
Direct Conversion of Human Fibroblasts into Adipocytes Using a Novel Small Molecular Compound: Implications for Regenerative Therapy for Adipose Tissue Defects

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Libraries and Compounds
2.2. Culture Media
2.3. Cells
2.4. Retroviral Vectors
2.5. Primary Screening
2.6. Secondary Screening
2.7. Tertiary and Quaternary Screening
2.8. Chemical Direct Conversion
2.9. Real Time RT-PCR
2.10. Immunostaining
2.11. ELISA
2.12. Glucose Uptake Assay
2.13. Surgical Procedure and Cell Implantation
2.14. Immunohistochemistry and Histology
2.15. Data Analysis
3. Results
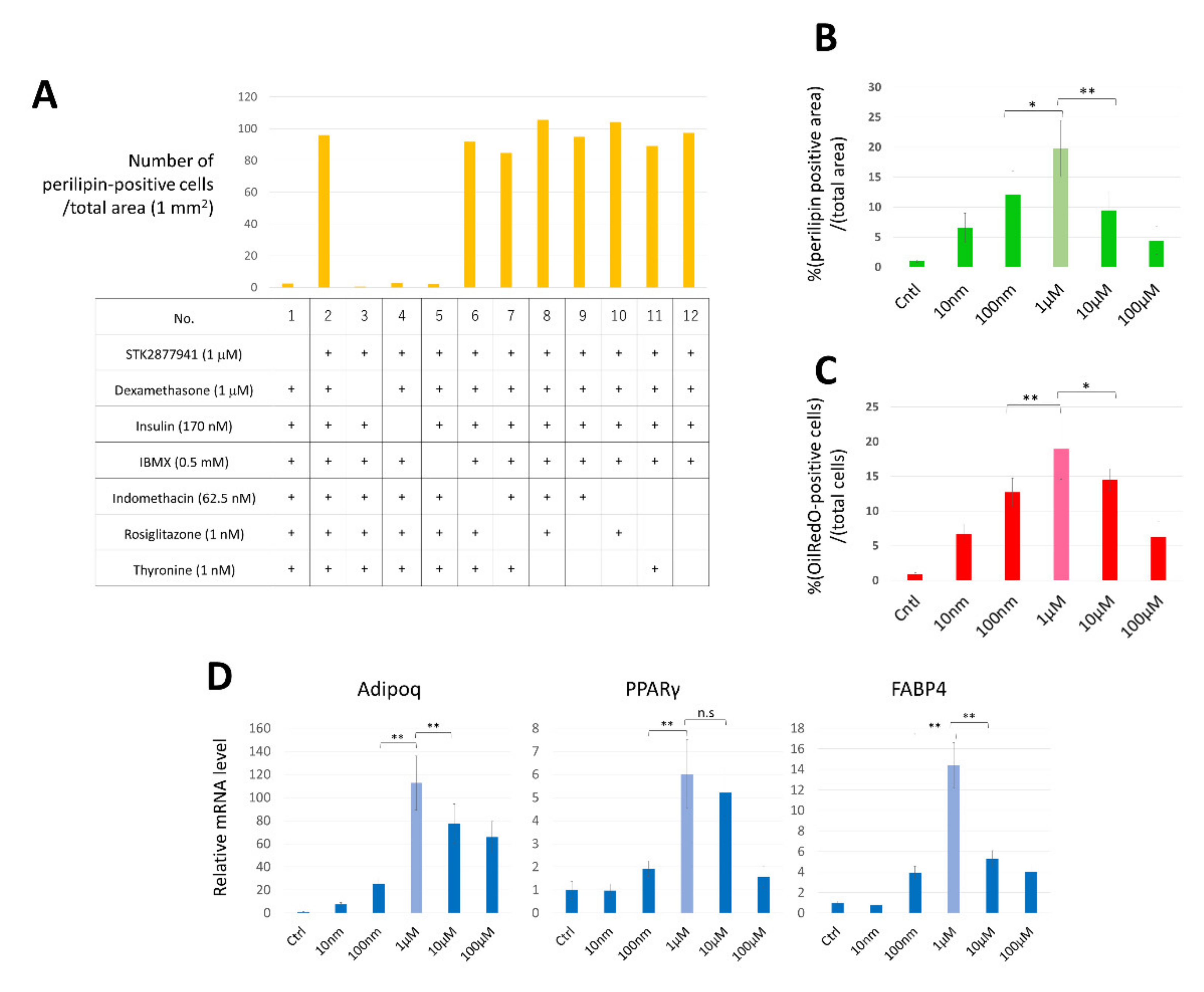
3.1. Identification of a Molecule That Induces an Adipocyte Phenotype in HDFs
3.2. Determination of Suitable Culture Conditions for CCCA Induction
3.3. Upregulation of C/EBPα and PPARγ during Conversion of HDFs into CCCAs
3.4. Absence of a Pluripotent Stem Cell-Like State during Conversion of HDFs into CCCAs
3.5. CCCAs Share Similar Functional Properties with Mature Adipocytes In Vitro
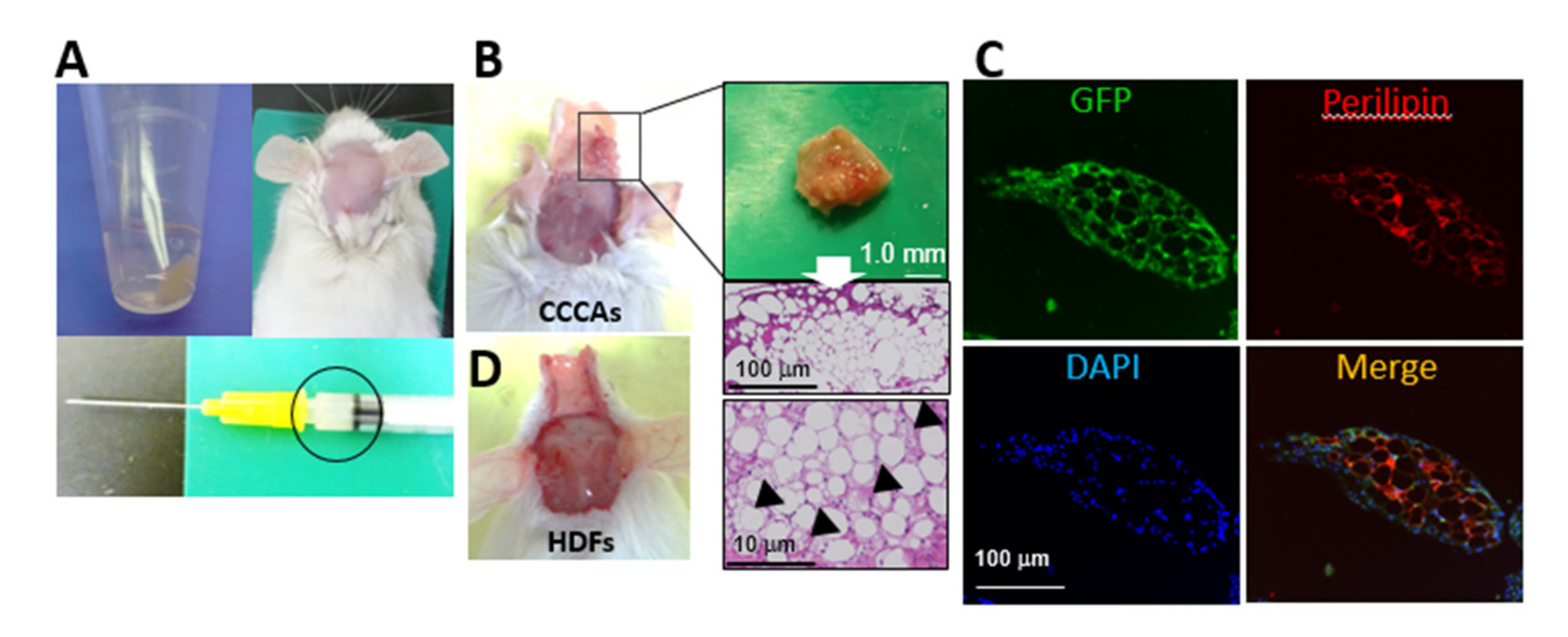
3.6. CCCAs Remain Adipocyte-Like and Are Found in Granulation Tissue Formed after Implantation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vallejo, A.; Garcia-Ruano, A.A.; Pinilla, C.; Castellano, M.; Deleyto, E.; Perez-Cano, R. Comparing Efficacy and Costs of Four Facial Fillers in Human Immunodeficiency Virus-Associated Lipodystrophy: A Clinical Trial. Plast. Reconstr. Surg. 2018, 141, 613–623. [Google Scholar] [CrossRef]
- Shuck, J.; Iorio, M.L.; Hung, R.; Davison, S.P. Autologous fat grafting and injectable dermal fillers for human immunodeficiency virus-associated facial lipodystrophy: A comparison of safety, efficacy, and long-term treatment outcomes. Plast. Reconstr. Surg. 2013, 131, 499–506. [Google Scholar] [CrossRef]
- Nelson, L.; Stewart, K.J. Plastic surgical options for HIV-associated lipodystrophy. J. Plast. Reconstr. Aesthetic Surg. 2008, 61, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Salibian, A.A.; Frey, J.D.; Bekisz, J.M.; Choi, M.; Karp, N.S. Fat Grafting and Breast Augmentation: A Systematic Review of Primary Composite Augmentation. Plast. Reconstr. Surg. Glob. Open 2019, 7, e2340. [Google Scholar] [CrossRef] [PubMed]
- Mallon, P.W. Pathogenesis of lipodystrophy and lipid abnormalities in patients taking antiretroviral therapy. AIDS Rev. 2007, 9, 3–15. [Google Scholar]
- Suzuki, K.; Akita, S.; Yoshimoto, H.; Ohtsuru, A.; Hirano, A.; Yamashita, S. Biological Features Implies Potential Use of Autologous Adipose-Derived Stem/Progenitor Cells in Wound Repair and Regenerations for the Patients with Lipodystrophy. Int. J. Mol. Sci. 2019, 20, 5505. [Google Scholar] [CrossRef] [Green Version]
- Stolarczyk, E. Adipose tissue inflammation in obesity: A metabolic or immune response? Curr. Opin. Pharmacol. 2017, 37, 35–40. [Google Scholar] [CrossRef]
- Zhang, H.H.; Kumar, S.; Barnett, A.H.; Eggo, M.C. Ceiling culture of mature human adipocytes: Use in studies of adipocyte functions. J. Endocrinol. 2000, 164, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessard, J.; Pelletier, M.; Biertho, L.; Biron, S.; Marceau, S.; Hould, F.S.; Lebel, S.; Moustarah, F.; Lescelleur, O.; Marceau, P.; et al. Characterization of dedifferentiating human mature adipocytes from the visceral and subcutaneous fat compartments: Fibroblast-activation protein alpha and dipeptidyl peptidase 4 as major components of matrix remodeling. PLoS ONE 2015, 10, e0122065. [Google Scholar] [CrossRef]
- Inoue, Y.; Kishida, T.; Kotani, S.I.; Akiyoshi, M.; Taga, H.; Seki, M.; Ukimura, O.; Mazda, O. Direct conversion of fibroblasts into urothelial cells that may be recruited to regenerating mucosa of injured urinary bladder. Sci. Rep. 2019, 9, 13850. [Google Scholar] [CrossRef] [Green Version]
- Sowa, Y.; Kishida, T.; Tomita, K.; Yamamoto, K.; Numajiri, T.; Mazda, O. Direct Conversion of Human Fibroblasts into Schwann Cells that Facilitate Regeneration of Injured Peripheral Nerve In Vivo. Stem Cells Transl. Med. 2017, 6, 1207–1216. [Google Scholar] [CrossRef]
- Wakao, J.; Kishida, T.; Fumino, S.; Kimura, K.; Yamamoto, K.; Kotani, S.I.; Mizushima, K.; Naito, Y.; Yoshikawa, T.; Tajiri, T.; et al. Efficient direct conversion of human fibroblasts into myogenic lineage induced by co-transduction with MYCL and MYOD1. Biochem. Biophys. Res. Commun. 2017, 488, 368–373. [Google Scholar] [CrossRef]
- Yamamoto, K.; Kishida, T.; Sato, Y.; Nishioka, K.; Ejima, A.; Fujiwara, H.; Kubo, T.; Yamamoto, T.; Kanamura, N.; Mazda, O. Direct conversion of human fibroblasts into functional osteoblasts by defined factors. Proc. Natl. Acad. Sci. USA 2015, 112, 6152–6157. [Google Scholar] [CrossRef] [Green Version]
- Kishida, T.; Ejima, A.; Yamamoto, K.; Tanaka, S.; Yamamoto, T.; Mazda, O. Reprogrammed Functional Brown Adipocytes Ameliorate Insulin Resistance and Dyslipidemia in Diet-Induced Obesity and Type 2 Diabetes. Stem Cell Rep. 2015, 5, 569–581. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Hu, W.; Qiu, B.; Zhao, J.; Yu, Y.; Guan, W.; Wang, M.; Yang, W.; Pei, G. Generation of neural progenitor cells by chemical cocktails and hypoxia. Cell Res. 2014, 24, 665–679. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zuo, X.; Jing, J.; Ma, Y.; Wang, J.; Liu, D.; Zhu, J.; Du, X.; Xiong, L.; Du, Y.; et al. Small-Molecule-Driven Direct Reprogramming of Mouse Fibroblasts into Functional Neurons. Cell Stem Cell 2015, 17, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Chabrat, A.; Lacassagne, E.; Billiras, R.; Landron, S.; Pontisso-Mahout, A.; Darville, H.; Dupront, A.; Coge, F.; Schenker, E.; Piwnica, D.; et al. Pharmacological Transdifferentiation of Human Nasal Olfactory Stem Cells into Dopaminergic Neurons. Stem Cells Int. 2019, 2019, 2945435. [Google Scholar] [CrossRef]
- Wan, X.Y.; Xu, L.Y.; Li, B.; Sun, Q.H.; Ji, Q.L.; Huang, D.D.; Zhao, L.; Xiao, Y.T. Chemical conversion of human lung fibroblasts into neuronal cells. Int. J. Mol. Med. 2018, 41, 1463–1468. [Google Scholar] [CrossRef] [Green Version]
- Thoma, E.C.; Merkl, C.; Heckel, T.; Haab, R.; Knoflach, F.; Nowaczyk, C.; Flint, N.; Jagasia, R.; Jensen Zoffmann, S.; Truong, H.H.; et al. Chemical conversion of human fibroblasts into functional Schwann cells. Stem Cell Rep. 2014, 3, 539–547. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Kishida, T.; Nakai, K.; Sato, Y.; Kotani, S.I.; Nishizawa, Y.; Yamamoto, T.; Kanamura, N.; Mazda, O. Direct phenotypic conversion of human fibroblasts into functional osteoblasts triggered by a blockade of the transforming growth factor-β signal. Sci. Rep. 2018, 8, 8463. [Google Scholar] [CrossRef]
- Hacein-Bey-Abina, S.; von Kalle, C.; Schmidt, M.; Le Deist, F.; Wulffraat, N.; McIntyre, E.; Radford, I.; Villeval, J.L.; Fraser, C.C.; Cavazzana-Calvo, M.; et al. A serious adverse event after successful gene therapy for X-linked severe combined immunodeficiency. N. Engl. J. Med. 2003, 348, 255–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banyard, D.A.; Salibian, A.A.; Widgerow, A.D.; Evans, G.R. Implications for human adipose-derived stem cells in plastic surgery. J. Cell Mol. Med. 2015, 19, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Luan, J.; Xin, M.; Wang, Q.; Xiao, R.; Gao, Y. Fate of adiposederived stromal vascular fraction cells after co-implantation with fat grafts: Evidence of cell survival and differentiation in ischemic adipose tissue. Plast. Reconstr. Surg. 2013, 132, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.Y.; Yim, S.; Kim, H.J.; Jin, U.S.; Lim, S.; Eo, S.; Chang, H.; Minn, K.W. The Fate of the Adipose-Derived Stromal Cells during Angiogenesis and Adipogenesis after Cell-Assisted Lipotransfer. Plast. Reconstr. Surg. 2018, 141, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Choy, L.; Skillington, J.; Derynck, R. Roles of autocrine TGF-beta receptor and Smad signaling in adipocyte differentiation. J. Cell Biol. 2000, 149, 667–682. [Google Scholar] [CrossRef] [PubMed]
- Fajas, L.; Fruchart, J.C.; Auwerx, J. Transcriptional control of adipogenesis. Curr. Opin. Cell Biol. 1998, 10, 165–173. [Google Scholar] [CrossRef]
- Rosen, E.D. The transcriptional basis of adipocyte development. Prostaglandins Leukot. Essent. Fat. Acids 2005, 73, 31–34. [Google Scholar] [CrossRef]
- Farmer, S.R. Transcriptional control of adipocyte formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Bae, S.; Yoon, Y. The anti-adipogenic effects of (-)epigallocatechin gallate are dependent on the WNT/β-catenin pathway. J. Nutr. Biochem. 2013, 24, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Li, Y.; Zhao, T.; Wang, Y.; Sun, C. Ursolic acid inhibits adipogenesis in 3T3-L1 adipocytes through LKB1/AMPK pathway. PLoS ONE 2013, 8, e70135. [Google Scholar] [CrossRef] [Green Version]
- Oishi, Y.; Manabe, I.; Tobe, K.; Tsushima, K.; Shindo, T.; Fujiu, K.; Nishimura, G.; Maemura, K.; Yamauchi, T.; Kubota, N.; et al. Krüppel-like transcription factor KLF5 is a key regulator of adipocyte differentiation. Cell Metab. 2005, 1, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPalpha induces adipogenesis through PPARgamma: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Porse, B.T.; Pedersen, T.A.; Xu, X.; Lindberg, B.; Wewer, U.M.; Friis-Hansen, L.; Nerlov, C. E2F repression by C/EBPalpha is required for adipogenesis and granulopoiesis in vivo. Cell 2001, 107, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; McKeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBP alpha and PPAR gamma controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell 1999, 3, 151–158. [Google Scholar] [CrossRef]
- Freytag, S.O.; Geddes, T.J. Reciprocal regulation of adipogenesis by Myc and C/EBP alpha. Science 1992, 256, 379–382. [Google Scholar] [CrossRef]
- Zhu, S.; Cheng, G.; Zhu, H.; Guan, G. A study of genes involved in adipocyte differentiation. J. Pediatr. Endocrinol. Metab. 2015, 28, 93–99. [Google Scholar] [CrossRef]
- Zhu, J.; Pang, D.; Zhou, Y.; Tang, X.; Huang, Y.; Xie, W.; Gao, F.; Lai, L.; Zhang, M.; Ouyang, H. Direct conversion of porcine embryonic fibroblasts into adipocytes by chemical molecules. Cell. Reprogram. 2012, 14, 99–105. [Google Scholar] [CrossRef]
- Cristofalo, V.J.; Allen, R.G.; Pignolo, R.J.; Martin, B.G.; Beck, J.C. Relationship between donor age and the replicative lifespan of human cells in culture: A reevaluation. Proc. Natl. Acad. Sci. USA 1998, 95, 10614–10619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Li, L. Current reprogramming systems in regenerative medicine: From somatic cells to induced pluripotent stem cells. Regen. Med. 2016, 11, 105–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagami, H. The mechanism of white and brown adipocyte differentiation. Diabetes Metab. J. 2013, 37, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Stephens, J.M. The fat controller: Adipocyte development. PLoS Biol. 2012, 10, e1001436. [Google Scholar] [CrossRef] [Green Version]
- Zeniou, M.; Nguekeu-Zebaze, L.; Dantzer, F. Therapeutic considerations of PARP in stem cell biology: Relevance in cancer and beyond. Biochem. Pharmacol. 2019, 167, 107–115. [Google Scholar] [CrossRef]
- Ishay-Ronen, D.; Diepenbruck, M.; Kalathur, R.K.R.; Sugiyama, N.; Tiede, S.; Ivanek, R.; Bantug, G.; Morini, M.F.; Wang, J.; Hess, C.; et al. Gain Fat-Lose Metastasis: Converting Invasive Breast Cancer Cells into Adipocytes Inhibits Cancer Metastasis. Cancer Cell 2019, 35, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Basu-Roy, U.; Han, E.; Rattanakorn, K.; Gadi, A.; Verma, N.; Maurizi, G.; Gunaratne, P.H.; Coarfa, C.; Kennedy, O.D.; Garabedian, M.J.; et al. PPARgamma agonists promote differentiation of cancer stem cells by restraining YAP transcriptional activity. Oncotarget 2016, 7, 60954–60970. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Yu, D.B.; Kim, M.; Choi, Y.L. Adipogenesis induces growth inhibition of dedifferentiated liposarcoma. Cancer Sci. 2019, 110, 2676–2683. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.B.; Onder, T.T.; Jiang, G.; Tao, K.; Kuperwasser, C.; Weinberg, R.A.; Lander, E.S. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell 2009, 138, 645–659. [Google Scholar] [CrossRef] [Green Version]
- Miehle, K.; Ebert, T.; Kralisch, S.; Hoffmann, A.; Kratzsch, J.; Schlögl, H.; Stumvoll, M.; Fasshauer, M. Adipocyte and epidermal fatty acid-binding protein serum concentrations in patients with lipodystrophy. Cytokine 2017, 92, 20–23. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sowa, Y.; Kishida, T.; Louis, F.; Sawai, S.; Seki, M.; Numajiri, T.; Takahashi, K.; Mazda, O. Direct Conversion of Human Fibroblasts into Adipocytes Using a Novel Small Molecular Compound: Implications for Regenerative Therapy for Adipose Tissue Defects. Cells 2021, 10, 605. https://doi.org/10.3390/cells10030605
Sowa Y, Kishida T, Louis F, Sawai S, Seki M, Numajiri T, Takahashi K, Mazda O. Direct Conversion of Human Fibroblasts into Adipocytes Using a Novel Small Molecular Compound: Implications for Regenerative Therapy for Adipose Tissue Defects. Cells. 2021; 10(3):605. https://doi.org/10.3390/cells10030605
Chicago/Turabian StyleSowa, Yoshihiro, Tsunao Kishida, Fiona Louis, Seiji Sawai, Makoto Seki, Toshiaki Numajiri, Kenji Takahashi, and Osam Mazda. 2021. "Direct Conversion of Human Fibroblasts into Adipocytes Using a Novel Small Molecular Compound: Implications for Regenerative Therapy for Adipose Tissue Defects" Cells 10, no. 3: 605. https://doi.org/10.3390/cells10030605
APA StyleSowa, Y., Kishida, T., Louis, F., Sawai, S., Seki, M., Numajiri, T., Takahashi, K., & Mazda, O. (2021). Direct Conversion of Human Fibroblasts into Adipocytes Using a Novel Small Molecular Compound: Implications for Regenerative Therapy for Adipose Tissue Defects. Cells, 10(3), 605. https://doi.org/10.3390/cells10030605