
Eukaryotic Translation Initiation Factor 4AI: A Potential Novel Target in Neuroblastoma

 ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Specimens
2.2. Cell Lines and Rocaglate CR-1-31-B
2.3. Statistical Analysis
2.4. Immunohistochemistry
2.5. Protein and RNA Isolation
2.6. Western Blot
2.7. Quantitative Real-Time PCR (RT-qPCR)
2.8. Survival Analysis
2.9. Cell Culture
2.10. Cell Viability
2.11. Cell Proliferation
2.12. Apoptosis Assays
2.13. Cell Cycle
2.14. Puromycin Labelling
3. Results
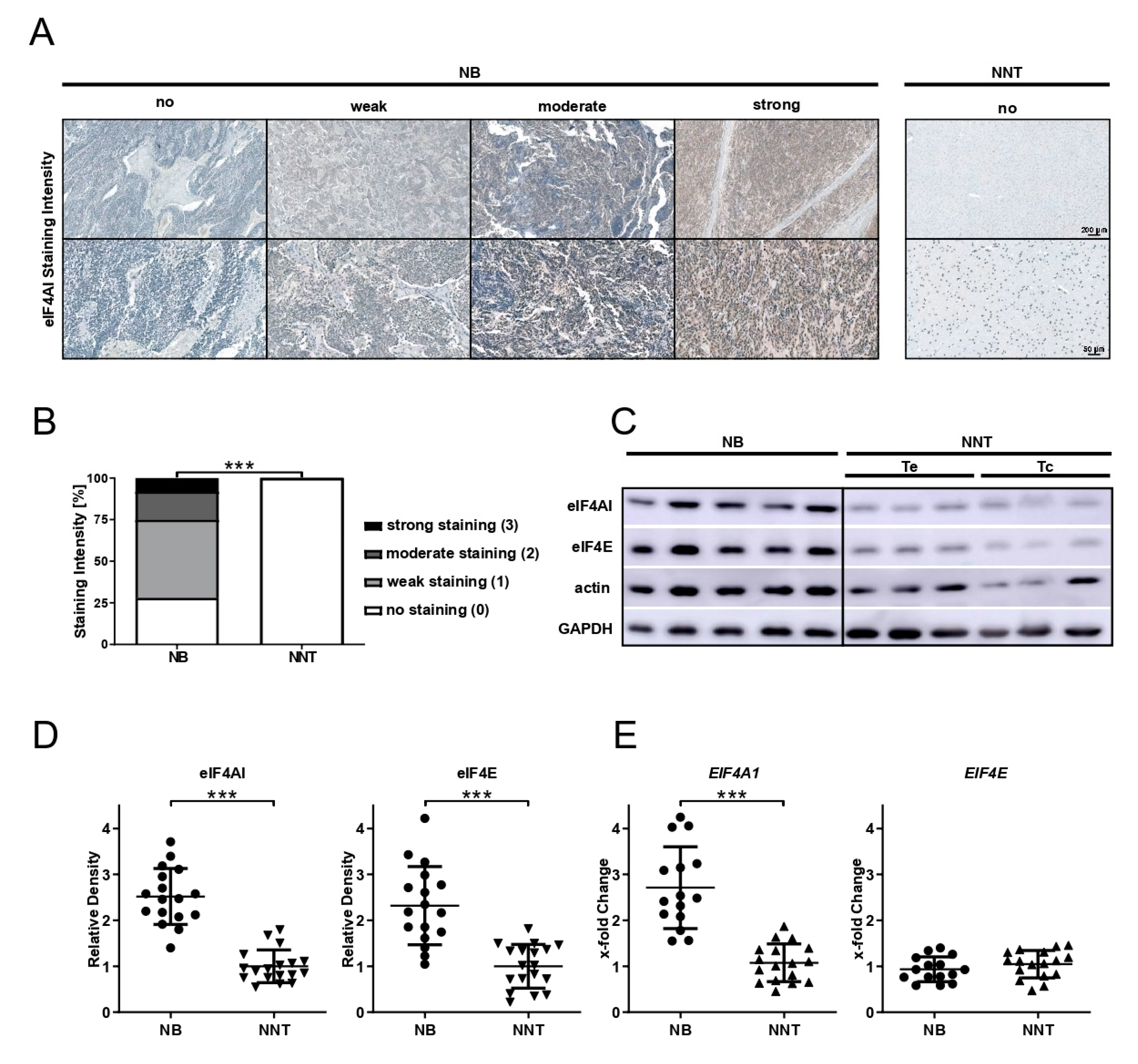
3.1. eIF4AI Is Significantly Overexpressed at the Protein and mRNA Level in FFPE and Fresh Frozen Human Neuroblastic Tissue Compared to Non-Neoplastic Tissue
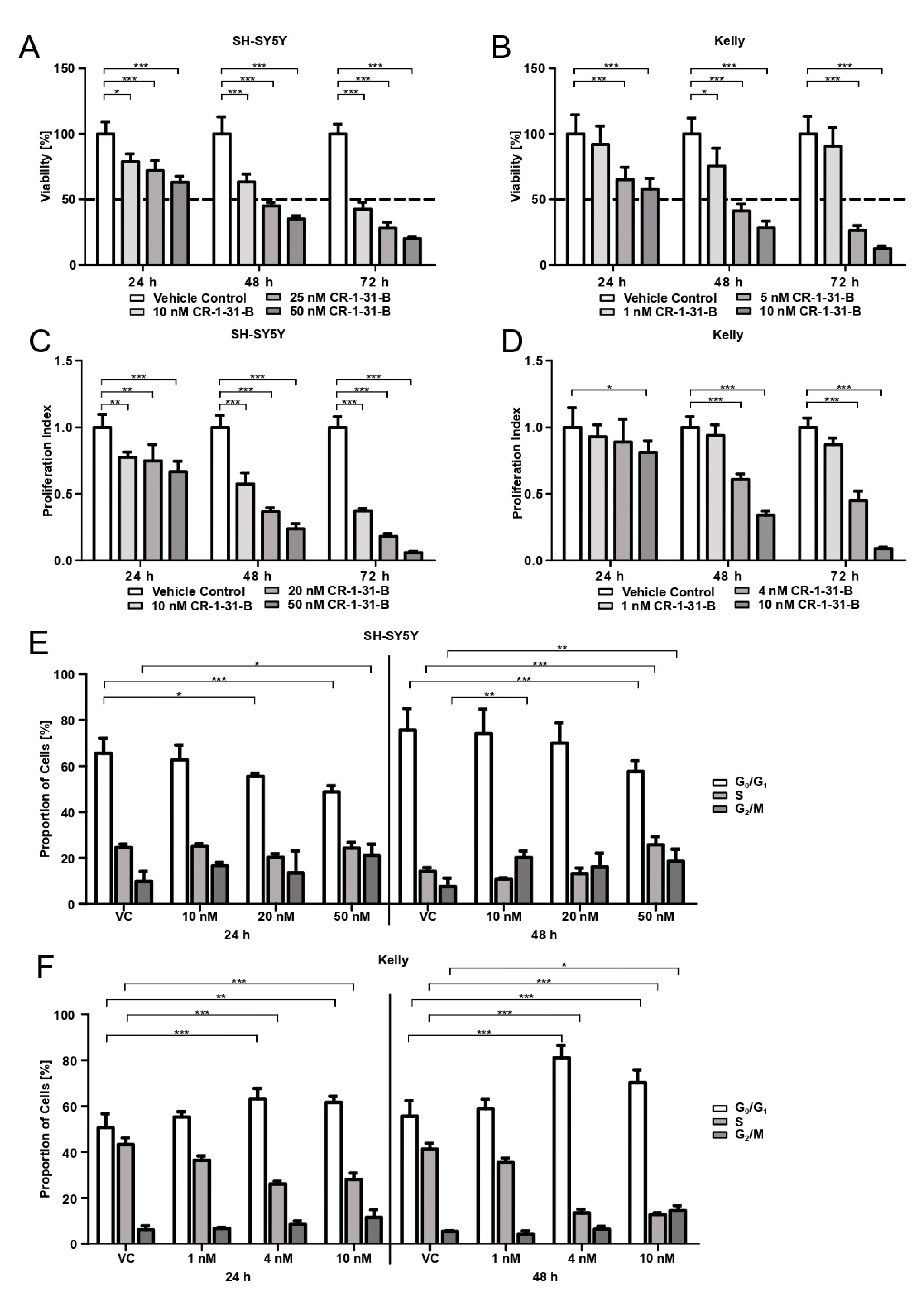
3.2. Targeting eIF4AI with CR-1-31-B Reduces Viability, Proliferation and Changes Cell Cycle Phase Distribution at Low nM Doses
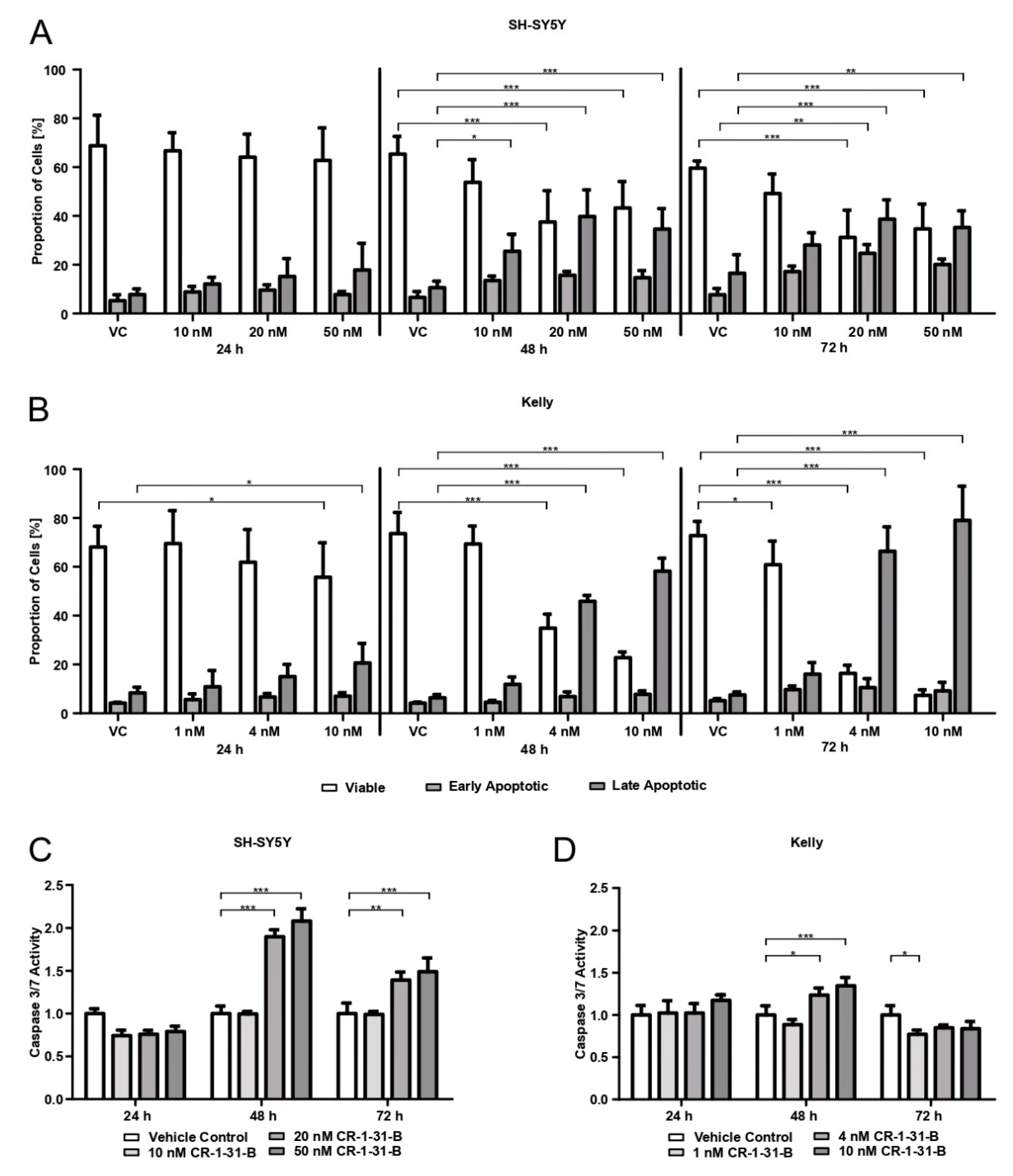
3.3. CR-1-31-B Treatment Leads to Apoptotic Cell Death of SH-SY5Y and Kelly Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Park, J.R.; Eggert, A.; Caron, H. Neuroblastoma: Biology, Prognosis, and Treatment. Hematol. Oncol. Clin. N. Am. 2010, 24, 65–86. [Google Scholar] [CrossRef] [PubMed]
- Whittle, S.B.; Smith, V.; Doherty, E.; Zhao, S.; McCarty, S.; Zage, P.E. Overview and recent advances in the treatment of neuroblastoma. Expert Rev. Anticancer Ther. 2017, 17, 369–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvera, D.; Formenti, S.C.; Schneider, R.J. Translational control in cancer. Nat. Rev. Cancer 2010, 10, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Robichaud, N.; Sonenberg, N.; Ruggero, D.; Schneider, R.J. Translational Control in Cancer. Cold Spring Harb. Perspect. Biol. 2019, 11, a032896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izycka-Świeszewska, E.; Drozyńska, E.; Rzepko, R.; Kobierska-Gulida, G.; Grajkowska, W.; Perek, D.; Balcerska, A. Analysis of PI3K/AKT/mTOR Signalling Pathway in High Risk Neuroblastic Tumours. Pol. J. Pathol. 2010, 61, 192–198. [Google Scholar]
- Brown, R.E.; Tan, D.; Taylor, J.S.; Miller, M.; Prichard, J.W.; Kott, M.M. Brief communication: Morphoproteomic Confirmation of Constitutively Activated mTOR, ERK, and NF-kappaB Pathways in High Risk Neuroblastoma, With Cell Cycle and Protein Analyte Correlates. Ann. Clin. Lab. Sci. 2007, 37, 141–147. [Google Scholar]
- Pelletier, J.; Graff, J.; Ruggero, D.; Sonenberg, N. Targeting the eIF4F Translation Initiation Complex: A Critical Nexus for Cancer Development. Cancer Res. 2015, 75, 250–263. [Google Scholar] [CrossRef] [Green Version]
- Galicia-Vazquez, G.; Cencic, R.; Robert, F.; Agenor, A.Q.; Pelletier, J. A Cellular Response Linking eIF4AI Activity to eIF4AII Transcription. RNA 2012, 18, 1373–1384. [Google Scholar] [CrossRef] [Green Version]
- Lazaris-Karatzas, A.; Montine, K.S.; Sonenberg, N. Malignant Transformation by a Eukaryotic Initiation Factor Subunit that Binds to mRNA 5′ Cap. Nature 1990, 345, 544–547. [Google Scholar] [CrossRef]
- Ruggero, D.; Montanaro, L.; Ma, L.; Xu, W.; Londei, P.; Cordon-Cardo, C.; Pandolfi, P.P. The Translation Factor eIF-4E Promotes Tumor Formation and Cooperates with c-Myc in Lymphomagenesis. Nat. Med. 2004, 10, 484–486. [Google Scholar] [CrossRef]
- Li, B.D.L.; McDonald, J.C.; Nassar, R.; De Benedetti, A. Clinical Outcome in Stage I to III Breast Carcinoma and eIF4E Overexpression. Ann. Surg. 1998, 227, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-L. Detection of Eukaryotic Translation Initiation Factor 4E and Its Clinical Significance in Hepatocellular Carcinoma. World J. Gastroenterol. 2012, 18, 2540. [Google Scholar] [CrossRef] [PubMed]
- Comtesse, N.; Keller, A.; Diesinger, I.; Bauer, C.; Kayser, K.; Huwer, H.; Lenhof, H.P.; Meese, E. Frequent Overexpression of the Genes FXR1, CLAPM1 and EIF4G Located on Amplicon 3q26-27 in Squamous Cell Carcinoma of the Lung. Int. J. Cancer 2007, 120, 2538–2544. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Zhou, Y.; Chen, Y.; Ke, G.; Wen, H.; Wu, X. Decreased Expression of EIF4A1 After Preoperative Brachytherapy Predicts Better Tumor-Specific Survival in Cervical Cancer. Int. J. Gynecol. Cancer 2014, 24, 908–915. [Google Scholar] [CrossRef]
- Dmitriev, S.E.; Vladimirov, D.O.; Lashkevich, K.A. A Quick Guide to Small-Molecule Inhibitors of Eukaryotic Protein Synthesis. Biochemistry 2020, 85, 1389–1421. [Google Scholar] [CrossRef]
- Kantarjian, H.M.; O’Brien, S.; Cortes, J. Homoharringtonine/omacetaxine mepesuccinate: The long and winding road to food and drug administration approval. Clin. Chem. Myeloma Leuk. 2013, 13, 530–533. [Google Scholar] [CrossRef] [Green Version]
- Garreau De Loubresse, N.; Prokhorova, I.; Holtkamp, W.; Rodnina, M.V.; Yusupova, G.; Yusupov, M. Structural basis for the inhibition of the eukaryotic ribosome. Nature 2014, 513, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.; Pelletier, J. Therapeutic Opportunities in Eukaryotic Translation. Cold Spring Harb. Perspect. Biol. 2018, 10, a032995. [Google Scholar] [CrossRef]
- Cencic, R.; Carrier, M.; Galicia-Vázquez, G.; Bordeleau, M.-E.; Sukarieh, R.; Bourdeau, A.; Brem, B.; Teodoro, J.G.; Greger, H.; Tremblay, M.L.; et al. Antitumor Activity and Mechanism of Action of the Cyclopenta[b]benzofuran, Silvestrol. PLoS ONE 2009, 4, e5223. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.; Galicia-Vázquez, G.; Cencic, R.; Mills, J.R.; Katigbak, A.; Porco, J.A.; Pelletier, J. CRISPR-Mediated Drug-Target Validation Reveals Selective Pharmacological Inhibition of the RNA Helicase, eIF4A. Cell Rep. 2016, 15, 2340–2347. [Google Scholar] [CrossRef] [Green Version]
- Hwang, B.Y.; Su, B.N.; Chai, H.; Mi, Q.; Kardono, L.B.S.; Afriastini, J.J.; Riswan, S.; Santarsiero, B.D.; Mesecar, A.D.; Wild, R.; et al. Silvestrol and Episilvestrol, Potential Anticancer Rocaglate Derivatives from Aglaia silvestris. J. Org. Chem. 2004, 69, 3350–3358. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, C.M.; Cencic, R.; Roche, S.P.; Pelletier, J.; Porco, J.A. Synthesis of Rocaglamide Hydroxamates and Related Compounds as Eukaryotic Translation Inhibitors: Synthetic and Biological Studies. J. Med. Chem. 2012, 55, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.; Anderson, C.; Weiss, K.L.; Grimley, M.; Sorrells, D. Eukaryotic Initiation Factor 4E Staining as a Clinical Marker in Pediatric Neuroblastoma. J. Pediatr. Hematol. Oncol. 2004, 26, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Golob-Schwarzl, N.; Wodlej, C.; Kleinegger, F.; Gogg-Kamerer, M.; Birkl-Toeglhofer, A.M.; Petzold, J.; Aigelsreiter, A.; Thalhammer, A.; Park, Y.N.; Haybaeck, J. Eukaryotic Translation Initiation Factor 6 Overexpression Plays a Major Role in the Translational Control of Gallbladder Cancer. J. Cancer Res. Clin. Oncol. 2019, 145, 2699–2711. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. 2016, 44, e71. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Therneau, T.M.; Grambsch, P.M. Therneau, T.M.; Grambsch, P.M. The Cox Model. In Modeling Survival Data: Extending the Cox Model; Statistics for Biology and Health; Springer: New York, NY, USA, 2000. [Google Scholar]
- Bhat, M.; Robichaud, N.; Hulea, L.; Sonenberg, N.; Pelletier, J.; Topisirovic, I. Targeting the Translation Machinery in Cancer. Nat. Rev. Drug Discov. 2015, 14, 261–278. [Google Scholar] [CrossRef]
- Wolfe, A.L.; Singh, K.; Zhong, Y.; Drewe, P.; Rajasekhar, V.K.; Sanghvi, V.R.; Mavrakis, K.J.; Jiang, M.; Roderick, J.E.; Van der Meulen, J.; et al. RNA G-quadruplexes cause eIF4A-dependent oncogene translation in cancer. Nature 2014. [Google Scholar] [CrossRef] [Green Version]
- Sadlish, H.; Galicia-Vazquez, G.; Paris, C.G.; Aust, T.; Bhullar, B.; Chang, L.; Helliwell, S.B.; Hoepfner, D.; Knapp, B.; Riedl, R.; et al. Evidence for a Functionally Relevant Rocaglamide Binding Site on the eIF4A–RNA Complex. ACS Chem. Biol. 2013, 8, 1519–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.; Robert, F.; Oertlin, C.; Kapeller-Libermann, D.; Avizonis, D.; Gutierrez, J.; Handly-Santana, A.; Doubrovin, M.; Park, J.; Schoepfer, C.; et al. eIF4A Supports an Oncogenic Translation Program in Pancreatic Ductal Adenocarcinoma. Nat. Commun. 2019, 10, 5151. [Google Scholar] [CrossRef] [PubMed]
- Rust, A.; Shah, S.; Hautbergue, G.M.; Davletov, B. Burkholderia lethal factor 1, a novel anti-cancer toxin, demonstrates selective cytotoxicity in MYCN-amplified neuroblastoma cells. Toxins 2018, 10, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwab, M.; Alitalo, K.; Klempnauer, K.-H.; Varmus, H.E.; Bishop, J.M.; Gilbert, F.; Brodeur, G.; Goldstein, M.; Trent, J. Amplified DNA with Limited Homology to myc Cellular Oncogene is Shared by Human Neuroblastoma Cell Lines and a Neuroblastoma Tumour. Nature 1983, 305, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Kong, T.; Xue, Y.; Cencic, R.; Zhu, X.; Monast, A.; Fu, Z.; Pilon, V.; Sangwan, V.; Guiot, M.C.; Foulkes, W.D.; et al. eIF4A Inhibitors Suppress Cell-Cycle Feedback Response and Acquired Resistance to CDK4/6 Inhibition in Cancer. Mol. Cancer Ther. 2019, 18, 2158–2170. [Google Scholar] [CrossRef] [Green Version]
- Brodeur, G.M.; Seeger, R.C.; Schwab, M.; Varmus, H.E.; Bishop, J.M. Amplification of N-myc in Untreated Human Neuroblastomas Correlates with Advanced Disease Stage. Science 1984, 224, 1121–1124. [Google Scholar] [CrossRef]
- Dhar, M.L.; Dhar, M.M.; Dhawan, B.N.; Mehrotra, B.N.; Srimal, R.C.; Tandon, J.S. Screening of Indian plants for biological activity IV. Indian J. Exp. Biol. 1973, 11, 43–54. [Google Scholar]
- Robert, F.; Roman, W.; Bramoullé, A.; Fellmann, C.; Roulston, A.; Shustik, C.; Porco, J.A., Jr.; Shore, G.C.; Sebag, M.; Pelletier, J. Translation initiation factor eIF4F modifies the dexamethasone response in multiple myeloma. Proc. Natl. Acad. Sci. USA 2014, 111, 13421–13426. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.; Zhang, W.; Cencic, R.; Devine, W.G.; Beglov, D.; Henkel, T.; Brown, L.E.; Vajda, S.; Porco, J.A., Jr.; Pelletier, J. Amidino-Rocaglates: A Potent Class of eIF4A Inhibitors. Cell Chem. Biol. 2019, 26, 1586–1593.e3. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Tissue for Biochemical Analyses (n = 17) | NB Tissue for IHC Analyses (n = 36) | NNT for IHC and Biochemical Analyses (n = 18) | |
|---|---|---|---|
| Age (Median) | 15 m | 27 m | 61 y |
| Gender | |||
| Female | 7 (41%) | 15 (42%) | 4 (22%) |
| Male | 10 (59%) | 21 (58%) | 14 (78%) |
| Stage | |||
| 1 | 1 (6%) | 5 (14%) | |
| 2 | 3 (18%) | 4 (11%) | |
| 3 | 4 (24%) | 13 (36%) | |
| 4 | 3 (18%) | 12 (33%) | |
| 5 | 0 (0%) | 2 (6%) | |
| Unknown | 6 (35%) | 0 (0%) | |
| MYCN Status | |||
| Non-amplified | 13 (76%) | 26 (72%) | |
| Amplified | 2 (12%) | 10 (28%) | |
| Gain | 1 (6%) | 0 (0%) | |
| Unknown | 1 (6%) | 0 (0%) | |
| Localization | |||
| Adrenal gland | 4 (24%) | 20 (56%) | |
| Retroperitoneal | 1 (6%) | 8 (22%) | |
| Mediastinal | 0 (0%) | 8 (22%) | |
| Abdominal | 3 (18%) | 0 (0%) | |
| Thoracic | 4 (24%) | 0 (0%) | 9 (50%) |
| Others | 5 (29%) | 0 (0%) | |
| Temporal lobe | 0 (0%) | 0 (0%) | 9 (50%) |
| Diagnosis | |||
| Ganglioneuroma | 0 (0%) | 4 (11%) | |
| Ganglioneuroblastoma | 6 (35%) | 5 (14%) | |
| Neuroblastoma | 11 (65%) | 27 (75%) | |
| Differentiated | n.d. | 13 (48%) | |
| Poorly differentiated | n.d. | 12 (44%) | |
| Undifferentiated | n.d. | 2 (7%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skofler, C.; Kleinegger, F.; Krassnig, S.; Birkl-Toeglhofer, A.M.; Singer, G.; Till, H.; Benesch, M.; Cencic, R.; Porco, J.A., Jr.; Pelletier, J.; et al. Eukaryotic Translation Initiation Factor 4AI: A Potential Novel Target in Neuroblastoma. Cells 2021, 10, 301. https://doi.org/10.3390/cells10020301
Skofler C, Kleinegger F, Krassnig S, Birkl-Toeglhofer AM, Singer G, Till H, Benesch M, Cencic R, Porco JA Jr., Pelletier J, et al. Eukaryotic Translation Initiation Factor 4AI: A Potential Novel Target in Neuroblastoma. Cells. 2021; 10(2):301. https://doi.org/10.3390/cells10020301
Chicago/Turabian StyleSkofler, Christina, Florian Kleinegger, Stefanie Krassnig, Anna Maria Birkl-Toeglhofer, Georg Singer, Holger Till, Martin Benesch, Regina Cencic, John A. Porco, Jr., Jerry Pelletier, and et al. 2021. "Eukaryotic Translation Initiation Factor 4AI: A Potential Novel Target in Neuroblastoma" Cells 10, no. 2: 301. https://doi.org/10.3390/cells10020301
APA StyleSkofler, C., Kleinegger, F., Krassnig, S., Birkl-Toeglhofer, A. M., Singer, G., Till, H., Benesch, M., Cencic, R., Porco, J. A., Jr., Pelletier, J., Castellani, C., Raicht, A., Izycka-Swieszewska, E., Czapiewski, P., & Haybaeck, J. (2021). Eukaryotic Translation Initiation Factor 4AI: A Potential Novel Target in Neuroblastoma. Cells, 10(2), 301. https://doi.org/10.3390/cells10020301