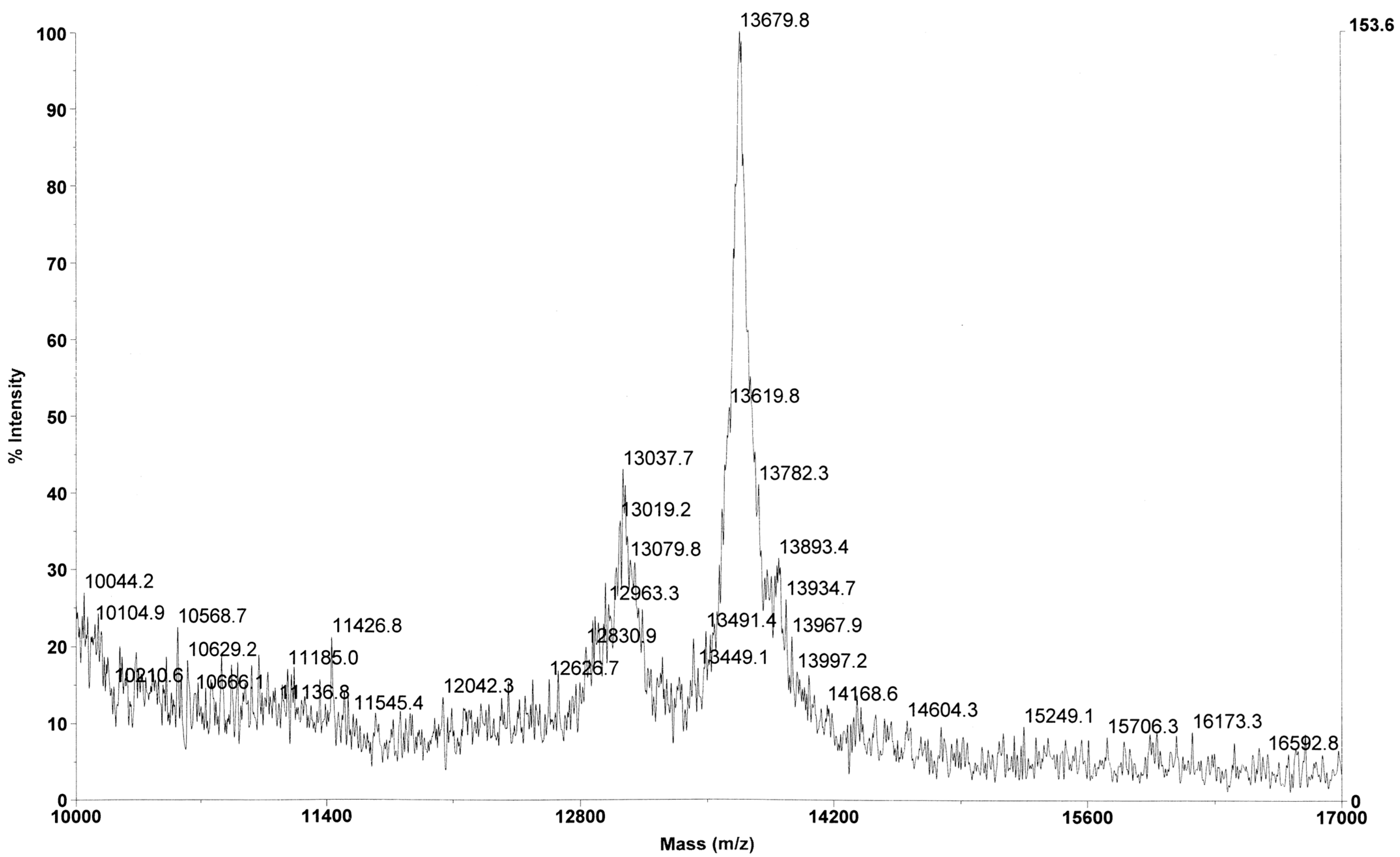
2.4. Mass Spectrometry Analysis
3. Results and Discussion
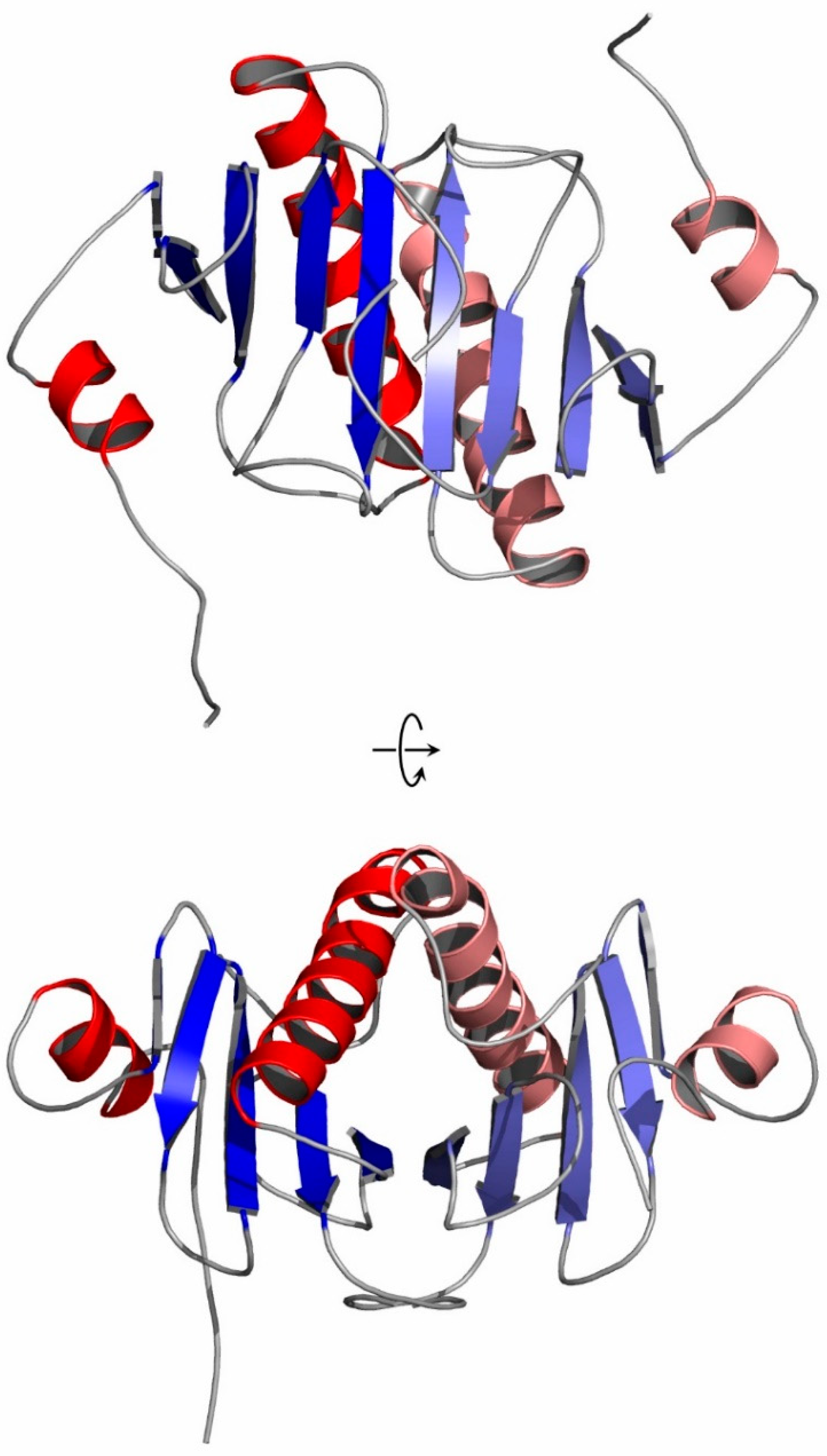
3.1. Overall Structure of a New Crystal Form of hHINT1 in Space Group P212121
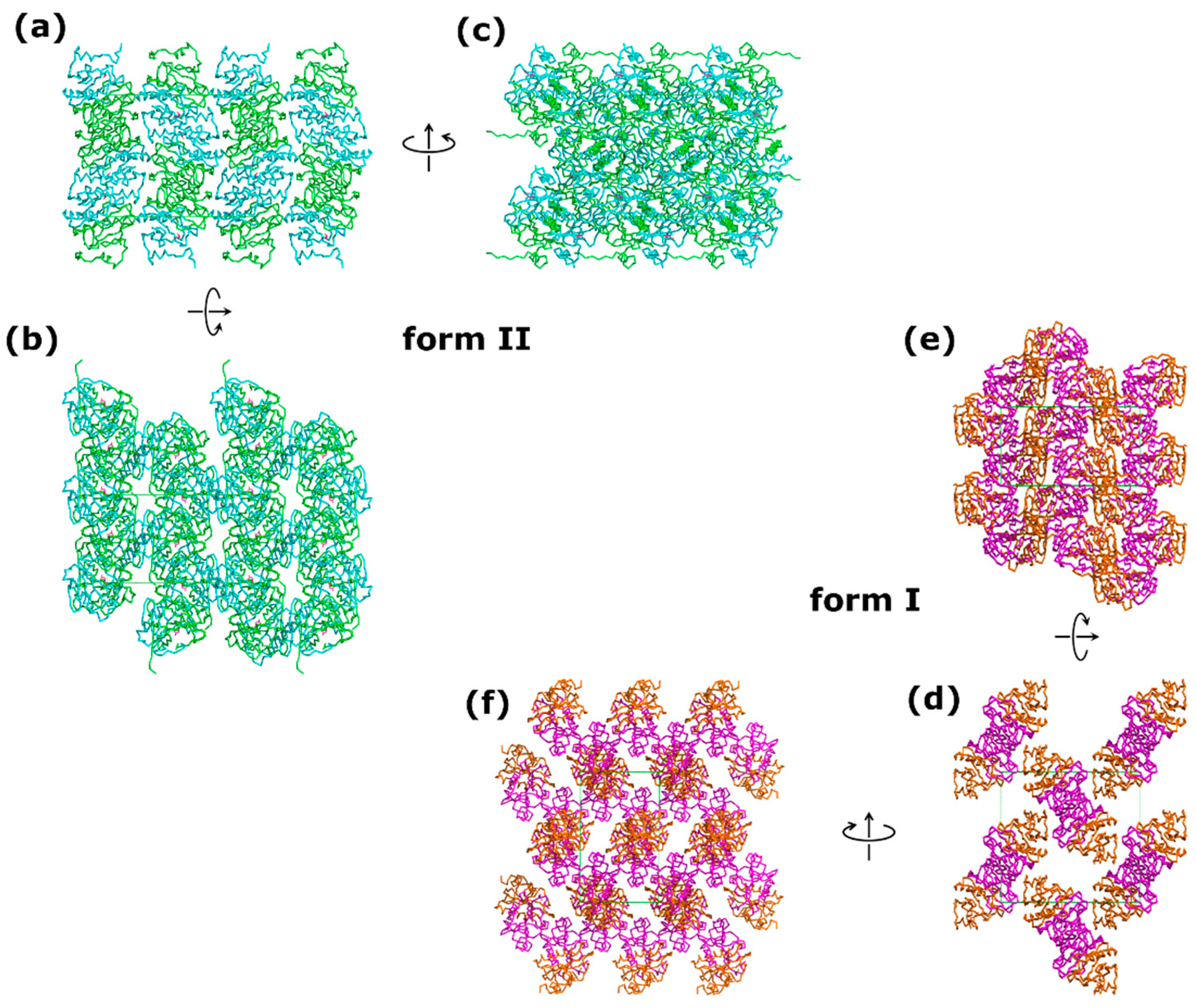
3.2. Two Crystal Forms—The Same Space Group
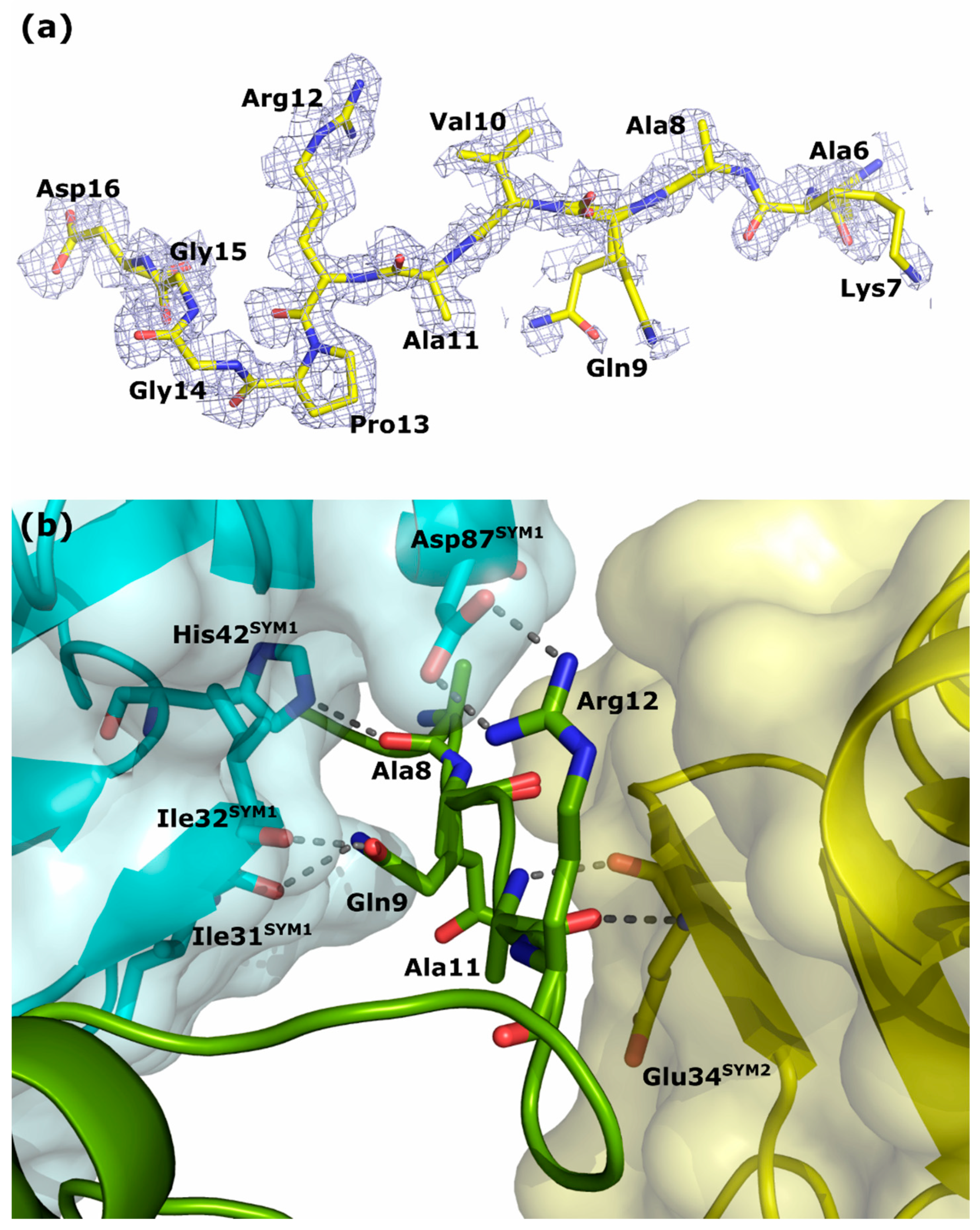
3.3. Structure of the N-Terminus
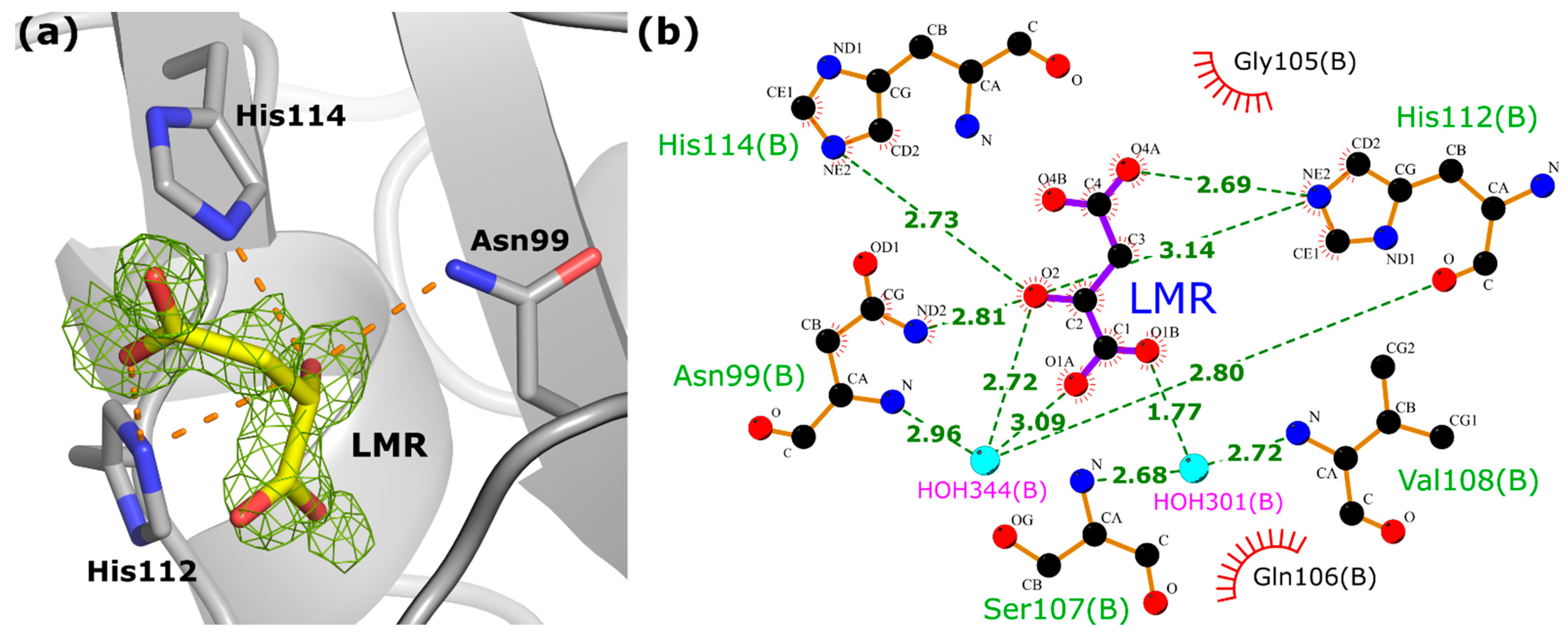
3.4. Analysis of the Nucleotide Binding Site/Ligand Molecule
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brenner, C. Hint, Fhit, and GalT: Function, structure, evolution, and mechanism of three branches of the histidine triad superfamily of nucleotide hydrolases and transferases. Biochemistry 2002, 41, 9003–9014. [Google Scholar] [PubMed] [Green Version]
- Bieganowski, P.; Garrison, P.N.; Hodawadekar, S.C.; Faye, G.; Barnes, L.D.; Brenner, C. Adenosine monophosphoramidase activity of Hint and Hnt1 supports function of Kin28, Ccl1, and Tfb3. J. Biol. Chem. 2002, 277, 10852–10860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, K.P.; Seidle, H.; Wright, N.; Sperry, J.B.; Bieganowski, P.; Howitz, K.; Wright, D.L.; Brenner, C. Altered specificity of Hint-W123Q supports a role for Hint inhibition by ASW in avian sex determination. Physiol. Genom. 2004, 20, 12–14. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Wu, X.; Deng, H.; Huang, Y. First identification and functional analysis of a histidine triad nucleotide binding protein in an invertebrate species Haliotis diversicolor supertexta. Dev. Comp. Immunol. 2010, 34, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Guranowski, A.; Wojdyla, A.M.; Rydzik, A.M.; Stepinski, J.; Jemielity, J. Plant nucleoside 5′-phosphoramidate hydrolase; simple purification from yellow lupin (Lupinus luteus) seeds and properties of homogenous enzyme. Acta Biochim. Pol. 2011, 58, 131–136. [Google Scholar]
- Korsisaari, N.; Rossi, D.J.; Luukko, K.; Huebner, K.; Henkemeyer, M.; Mäkelä, T.P. The histidine triad protein Hint is not required for murine development or Cdk7 function. Mol. Cell. Biol. 2003, 23, 3929–3935. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.-F.; Bieganowski, P.; Shilinski, K.; Cheng, J.; Brenner, C.; Wagner, C.R. 31P NMR and genetic analysis establish hint as the only Escherichia coli purine nucleoside phosphoramidase and as essential for growth under high salt conditions. J. Biol. Chem. 2005, 280, 15356–15361. [Google Scholar] [CrossRef] [Green Version]
- Bardaweel, S.; Pace, J.; Chou, T.F.; Cody, V.; Wagner, C.R. Probing the impact of the echinT C-terminal domain on structure and catalysis. J. Mol. Biol. 2010, 404, 627–638. [Google Scholar]
- Baugh, L.; Phan, I.; Begley, D.W.; Clifton, M.C.; Armour, B.; Dranow, D.M.; Taylor, B.M.; Muruthi, M.M.; Abendroth, J.; Fairman, J.W.; et al. Increasing the structural coverage of tuberculosis drug targets. Tuberculosis 2015, 95, 142–148. [Google Scholar]
- Lorimer, D.D.; Choi, R.; Abramov, A.; Nakazawa Hewitt, S.; Gardberg, A.S.; Van Voorhis, W.C.; Staker, B.L.; Myler, P.J.; Edwards, T.E. Structures of a histidine triad family protein from Entamoeba histolytica bound to sulfate, AMP and GMP. Acta Crystallogr. F Struct. Biol. Commun. 2015, 71, 572–576. [Google Scholar]
- Lee, Y.N.; Nechushtan, H.; Figov, N.; Razin, E. The function of lysyl-tRNA synthetase and Ap4A as signaling regulators of MITF activity in FcεRI-activated mast cells. Immunity 2004, 20, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.N.; Razin, E. Nonconventional involvement of LysRS in the molecular mechanism of USF2 transcriptional activity in FcεRI-activated mast cells. Mol. Cell Biol. 2005, 25, 8904–8912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiske, J.; Huber, O. The histidine triad protein Hint1 triggers apoptosis independent of its enzymatic activity. J. Biol. Chem. 2006, 281, 27356–27366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cen, B.; Li, H.; Weinstein, I.B. Histidine triad nucleotide-binding protein 1 up-regulates cellular levels of p27KIP1 by targeting ScfSKP2 ubiquitin ligase and Src. J. Biol. Chem. 2009, 284, 5265–5276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Li, H.; Zhang, Y.; Santella, R.M.; Weinstein, I.B. HINT1 inhibits beta-catenin/TCF4, USF2 and NFkappaB activity in human hepatoma cells. Int. J. Cancer 2009, 124, 1526–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiske, J.; Huber, O. The histidine triad protein Hint1 interacts with Pontin and Reptin and inhibits TCF-beta-catenin-mediated transcription. J. Cell Sci. 2005, 118, 3117–3129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, T.; Suzui, M.; Wang, L.; Lin, C.S.; Xing, W.Q.; Weinstein, I.B. Deletion of histidine triad nucleotide-binding protein1/PKC-interacting protein in mice enhances cell growth and carcinogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 7824–7829. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Y.; Su, T.; Santella, R.M.; Weinstein, I.B. Hint1 is a haplo-insufficient tumor suppressor in mice. Oncogene 2006, 25, 713–721. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, Y.; Li, H.; Xu, Z.; Santella, R.M.; Weinstein, I.B. Hint1 inhibits growth and activator protein-1 activity in human colon cancer cells. Cancer Res. 2007, 67, 4700–4708. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, S.Y.; Hsu, C.Y.; He, J.R.; Liu, C.L.; Lo, S.J.; Chen, Y.C.; Huang, H.H. Identifying apoptosis-evasion proteins/pathways in human hepatoma cells via induction of cellular hormesis by UV irradiation. J. Proteome Res. 2009, 8, 3977–3986. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Li, H.; Wu, H.-C.; Shen, J.; Wang, L.; Yu, M.-W.; Lee, P.-H.; Weinstein, I.B.; Santella, R.M. Silencing of Hint1, a novel tumor suppressor gene, by promoter hypermetylation in hepatocellular carcinoma. Cancer Lett. 2009, 275, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Zambelli, D.; Zuntini, M.; Nardi, F.; Manara, M.C.; Serra, M.; Landuzzi, L.; Lollini, P.L.; Ferrari, S.; Alberghini, M.; Llombart-Bosch, A.; et al. Biological indicators of prognosis in Ewing’s sarcoma: An emerging role for lectin galactoside-binding soluble 3 binding protein (LGALS3BP). Int. J. Cancer 2010, 126, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Zuk, K.; Peczek, L.; Stec-Michalska, K.; Medrek, M.; Nawrot, B. Family history of gastric cancer correlates with decreased expression of HINT1 tumor suppressor gene in gastric mucosa of dyspeptic patients. Oncol. Lett. 2012, 3, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Muñoz, M.; Garzón, J. Nitric oxide and zinc-mediated protein assemblies involved in mu opioid receptor signaling. Mol Neurobiol. 2013, 48, 769–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente-Sánchez, A.; Sánchez-Blázquez, P.; Rodríguez-Muñoz, M.; Garzón, J. HINT1 protein cooperates with cannabinoid 1 receptor to negatively regulate glutamate NMDA receptor activity. Mol. Brain 2013, 6, 42. [Google Scholar] [CrossRef] [Green Version]
- Lima, C.D.; Klein, M.G.; Weinstein, I.B.; Hendrickson, W.A. Three-dimensional structure of human protein kinase C interacting protein 1, a member of the HIT family of proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 5357–5362. [Google Scholar] [CrossRef]
- Pearson, J.D.; DeWald, D.B.; Mathews, W.R.; Mozier, N.M.; Zürcher-Neely, H.A.; Heinrikson, R.L.; Morris, M.A.; McCubbin, W.D.; McDonald, J.R.; Fraser, E.D.; et al. Amino acid sequence and characterization of a protein inhibitor of protein kinase C. J. Biol. Chem. 1990, 265, 4583–4591. [Google Scholar] [CrossRef]
- Klein, M.G.; Yao, Y.; Slosberg, E.D.; Lima, C.D.; Doki, Y.; Weinstein, I.B. Characterization of PKCI and comparative studies with FHIT, related members of the HIT protein family. Exp. Cell Res. 1998, 244, 26–32. [Google Scholar] [CrossRef]
- Brenner, C.; Garrison, P.; Gilmour, J.; Peisach, D.; Ringe, D.; Petsko, G.A.; Lowenstein, J.M. Crystal structures of HINT1 demonstrate that histidine triad proteins are GalT-related nucleotide-binding proteins. Nat. Struct. Biol. 1997, 4, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Lima, C.D.; Klein, M.G.; Hendrickson, W.A. Structure-based analysis of catalysis and substrate definition in the HIT protein family. Science 1997, 278, 286–290. [Google Scholar] [CrossRef]
- Wang, J.; Fang, P.; Schimmel, P.; Guo, M. Side chains independent recognition of aminoacyl adenylates by the Hint1 transcription suppressor. J. Phys. Chem. B 2012, 116, 6798–6805. [Google Scholar] [CrossRef] [PubMed]
- Dolot, R.; Ozga, M.; Włodarczyk, A.; Krakowiak, A.; Nawrot, B. A new crystal form of human histidine triad nucleotide-binding protein 1 (hHINT1) in complex with adenosine 5′-monophosphate at 1.38 Å resolution. Acta Crystallogr. Sect. F Struct. Cryst. Commun. 2012, 68, 883–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krakowiak, A.; Pace, H.C.; Blackburn, M.; Adams, M.; Mekhalfia, A.; Kaczmarek, R.; Baraniak, J.; Stec, W.J.; Brenner, C. Biochemical, crystallographic, and mutagenic characterization of Hint1, the AMP-lysine hydrolase, with novel substrates and inhibitors. J. Biol. Chem. 2004, 279, 18711–18716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolot, R.; Ozga, M.; Krakowiak, A.; Nawrot, B. High-resolution X-ray structure of the rabbit histidine triad nucleotide-binding protein 1 (rHINT1)-adenosine complex at 1.10 Å resolution. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 601–607. [Google Scholar] [CrossRef]
- Yu, J.; Liu, Z.; Liang, Y.; Luo, F.; Zhang, J.; Tian, C.; Motzik, A.; Zheng, M.; Kang, J.; Zhong, G.; et al. Second messenger Ap4A polymerizes target protein HINT1 to transduce signals in FcεRI-activated mast cell. Nat. Commun. 2019, 10, 4664. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.-F.; Wagner, C.R. Lysyl-tRNA synthetase-generated lysyl-adenylate is a substrate for histidine triad nucleotide binding proteins. J. Biol. Chem. 2007, 282, 4719–4727. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.-F.; Tikh, I.B.; Horta, B.A.; Ghosh, B.; De Alencastro, R.B.; Wagner, C.R. Engineered monomeric human histidine triad nucleotide-binding protein 1 hydrolyzes fluorogenic acyl-adenylate and lysyl-tRNA synthetase-generated lysyl-adenylate. J. Biol. Chem. 2007, 282, 15137–15147. [Google Scholar] [CrossRef] [Green Version]
- Murakami, E.; Tolstykh, T.; Bao, H.; Niu, C.; Steuer, H.M.; Bao, D.; Chang, W.; Espiritu, C.; Bansal, S.; Lam, A.M.; et al. Mechanism of activation of PSI-7851 and its diastereoisomer PSI-7977. J. Biol. Chem. 2010, 285, 34337–34347. [Google Scholar] [CrossRef] [Green Version]
- Krakowiak, A.; Kaczmarek, R.; Baraniak, J.; Wieczorek, M.; Stec, W.J. Stereochemistry of rHint1 hydrolase assisted cleavage of P-N bond in nucleoside 5′-O-phosphoramidothioates. Chem. Commun. 2007, 21, 2163–2165. [Google Scholar] [CrossRef]
- Gilmour, J.; Liang, N.; Lowenstein, J.M. Isolation, cloning and characterization of a low-molecular -mass purine nucleoside- and nucleotide-binding-protein. Biochem. J. 1997, 326, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Mueller, U.; Darowski, N.; Fuchs, M.R.; Förster, R.; Hellmig, M.; Paithankar, K.S.; Pühringer, S.; Steffien, M.; Zocher, G.; Weiss, M.S. Facilities for macromolecular crystallography at the Helmholtz-Zentrum Berlin. J. Synchrotron Radiat. 2012, 19, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W. Integration, scaling, space-group assignment and post-refinement. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagin, A.; Teplyakov, A. Molecular replacement with MOLREP. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 22–25. [Google Scholar] [CrossRef]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. The PyMOL Molecular Graphics System; DeLano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Maize, K.M.; Shah, R.; Strom, A.; Kumarapperuma, S.; Zhou, A.; Wagner, C.R.; Finzel, B.C. A crystal structure based guide to the design of human histidine triad nucleotide binding protein (hHINT1) activated ProTides. Mol. Pharm. 2017, 14, 3987–3997. [Google Scholar] [CrossRef]
- Kabsch, W.; Sander, C. Dictionary of protein secondary structure: Pattern recognition of hydrogen-bonded and geometrical features. Biopolymers 1983, 22, 2577–2637. [Google Scholar] [CrossRef]
- Varnum, J.M.; Baraniak, J.; Kaczmarek, R.; Stec, W.J.; Brenner, C. Di-, tri- and tetra-5′-O-phosphorothioadenosyl substituted polyols as inhibitors of Fhit: Importance of the alpha-beta bridging oxygen and beta phosphorus replacement. BMC Chem. Biol. 2001, 1, 3. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PDB Entry | 6g9z |
|---|---|
| Space group | P212121 |
| Wavelength (Å) | 0.96690 |
| Unit-cell parameters | |
| a (Å) | 46.08 |
| b (Å) | 63.38 |
| c (Å) | 76.84 |
| Total no. of reflections | 154,268 |
| Unique reflections | 40,954 |
| Resolution (Å) | 39.52–1.43 (1.51–1.43) |
| Completeness (%) | 96.1 (92.7) |
| CC(1/2) (%) | 99.4 (83.3) |
| Rmergea (%) | 13.7 (70.3) |
| Redundancy | 3.77 |
| Mosaicity (°) | 0.21 |
| Wilson B factor (Å2) | 15.0 |
| Mean I/σ(I) | 8.78 (1.96) |
| Refinement statistics | |
| No. reflections used in refinement | 38,759 |
| No. reflections used to Rfree | 2041 |
| Rcryst (Rfree) b | 0.224/0.277 |
| No. non-H atoms | |
| Protein | 1998 |
| Solvent | 401 |
| Ligands | 9 (malate ion) |
| R.m.s.d. from ideal values | |
| Bond lengths (Å) | 0.016 |
| Bond angles (°) | 1.812 |
| Ramachandran plot c | |
| Favored (%) | 98.4 |
| Allowed (%) | 1.2 |
| Outliers (%) | 0.4 |
| Mean B values d | |
| Protein (Å2) | 8.53 |
| Malate ion (Å2) | 14.26 |
| Water (Å2) | 25.53 |
| Space Group | PDB ID | Cell Dimensions a, b, c [Å], α, β, γ [°] | Matthews Coefficient [Å3 Da−1] | Solvent Content [%] |
|---|---|---|---|---|
| Triclinic | ||||
| P1 | 6j5z | 45.42 45.44 63.92 86.57 86.48 61.35 | 2.07 | 40.54 |
| Monoclinic | ||||
| P21 | 4zkl | 63.71 46.42 103.43 90 97.41 90 | 3.03 | 59.45 |
| P21 | 4zkv | 46.21 79.00 63.88 90 90.10 90 | 2.27 | 45.92 |
| P21 | 5km0 | 64.33 89.59 46.37 90 90.03 90 | 2.37 | 48.10 |
| C2 | 3tw2 | 77.42 46.45 64.03 90 94.42 90 | 2.08 | 40.75 |
| Orthorhombic | ||||
| P212121 | 5klz | 45.83 75.92 80.79 90 90 90 | 2.49 | 50.66 |
| Tetragonal | ||||
| P41 | 7q2u | 112.89 112.89 43.63 90 90 90 | 2.52 | 51.18 |
| P43212 | 6b42 | 39.74 39.74 141.13 90 90 90 | 1.98 | 37.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dolot, R.; Mikołajczyk, A.; Nawrot, B. Crystal Packing Differences as a Key Factor for Stabilization of the N-Terminal Fragment of the Human HINT1 Protein. Crystals 2023, 13, 1197. https://doi.org/10.3390/cryst13081197
Dolot R, Mikołajczyk A, Nawrot B. Crystal Packing Differences as a Key Factor for Stabilization of the N-Terminal Fragment of the Human HINT1 Protein. Crystals. 2023; 13(8):1197. https://doi.org/10.3390/cryst13081197
Chicago/Turabian StyleDolot, Rafał, Aleksandra Mikołajczyk, and Barbara Nawrot. 2023. "Crystal Packing Differences as a Key Factor for Stabilization of the N-Terminal Fragment of the Human HINT1 Protein" Crystals 13, no. 8: 1197. https://doi.org/10.3390/cryst13081197
APA StyleDolot, R., Mikołajczyk, A., & Nawrot, B. (2023). Crystal Packing Differences as a Key Factor for Stabilization of the N-Terminal Fragment of the Human HINT1 Protein. Crystals, 13(8), 1197. https://doi.org/10.3390/cryst13081197