Structural Analyses of Helicobacter Pylori FolC Conducting Glutamation in Folate Metabolism

 , ,
, , 
Abstract
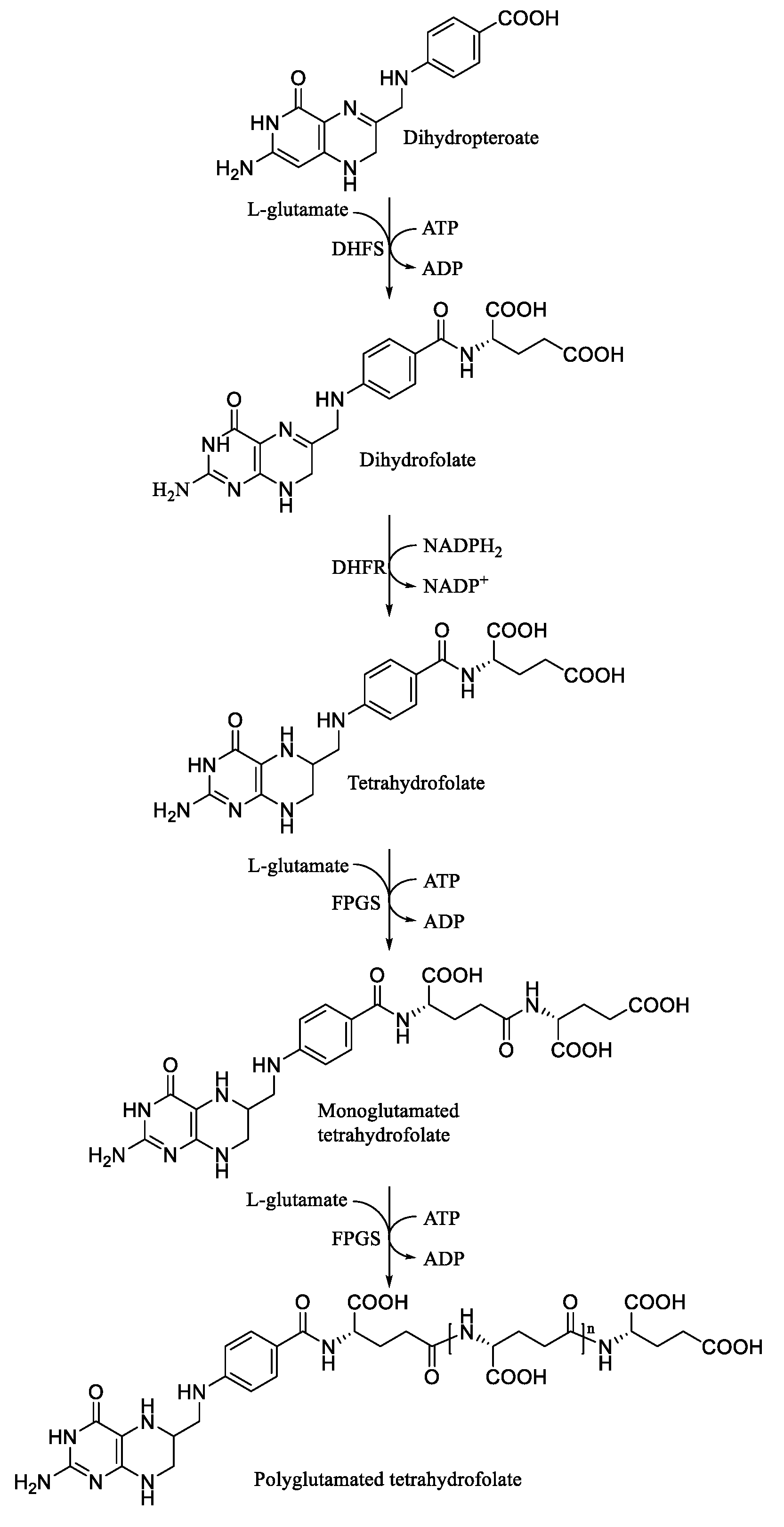
1. Introduction
2. Materials and Methods
2.1. Cloning, Protein Expression, and Purification
2.2. Crystallography
2.3. Data Availibility
3. Results and Discussions
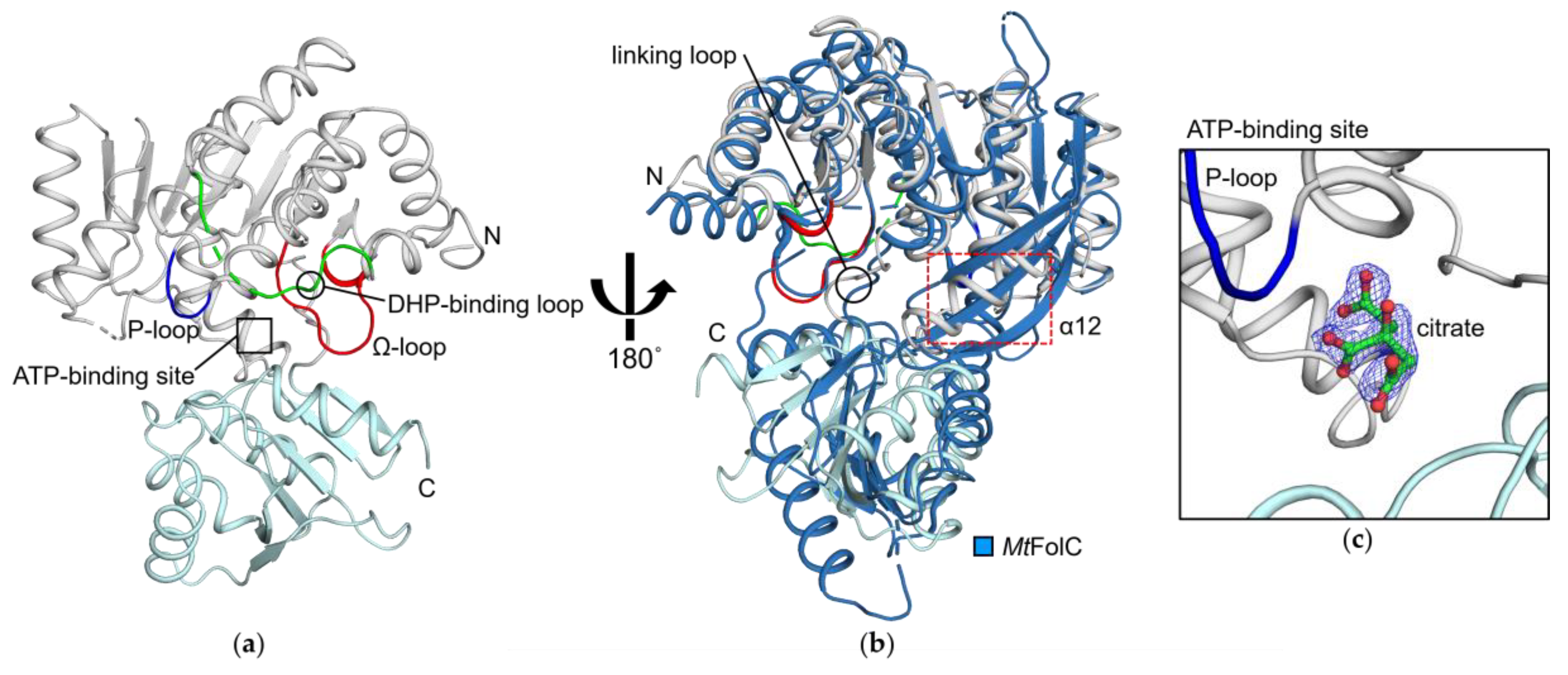
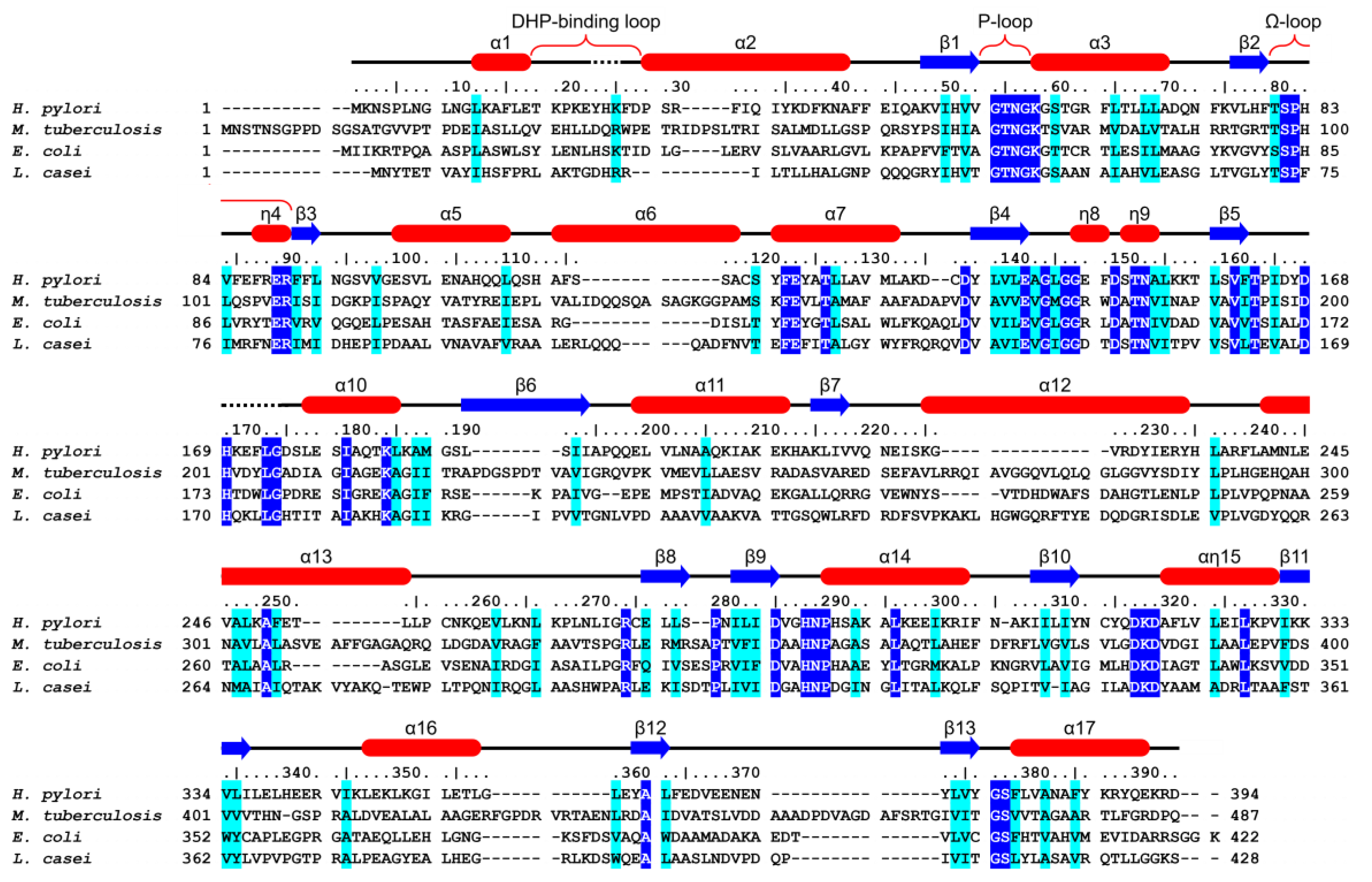
3.1. HpFolC Shares Structural Features of Other FolC/FPGSs
3.2. Binding Site of HpFolC for DHP in the Vicinity of the DHP-Binding Loop
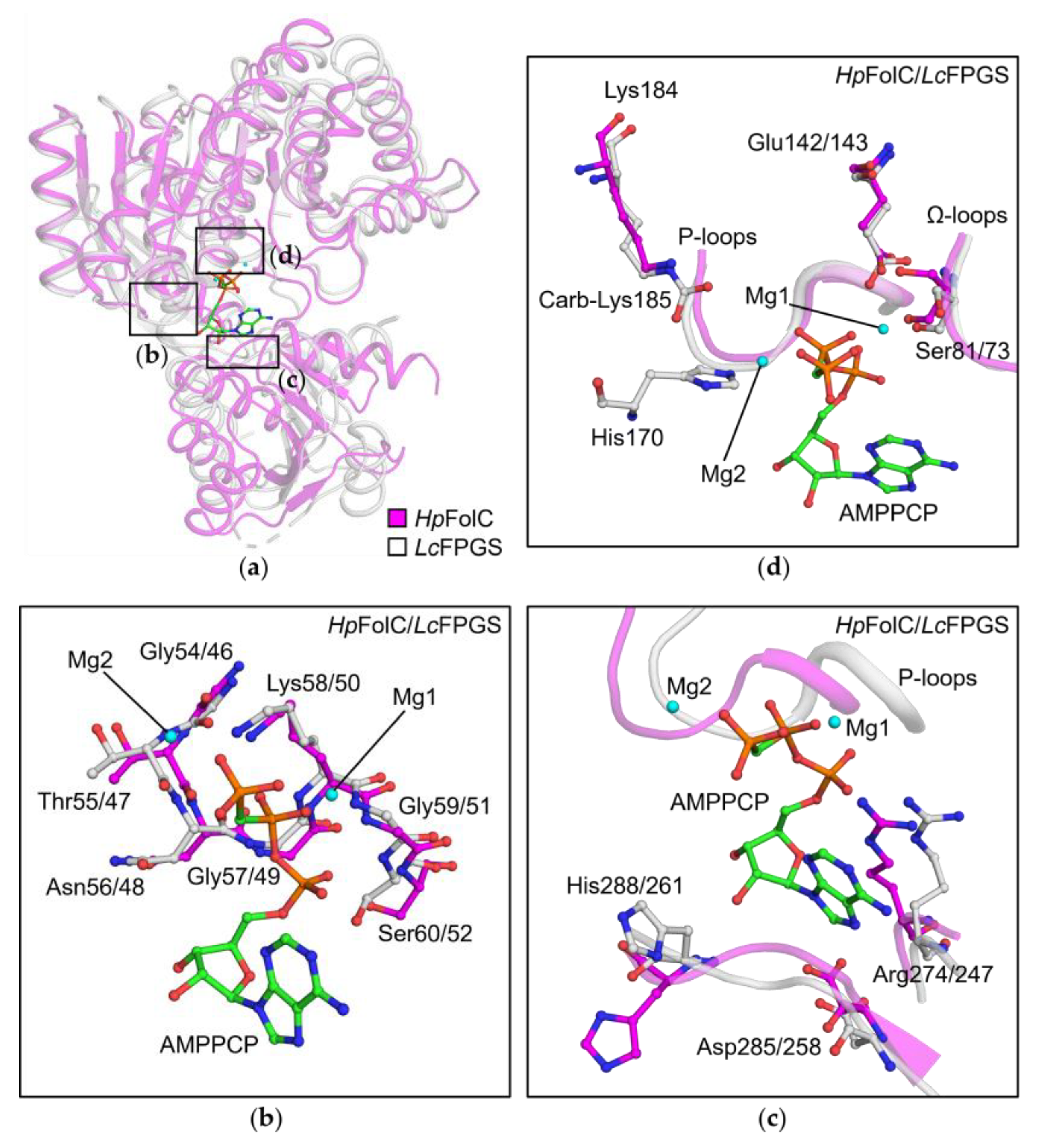
3.3. P-Loop, a Structural Motif of HpFolC for ATP Binding
3.4. Structural Motifs of HpFolC for ATP Binding via Two Magnesium Ions
3.5. Analyses of Folate Binding of HpFolC, Focused on the Ω-Loop
3.6. l-Glu Binding Site of FolC/FPGSs
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Warren, J.R.; Marshall, B. Unidentified curved bacilli on gastric epithelium in active chronic gastritis. Lancet 1983, 1, 1273–1275. [Google Scholar] [PubMed]
- Marshall, B.J.; Warren, J.R. Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet 1984, 1, 1311–1315. [Google Scholar] [CrossRef]
- Harguindey, S.; Pedraz, J.L.; Garcia Canero, R.; Perez de Diego, J.; Cragoe, E.J., Jr. Hydrogen ion-dependent oncogenesis and parallel new avenues to cancer prevention and treatment using a H(+)-mediated unifying approach: pH-related and pH-unrelated mechanisms. Crit. Rev. Oncog. 1995, 6, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Venerito, M.; Schulz, C. Helicobacter pylori Infection: New Facts in Clinical Management. Curr. Treat. Options Gastroenterol. 2018, 16, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet. Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Shane, B. Folylpolyglutamate synthesis and role in the regulation of one-carbon metabolism. Vitam. Horm. 1989, 45, 263–335. [Google Scholar] [PubMed]
- Lu, Y.Z.; Aiello, P.D.; Matthews, R.G. Studies on the polyglutamate specificity of thymidylate synthase from fetal pig liver. Biochemistry 1984, 23, 6870–6876. [Google Scholar] [CrossRef] [PubMed]
- Schirch, V.; Strong, W.B. Interaction of folylpolyglutamates with enzymes in one-carbon metabolism. Arch. Biochem. Biophys. 1989, 269, 371–380. [Google Scholar] [CrossRef]
- Lowe, K.E.; Osborne, C.B.; Lin, B.F.; Kim, J.S.; Hsu, J.C.; Shane, B. Regulation of folate and one-carbon metabolism in mammalian cells. II. Effect of folylpoly-gamma-glutamate synthetase substrate specificity and level on folate metabolism and folylpoly-gamma-glutamate specificity of metabolic cycles of one-carbon metabolism. J. Biol. Chem. 1993, 268, 21665–21673. [Google Scholar]
- Synold, T.W.; Willits, E.M.; Barredo, J.C. Role of folylpolygutamate synthetase (FPGS) in antifolate chemotherapy; a biochemical and clinical update. Leuk. Lymphoma 1996, 21, 9–15. [Google Scholar] [CrossRef]
- Shane, B. Pteroylpoly(gamma-glutamate) synthesis by Corynebacterium species. In vivo synthesis of folates. J. Biol. Chem. 1980, 255, 5649–5654. [Google Scholar] [PubMed]
- Bognar, A.L.; Osborne, C.; Shane, B.; Singer, S.C.; Ferone, R. Folylpoly-gamma-glutamate synthetase-dihydrofolate synthetase. Cloning and high expression of the Escherichia coli folC gene and purification and properties of the gene product. J Biol. Chem. 1985, 260, 5625–5630. [Google Scholar]
- Fussenegger, M.; Meyer, T.F. Cloning and characterization of the Neisseria gonorrhoeae MS11 folC gene. Mol. Gen. Genet. 1996, 250, 277–285. [Google Scholar] [PubMed]
- Sassetti, C.M.; Boyd, D.H.; Rubin, E.J. Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol. 2003, 48, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, Q.; Yang, Y.; Coward, J.K.; Nzila, A.; Sims, P.F.; Hyde, J.E. Characterisation of the bifunctional dihydrofolate synthase-folylpolyglutamate synthase from Plasmodium falciparum; a potential novel target for antimalarial antifolate inhibition. Mol. Biochem. Parasitol. 2010, 172, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, P.; Livstone, M.S.; Lewis, S.E.; Thomas, P.D. Phylogenetic-based propagation of functional annotations within the Gene Ontology consortium. Brief. Bioinform. 2011, 12, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Bourne, C.R. Utility of the Biosynthetic Folate Pathway for Targets in Antimicrobial Discovery. Antibiotics (Basel) 2014, 3, 1–28. [Google Scholar] [CrossRef]
- Mathieu, M.; Debousker, G.; Vincent, S.; Viviani, F.; Bamas-Jacques, N.; Mikol, V. Escherichia coli FolC structure reveals an unexpected dihydrofolate binding site providing an attractive target for anti-microbial therapy. J. Biol. Chem. 2005, 280, 18916–18922. [Google Scholar] [CrossRef]
- Sun, X.; Bognar, A.L.; Baker, E.N.; Smith, C.A. Structural homologies with ATP- and folate-binding enzymes in the crystal structure of folylpolyglutamate synthetase. Proc. Natl. Acad. Sci. USA 1998, 95, 6647–6652. [Google Scholar] [CrossRef]
- Sun, X.; Cross, J.A.; Bognar, A.L.; Baker, E.N.; Smith, C.A. Folate-binding triggers the activation of folylpolyglutamate synthetase. J. Mol. Biol. 2001, 310, 1067–1078. [Google Scholar] [CrossRef]
- Smith, C.A.; Cross, J.A.; Bognar, A.L.; Sun, X. Mutation of Gly51 to serine in the P-loop of Lactobacillus casei folylpolyglutamate synthetase abolishes activity by altering the conformation of two adjacent loops. Acta Crystallogr. D Biol. Crystallogr. 2006, 62, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Young, P.G.; Smith, C.A.; Metcalf, P.; Baker, E.N. Structures of Mycobacterium tuberculosis folylpolyglutamate synthase complexed with ADP and AMPPCP. Acta Crystallogr. D Biol. Crystallogr. 2008, D64, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar] [PubMed]
- Terwilliger, T.C. SOLVE and RESOLVE: Automated structure solution and density modification. Methods Enzymol. 2003, 374, 22–37. [Google Scholar] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Cystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.B.; Arendall, W.B., 3rd; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, J.A.; Auger, G.; Fanchon, E.; Martin, L.; Blanot, D.; van Heijenoort, J.; Dideberg, O. Crystal structure of UDP-N-acetylmuramoyl-L-alanine:D-glutamate ligase from Escherichia coli. EMBO J. 1997, 16, 3416–3425. [Google Scholar] [CrossRef]
- Bertrand, J.A.; Auger, G.; Martin, L.; Fanchon, E.; Blanot, D.; Le Beller, D.; van Heijenoort, J.; Dideberg, O. Determination of the MurD mechanism through crystallographic analysis of enzyme complexes. J. Mol. Biol. 1999, 289, 579–590. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HpFolC | SeMet-substituted HpFolC | |
|---|---|---|
| PDB code | 6K8C | |
| Data collection | ||
| Beam line | PLS-5C | |
| Space group | C2 | P41212 |
| Wavelength (Å) | 0.9795 | 0.9798 (Peak) |
| Unit cell parameter | ||
| a, b, c (Å) | 131.45, 61.64, 69.74 | 117.90, 117.90, 69.18 |
| α, β, γ (°) | 90, 115.96, 90 | 90, 90, 90 |
| Resolution range (Å) | 50.00–1.95 (1.98–1.95)1 | 50.00–2.50 (2.54–2.50)1 |
| No. of unique reflections | 36178 (3591)1 | 17401 (1701)1 |
| I/σ(I) | 26.5 (4.2)1 | 69.2 (15.9)1 |
| Wilson B-factor (Å2) | 33.9 | 28.4 |
| Rmerge (%) | 0.059 (0.431)1 | 0.148 (0.635)1 |
| Redundancy | 7.4 (7.5)1 | 38.1 (39.9)1 |
| Completeness (%) | 98.9 (98.9)1 | 100.0 (100.0)1 |
| Refinement | ||
| Resolution | 30.00–1.95 (2.02–1.95)1 | |
| Rwork/Rfree2 | 0.184/0.219 | |
| R.m.s. deviations | ||
| Bonds (Å) | 0.004 | |
| Angles (°) | 0.640 | |
| No. of non-H atoms | 3362 | |
| Protein | 3029 | |
| Ligand | 193 | |
| Water | 314 | |
| Average B-factors (Å2) | 41.22 | |
| Protein | 40.42 | |
| Ligand | 62.68 | |
| Water | 47.59 | |
| Ramachandran favored/outlier (%) | 98.4/0.04 | |
| Rotamer outlier (%) | 0.00 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.S.; Kim, H.S.; Park, S.H.; Park, M.S.; Kang, S.-M.; Kim, H.-J.; Han, B.W. Structural Analyses of Helicobacter Pylori FolC Conducting Glutamation in Folate Metabolism. Crystals 2019, 9, 429. https://doi.org/10.3390/cryst9080429
Park JS, Kim HS, Park SH, Park MS, Kang S-M, Kim H-J, Han BW. Structural Analyses of Helicobacter Pylori FolC Conducting Glutamation in Folate Metabolism. Crystals. 2019; 9(8):429. https://doi.org/10.3390/cryst9080429
Chicago/Turabian StylePark, Joon Sung, Hyoun Sook Kim, Sang Ho Park, Mi Seul Park, Sung-Min Kang, Hyun-Jung Kim, and Byung Woo Han. 2019. "Structural Analyses of Helicobacter Pylori FolC Conducting Glutamation in Folate Metabolism" Crystals 9, no. 8: 429. https://doi.org/10.3390/cryst9080429
APA StylePark, J. S., Kim, H. S., Park, S. H., Park, M. S., Kang, S.-M., Kim, H.-J., & Han, B. W. (2019). Structural Analyses of Helicobacter Pylori FolC Conducting Glutamation in Folate Metabolism. Crystals, 9(8), 429. https://doi.org/10.3390/cryst9080429