
Immunophenotypical Characterization of M1/M2 Macrophages and Lymphocytes in Cisplatin-Induced Rat Progressive Renal Fibrosis


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Procedures
2.2. Histopathology and Immunohistochemistry
2.3. Double Immunofluorescence
2.4. Reverse-Transcription Polymerase Chain Reaction (RT–PCR)
2.5. Statistical Analysis
3. Results
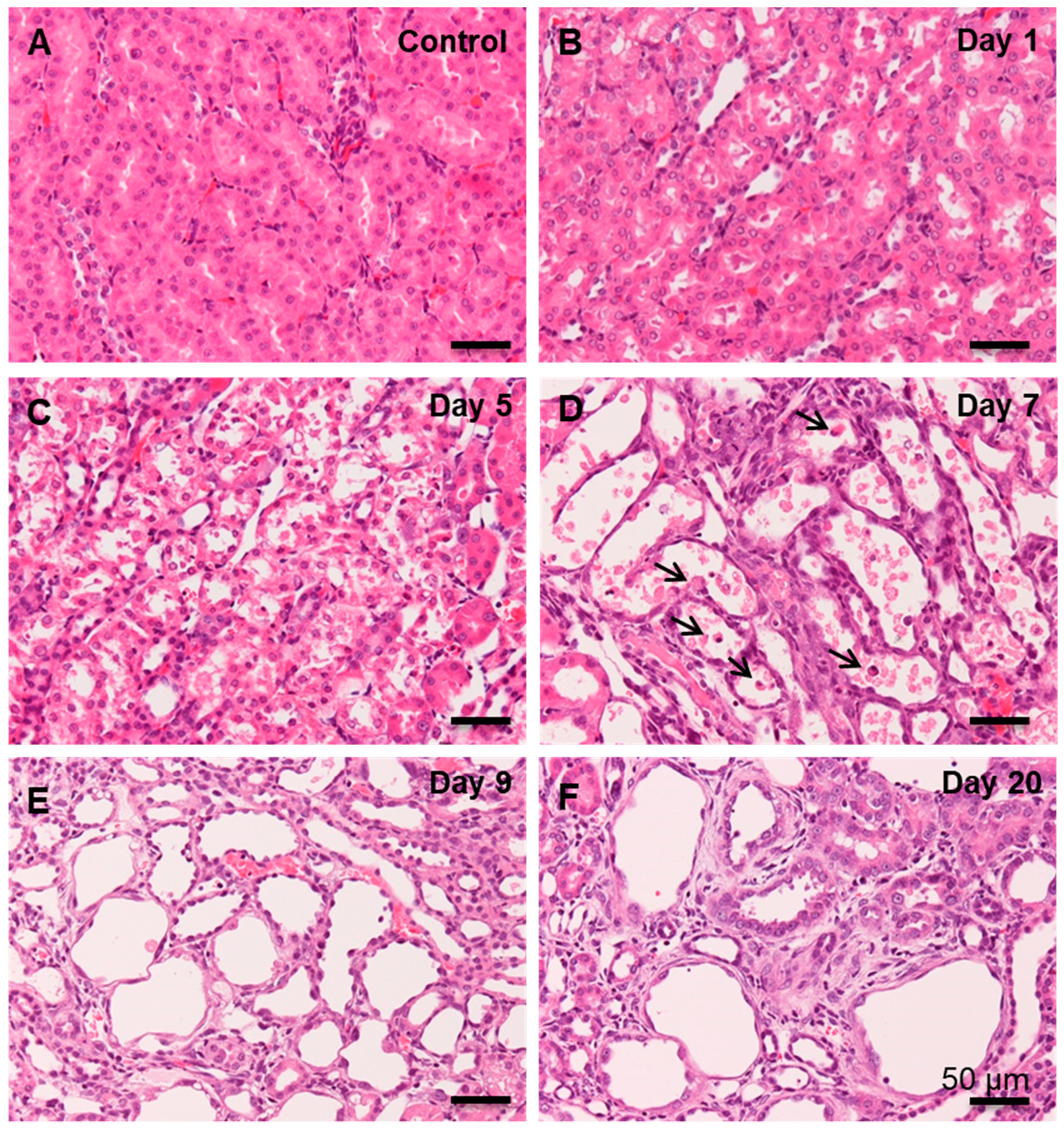
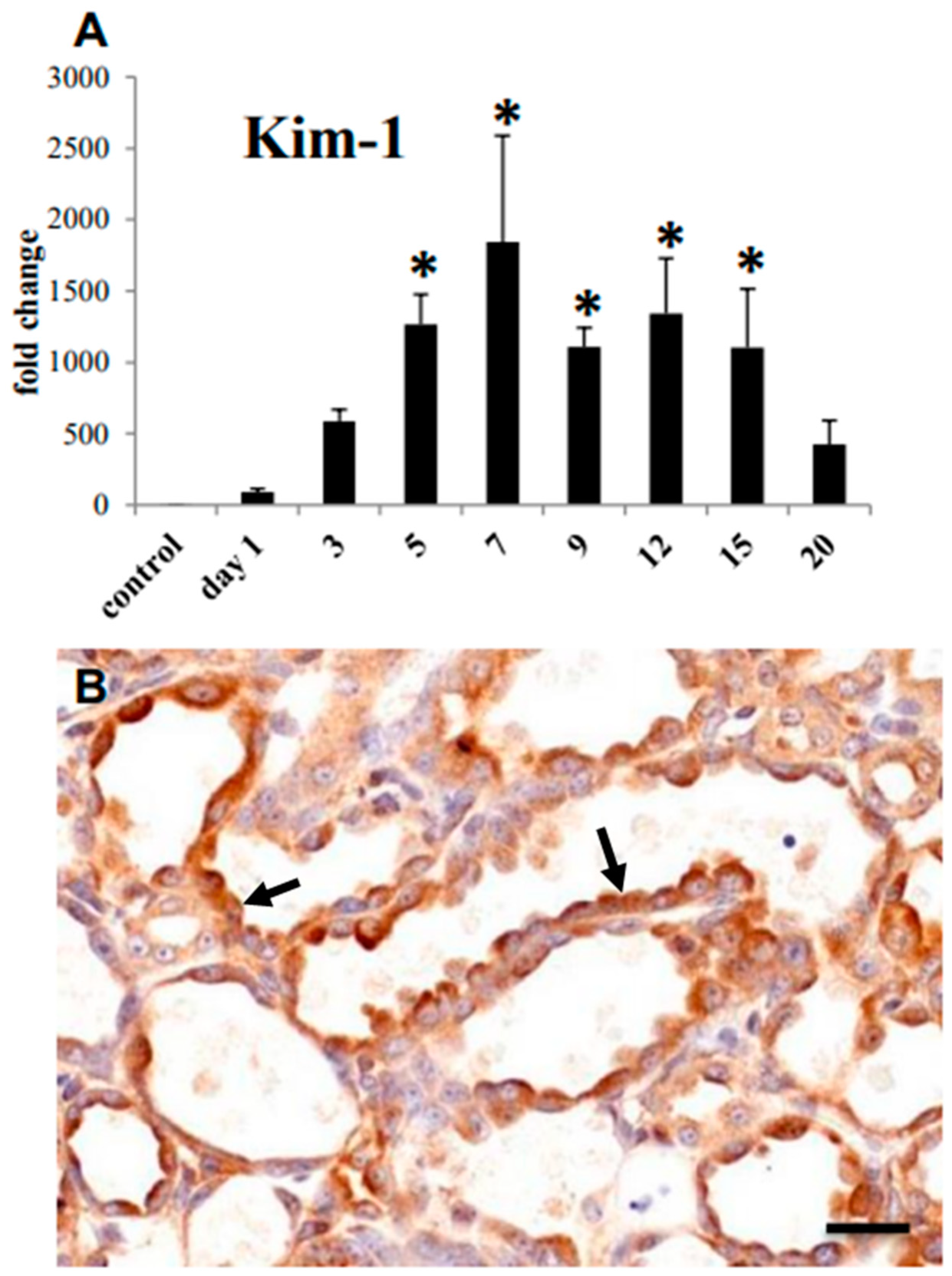
3.1. Histopathology of CDDP-Induced Rat Renal Lesion
3.2. Collagen Deposition and Myofibroblast Appearance in CDDP-Induced Rat Renal Lesion
3.3. Macrophages Appearing in CDDP-Induced Rat Renal Lesion
3.3.1. CD68+ Macrophages
3.3.2. CD163+ Macrophages
3.3.3. MHC Class II+ Macrophages
3.3.4. CD204+ Macrophages
3.4. M1/M2 Macrophage Polarization in CDDP-Induced Rat Renal Lesion
3.5. M1- and M2-Related Factors in CDDP-Induced Rat Renal Lesion
3.6. Lymphocyte Appearance in CDDP-Induced Rat Renal Lesion
3.6.1. CD4+ T Lymphocytes
3.6.2. CD8+ T Lymphocytes
3.6.3. CD20+ B Lymphocytes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ricardo, S.D.; van Goor, H.; Eddy, A.A. Macrophage diversity in renal injury and repair. J. Clin. Investig. 2008, 118, 3522–3530. [Google Scholar] [CrossRef] [Green Version]
- Henderson, N.C.; Mackinnon, A.C.; Farnworth, S.L.; Kipari, T.; Haslett, C.; Iredale, J.P.; Liu, F.T.; Hughes, J.; Sethi, T. Galectin-3 expression and secretion links macrophages to the promotion of renal fibrosis. Am. J. Pathol. 2008, 172, 288–298. [Google Scholar] [CrossRef] [Green Version]
- LeBleu, V.S.; Taduri, G.; O’Connell, J.; Teng, Y.; Cooke, V.G.; Woda, C.; Sugimoto, H.; Kalluri, R. Origin and function of myofibroblasts in kidney fibrosis. Nat. Med. 2013, 19, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Fadok, V.A.; Bratton, D.L.; Konowal, A.; Freed, P.W.; Westcott, J.Y.; Henson, P.M. Macrophages that have ingested apoptotic cells in vitro inhibit proinflammatory cytokine production through autocrine/paracrine mechanisms involving TGF-beta, PGE2, and PAF. J. Clin. Investig. 1998, 101, 890–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Border, W.A.; Noble, N.A. Evidence that TGF-beta should be a therapeutic target in diabetic nephropathy. Kidney Int. 1998, 54, 1390–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Qu, X.; Yao, J.; Caruana, G.; Ricardo, S.D.; Yamamoto, Y.; Yamamoto, H.; Bertram, J.F. Blockade of endothelial-mesenchymal transition by a Smad3 inhibitor delays the early development of streptozotocin-induced diabetic nephropathy. Diabetes 2010, 59, 2612–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, H.J.; Ryu, M. Renal microenvironments and macrophage phenotypes determine progression or resolution of renal inflammation and fibrosis. Kidney Int. 2011, 80, 915–925. [Google Scholar] [CrossRef] [Green Version]
- Lech, M.; Anders, H.J. Macrophages and fibrosis: How resident and infiltrating mononuclear phagocytes orchestrate all phases of tissue injury and repair. Biochim. Biophys. Acta 2013, 1832, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Nahrendorf, M.; Swirski, F.K. Abandoning M1/M2 for a network model ofmacrophage function. Circ. Res. 2016, 119, 414–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damoiseaux, J.G.; Döpp, E.A.; Calame, W.; Chao, D.; MacPherson, G.G.; Dijkstra, C.D. Rat macrophage lysosomal membrane antigen recognized by monoclonal antibody ED1. Immunology 1994, 83, 140–147. [Google Scholar] [PubMed]
- Sisino, G.; Bouckenooghe, T.; Aurientis, S.; Fontaine, P.; Storme, L.; Vambergue, A. Diabetes during pregnancy influences Hofbauer cells, a subtype of placental macrophages, to acquire a pro-inflammatory phenotype. Biochim. Biophys. Acta 2013, 1832, 1959–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijesundera, K.K.; Izawa, T.; Murakami, H.; Tennakoon, A.H.; Golbar, M.H.; Ichikawa, C.H.; Tanaka, M.; Kuwamura, M.; Yamate, J. M1- and M2-macrophage polarization in thioacetamide (TAA)-induced rat liver lesions; a possible analysis for hepato-pathology. Histol. Histopathol. 2014, 29, 497–511. [Google Scholar] [PubMed]
- Polfliet, M.M.; Fabriek, B.O.; Daniëls, W.P.; Dijkstra, C.D.; van den Berg, T.K. The rat macrophage scavenger receptor CD163: Expression, regulation and role in inflammatory mediator production. Immunobiology 2006, 211, 419–425. [Google Scholar] [CrossRef]
- Yamate, J.; Sato, K.; Ide, M.; Nakanishi, M.; Kuwamura, M.; Sakuma, S.; Nakatsuji, S. Participation of different macrophage populations and myofibroblasticcells in chronically developed renal interstitial fibrosis after cisplatin-induced canal injury in rats. Vet. Pathol. 2002, 39, 322–333. [Google Scholar] [CrossRef]
- Yamate, J.; Machida, Y.; Ide, M.; Kuwamura, M.; Sawamoto, O.; LaMarre, J. Effects of lipopolysaccharide on the appearance of macrophage populations and fibrogenesis in cisplatin-induced rat renal injury. J. Exp. Toxicol. Pathol. 2004, 56, 13–24. [Google Scholar] [CrossRef]
- Pervin, M.; Golbar, H.M.; Bondoc, A.; Izawa, T.; Kuwamura, M.; Yamate, J. Immunophenotypical characterization and influence on liver homeostasis of depleting and repopulating hepatic macrophages in rats injected with clodronate. Exp. Toxicol. Pathol. 2016, 68, 113–124. [Google Scholar] [CrossRef]
- Yamamoto, E.; Izawa, T.; Juniantito, V.; Kuwamura, M.; Sugiura, K.; Takeuchi, T.; Yamate, J. Involvement of endogenous prostaglandin E2 in tubular epithelial regeneration through inhibition of apoptosis and epithelial-mesenchymal transition in cisplatin-induced rat renal lesions. Histol. Histopathol. 2010, 25, 995–1007. [Google Scholar]
- Yuasa, T.; Juniantito, V.; Ichikawa, C.; Yano, R.; Izawa, T.; Kuwamura, M.; Yamate, J. Thy-1 expression, a possible marker of early myofibroblast development, in renal tubulointerstitial fibrosis induced in rats by cisplatin. Exp. Toxicol. Pathol. 2013, 65, 651–659. [Google Scholar] [CrossRef]
- Han, W.K.; Bailly, V.; Abichandani, R.; Thadhani, R.; Bonventre, J.V. Kidney Injury Molecule-1 (KIM-1): A novel biomarker for human renal proximal tubule injury. Kidney Int. 2002, 62, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Timmeren, M.M.; van den Heuvel, M.C.; Bailly, V.; Bakker, S.J.; van Goor, H.; Stegeman, C.A. Tubular kidney injury molecule-1 (KIM-1) in human renal disease. J. Pathol. 2007, 212, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Terada, N.; Karim, M.R.; Izawa, T.; Kuwamura, M.; Yamate, J. Expression of β-catenin in regenerating renal tubules of cisplatin-induced kidney failure in rats. Clin. Exp. Nephrol. 2018, 22, 1240–1250. [Google Scholar] [CrossRef]
- Liu, Y. Cellular and molecular mechanisms of renal fibrosis. Nat. Rev. Nephrol. 2011, 7, 684–696. [Google Scholar] [CrossRef]
- Wang, Y.; Harris, D.C. Macrophages in renal disease. J. Am. Soc. Nephrol. 2011, 22, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Chen, X.; Zhang, Y.; George, J.; Cobbs, A.; Wang, G.; Li, L.; Emmett, N. Kidney injury molecule-1 is upregulated in renal lipotoxicity and mediates palmitate-induced tubular cell injury and inflammatory response. Int. J. Mol. Sci. 2019, 20, 3406. [Google Scholar] [CrossRef] [Green Version]
- Rahman, N.; Pervin, M.; Kuramochi, M.; Karim, M.R.; Izawa, T.; Kuwamura, M.; Yamate, J. M1/M2-macrophage polarization-based hepatotoxicity in d-galactosamine-induced acute liver injury in rats. Toxicol. Pathol. 2018, 46, 764–776. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Komohara, Y.; Takaishi, K.; Katabuchi, H.; Takeya, M. Detection of M2 macrophages and colony-stimulating factor 1 expression in serous and mucinous ovarian epithelial tumors. Pathol. Int. 2009, 59, 300–305. [Google Scholar] [CrossRef]
- Liu, L.; Kou, P.; Zeng, Q.; Pei, G.; Li, Y.; Liang, H.; Xu, G.; Chen, S. CD4+ T Lymphocytes, especially Th2 cells, contribute to the progress of renal fibrosis. Am. J. Nephrol. 2012, 36, 386–396. [Google Scholar] [CrossRef]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Zhu, J.; Wang, Y.; Zhu, Z.; Chen, Y.; Lu, L.; Jin, W.; Yan, X.; Zhang, R. Renal recruitment of B lymphocytes exacerbates tubulointerstitial fibrosis by promoting monocyte mobilization and infiltration after unilateral ureteral obstruction. J. Pathol. 2017, 241, 80–90. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Type | Dilution | Pretreatment | Source |
|---|---|---|---|---|
| CD163 (ED2) | Mouse monoclonal | 1/300 | - | AbDSerotec, Oxford, UK |
| CD68 (ED1) | Mouse monoclonal | 1/500 | 10 µg/mL proteinase K, 20 min | Merck Millipore, MA, USA |
| MHC class II (OX-6) | Mouse monoclonal | 1/300 | Microwaving in citrate buffer, 20 min | AbDSerotec, Oxford, UK |
| CD204 (SRA-E5) | Mouse monoclonal | 1/500 | Microwaving in citrate buffer, 20 min | Transgenic Inc., Kumamoto, Japan |
| α-SMA | Mouse monoclonal | 1/1000 | - | Dako, CA, USA |
| CD4 | Mouse monoclonal | 1/2000 | - | AbDSerotec, Oxford, UK |
| CD8 | Mouse monoclonal | 1/200 | - | AbDSerotec, Oxford, UK |
| CD20 | Goat polyclonal | 1/200 | Microwaving in citrate buffer, 20 min | Santa Cruz Biotechnology Inc., TX, USA |
| KIM-1 | Goat polyclonal | 1/500 | - | Immunology Consultants Laboratory, OR, USA |
| Primer | NCBI Symbol | Forward | Reverse |
|---|---|---|---|
| IFN-γ | Ifng | tcgcacctgatcactaacttcttc | cgactccttttccgcttcc |
| TNF-α | Tnf | tgcctcagcctcttctcattc | gctcctctgcttggtggttt |
| TGF-β1 | Tgfb1 | cttcagctccacagagaagaactgc | cacgatcatgttggacaactgctcc |
| KIM-1 | Havcr1 | gtctgtattgttgccgagtgga | gttgtgggtcttgttggagga |
| β-actin | Actb | taaagacctctatgccaacac | ctcctgcttgctgatccacat |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakagawa, M.; Karim, M.R.; Izawa, T.; Kuwamura, M.; Yamate, J. Immunophenotypical Characterization of M1/M2 Macrophages and Lymphocytes in Cisplatin-Induced Rat Progressive Renal Fibrosis. Cells 2021, 10, 257. https://doi.org/10.3390/cells10020257
Nakagawa M, Karim MR, Izawa T, Kuwamura M, Yamate J. Immunophenotypical Characterization of M1/M2 Macrophages and Lymphocytes in Cisplatin-Induced Rat Progressive Renal Fibrosis. Cells. 2021; 10(2):257. https://doi.org/10.3390/cells10020257
Chicago/Turabian StyleNakagawa, Minto, Mohammad Rabiul Karim, Takeshi Izawa, Mitsuru Kuwamura, and Jyoji Yamate. 2021. "Immunophenotypical Characterization of M1/M2 Macrophages and Lymphocytes in Cisplatin-Induced Rat Progressive Renal Fibrosis" Cells 10, no. 2: 257. https://doi.org/10.3390/cells10020257