
SRSF9 Regulates Cassette Exon Splicing of Caspase-2 by Interacting with Its Downstream Exon

 , , ,
, , ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, RNA Extraction and RT-PCR
2.2. Plasmid Construction
2.3. shRNA Treatment and Plasmid Transfection
2.4. RNA Pulldown Assay and Immunoblotting Assay
2.5. Splicing Analysis of RNA-seq and Gene Ontology (GO) Analysis
2.6. Correlation Analysis
2.7. Annexin V/PI Staining
2.8. Quantitation and Statistical Analysis
3. Results
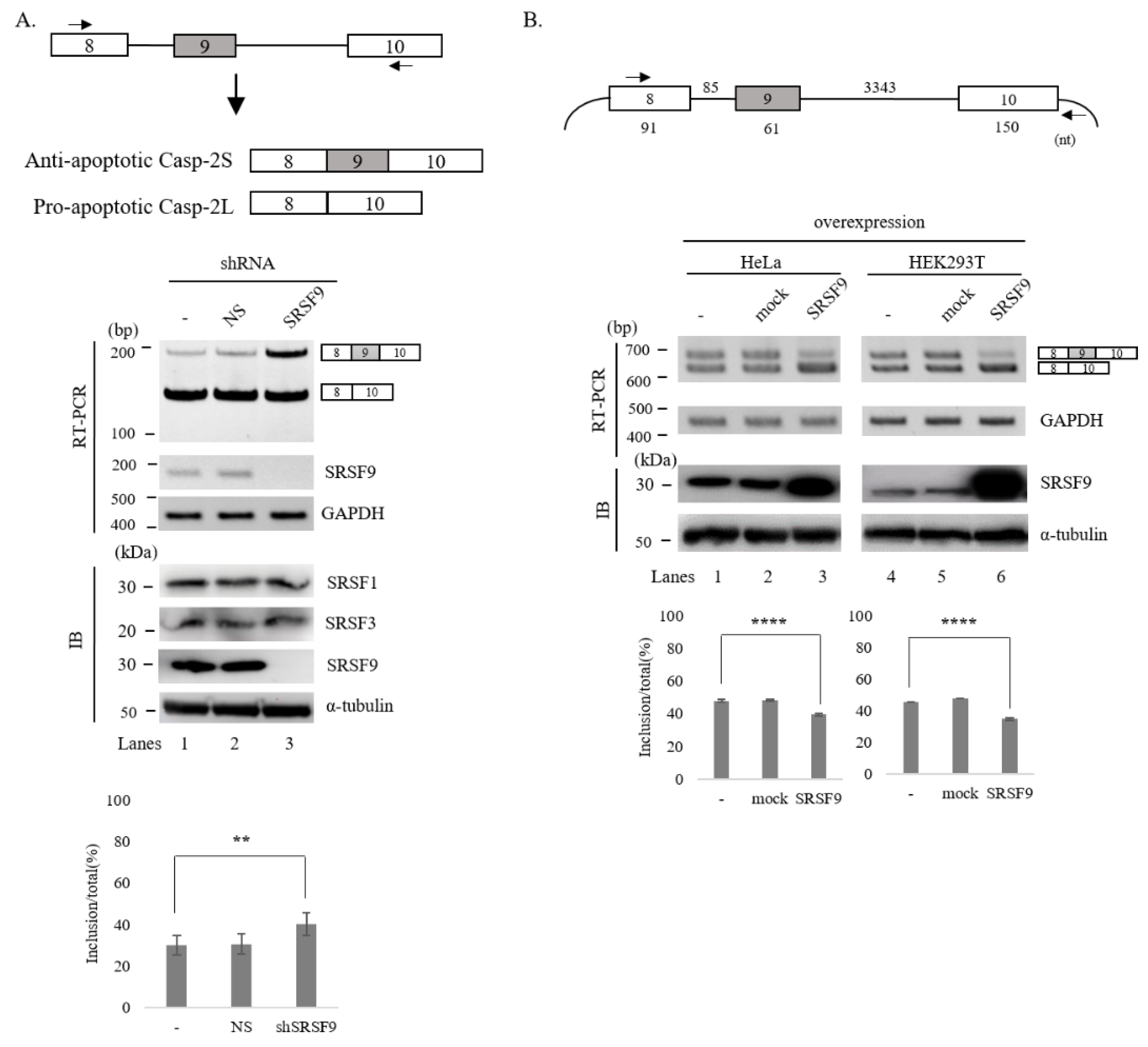
3.1. SRSF9 Regulates AS of Caspase-2
3.2. SRSF9 Targets Downstream Exon to Regulate AS of Caspase-2
3.3. SRSF9 Interacts with the Functional Target Sequence
3.4. SRSF9 Globally Affects AS of Apoptosis-Related Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Padgett, R.A.; Gr1, A.; Padgett, R.; Grabowski, P.J.; Konarska, M.M.; Seiler, S.; Sharp, P.A. Splicing of Messenger RNA Precursors. Annu. Rev. Biochem. 1986, 55, 1119–1150. [Google Scholar] [CrossRef] [PubMed]
- Maniatis, T.; Reed, R. The role of small nuclear ribonucleoprotein particles in pre-mRNA splicing. Nature 1987, 325, 673–678. [Google Scholar] [CrossRef]
- Gebauer, F.; Schwarzl, T.; Valcárcel, J.; Hentze, M.W. RNA-binding proteins in human genetic disease. Nat. Rev. Genet. 2021, 22, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Will, C.L.; Luhrmann, R. Spliceosome structure and function. Cold Spring Harb. Perspect. Biol. 2011, 3, a003707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcrip-tome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef]
- Nilsen, T.W.; Graveley, B.R. Expansion of the eukaryotic proteome by alternative splicing. Nature 2010, 463, 457–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghigna, C.; Valacca, C.; Biamonti, G. Alternative Splicing and Tumor Progression. Curr. Genom. 2008, 9, 556–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, M.; Schweer, D.; Ziegler, A.; Gaber, R.; Schock, S.; Schwinzer, R.; Wonigeit, K.; Lindert, R.-B.; Kantarci, O.; Schaefer-Klein, J.; et al. A point mutation in PTPRC is associated with the development of multiple sclerosis. Nat. Genet. 2000, 26, 495–499. [Google Scholar] [CrossRef]
- Cartegni, L.; Krainer, A.R. Disruption of an SF2/ASF-dependent exonic splicing enhancer in SMN2 causes spinal muscular atrophy in the absence of SMN1. Nat. Genet. 2002, 30, 377–384. [Google Scholar] [CrossRef]
- Cooper, T.A.; Wan, L.; Dreyfuss, G. RNA and Disease. Cell 2009, 136, 777–793. [Google Scholar] [CrossRef] [Green Version]
- Wong, R.S.Y. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadeel, B.; Orrenius, S.; Zhivotovsky, B. Apoptosis in human disease: A new skin for the old ceremony? Biochem. Biophys. Res. Commun. 1999, 266, 699–717. [Google Scholar] [CrossRef] [Green Version]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.S.; Yuan, J. Caspases in apoptosis and beyond. Oncogene 2008, 27, 6194–6206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safa, A.R.; Day, T.W.; Wu, C.H. Cellular FLICE-like inhibitory protein (C-FLIP): A novel target for cancer therapy. Curr. Cancer Drug Targets 2008, 8, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledgerwood, E.; Morison, I.M. Targeting the Apoptosome for Cancer Therapy. Clin. Cancer Res. 2009, 15, 420–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Mercatante, D.R.; Bortner, C.D.; Cidlowski, J.A.; Kole, R. Modification of alternative splicing of Bcl-x pre-mRNA in prostate and breast cancer cells. analysis of apoptosis and cell death. J. Biol. Chem. 2001, 276, 16411–16417. [Google Scholar] [CrossRef] [Green Version]
- Thomas, L.W.; Lam, C.; Edwards, S.W. Mcl-1 the molecular regulation of protein function. FEBS Lett. 2010, 584, 2981–2989. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.; Leo, C.P.; Hsu, S.Y.; Hsueh, A.J. MCL-1S, a Splicing Variant of the Antiapoptotic BCL-2 Family Member MCL-1, Encodes a Proapoptotic Protein Possessing Only the BH3 Domain. J. Biol. Chem. 2000, 275, 25255–25261. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, L.; Strasser, A.; O’Reilly, L.A.; Hausmann, G.; Adams, J.M.; Cory, S.; Huang, D.C. Bim: A novel member of the Bcl-2 family that promotes apoptosis. EMBO J. 1998, 17, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Hengartner, M.O. The biochemistry of apoptosis. Nat. Cell Biol. 2000, 407, 770–776. [Google Scholar] [CrossRef]
- Chautan, M.; Chazal, G.; Cecconi, F.; Gruss, P.; Golstein, P. Interdigital cell death can occur through a necrotic and caspa-se-independent pathway. Curr. Biol. 1999, 9, 967–970. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S. Caspase 2 in apoptosis, the DNA damage response and tumour suppression: Enigma no more? Nat. Rev. Cancer 2009, 9, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Krumschnabel, G.; Manzl, C.; Villunger, A. Caspase-2: Killer, savior and safeguard—Emerging versatile roles for an ill-defined caspase. Oncogene 2009, 28, 3093–3096. [Google Scholar] [CrossRef] [PubMed]
- Cote, J.; Dupuis, S.; Jiang, Z.; Wu, J.Y. Caspase-2 pre-mRNA alternative splicing: Identification of an intronic element containing a decoy 3’ acceptor site. Proc. Natl. Acad. Sci. USA 2001, 98, 938–943. [Google Scholar] [CrossRef]
- Wang, L.; Miura, M.; Bergeron, L.; Zhu, H.; Yuan, J. Ich-1, an Ice/ced-3-related gene, encodes both positive and negative regulators of programmed cell death. Cell 1994, 78, 739–750. [Google Scholar] [CrossRef]
- Côté, J.; Dupuis, S.; Wu, J.Y. Polypyrimidine Track-binding Protein Binding Downstream of Caspase-2 Alternative Exon 9 Represses Its Inclusion. J. Biol. Chem. 2001, 276, 8535–8543. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.N.; Lee, M.; Loh, T.J.; Choi, S.W.; Oh, H.K.; Moon, H.; Cho, S.; Hong, S.E.; Kim, D.H.; Sheng, Z.; et al. Exon 9 skipping of apoptotic caspase-2 pre-mRNA is promoted by SRSF3 through interaction with exon 8. Biochim. Biophys. Acta 2014, 1839, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.H.; Zhang, W.J.; Rao, Y.; Wu, J.Y. Regulation of Ich-1 pre-mRNA alternative splicing and apoptosis by mammalian splicing factors. Proc. Natl. Acad. Sci. USA 1998, 95, 9155–9160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kędzierska, H.; Piekiełko-Witkowska, A. Splicing factors of SR and hnRNP families as regulators of apoptosis in cancer. Cancer Lett. 2017, 396, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, K.; Ray, P.; Kar, A.; Wang, L.; Sutherland, L.C.; Wu, J.Y. Up-regulation of the proapoptotic caspase 2 splicing isoform by a candidate tumor suppressor, RBM5. Proc. Natl. Acad. Sci. USA 2008, 105, 15708–15713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graveley, B.R. Sorting out the complexity of SR protein functions. RNA 2000, 6, 1197–1211. [Google Scholar] [CrossRef] [Green Version]
- Paradis, C.; Cloutier, P.; Shkreta, L.; Toutant, J.; Klarskov, K.; Chabot, B. hnRNP I/PTB can antagonize the splicing repressor activity of SRp30c. RNA 2007, 13, 1287–1300. [Google Scholar] [CrossRef] [Green Version]
- Young, P.J.; DiDonato, C.J.; Hu, D.; Kothary, R.; Androphy, E.J.; Lorson, C.L. SRp30c-dependent stimulation of survival motor neuron (SMN) exon 7 inclusion is facilitated by a direct interaction with hTra2beta1. Hum. Mol. Genet. 2002, 11, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Simard, M.J.; Chabot, B. SRp30c Is a Repressor of 3′ Splice Site Utilization. Mol. Cell. Biol. 2002, 22, 4001–4010. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Liu, Y.; Choi, N.; Ha, J.; Pradella, D.; Ghigna, C.; Zheng, X.; Shen, H. Opposite Roles of Tra2beta and SRSF9 in the v10 Exon Splicing of CD44. Cancers 2020, 12, 3195. [Google Scholar] [CrossRef]
- Cloutier, P.; Toutant, J.; Shkreta, L.; Goekjian, S.; Revil, T.; Chabot, B. Antagonistic Effects of the SRp30c Protein and Cryptic 5 ′ Splice Sites on the Alternative Splicing of the Apoptotic Regulator Bcl-x. J. Biol. Chem. 2008, 283, 21315–21324. [Google Scholar] [CrossRef] [Green Version]
- Screaton, G.R.; Caceres, J.F.; Mayeda, A.; Bell, M.V.; Plebanski, M.; Jackson, D.G.; Bell, J.I.; Krainer, A.R. Identification and characterization of three members of the human SR family of pre-mRNA splicing factors. EMBO J. 1995, 14, 4336–4349. [Google Scholar] [CrossRef]
- Huang, H.; Kapeli, K.; Jin, W.; Wong, Y.P.; Arumugam, T.V.; Koh, J.H.; Srimasorn, S.; Mallilankaraman, K.; Chua, J.J.E.; Yeo, G.W.; et al. Tis-sue-selective restriction of RNA editing of CaV1.3 by splicing factor SRSF9. Nucleic Acids Res. 2018, 46, 7323–7338. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, R.; Zhang, F.; Srinivasan, H.; Richard, J.L.C.; Liu, K.I.; Zhang, X.; Woo, C.W.A.; Chua, Z.H.M.; Buschdorf, J.P.; Meaney, M.J.; et al. SRSF9 selectively represses ADAR2-mediated editing of brain-specific sites in primates. Nucleic Acids Res. 2018, 46, 7379–7395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Lv, R.; Guo, W.; Li, X. microRNA-802 inhibits cell proliferation and induces apoptosis in human cervical cancer by targeting serine/arginine-rich splicing factor 9. J. Cell Biochem. 2019, 120, 10370–10379. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Na Jang, H.; Liu, Y.; Choi, N.; Oh, J.; Ha, J.; Zheng, X.; Shen, H. Activation of Cryptic 3′ Splice-Sites by SRSF2 Contributes to Cassette Exon Skipping. Cells 2019, 8, 696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.; Park, J.W.; Lu, Z.X.; Lin, L.; Henry, M.D.; Wu, Y.N.; Zhou, Q.; Xing, Y. rMATS: Robust and flexible detection of differential alternative splicing from replicate RNA-Seq data. Proc. Natl. Acad. Sci. USA 2014, 111, E5593–E5601. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Collins, J.R.; Alvord, W.G.; Roayaei, J.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. The DAVID Gene Functional Classification Tool: A novel biological module-centric algorithm to functionally analyze large gene lists. Genome Biol. 2007, 8, R183. [Google Scholar] [CrossRef] [Green Version]
- Tapial, J.; Ha, K.C.H.; Sterne-Weiler, T.; Gohr, A.; Braunschweig, U.; Hermoso-Pulido, A.; Quesnel-Vallières, M.; Permanyer, J.; Sodaei, R.; Marquez, Y.; et al. An atlas of alternative splicing profiles and functional associations reveals new regulatory programs and genes that simultaneously express multiple major isoforms. Genome Res. 2017, 27, 1759–1768. [Google Scholar] [CrossRef] [Green Version]
- Piva, F.; Giulietti, M.; Burini, A.B.; Principato, G. SpliceAid 2: A database of human splicing factors expression data and RNA target motifs. Hum. Mutat. 2011, 33, 81–85. [Google Scholar] [CrossRef]
- Paz, I.; Kosti, I.; Ares, J.M.; Cline, M.; Mandel-Gutfreund, Y. RBPmap: A web server for mapping binding sites of RNA-binding proteins. Nucleic Acids Res. 2014, 42, W361–W367. [Google Scholar] [CrossRef]
- Xiao, W.; Adhikari, S.; Dahal, U.; Chen, Y.-S.; Hao, Y.-J.; Sun, B.-F.; Sun, H.-Y.; Li, A.; Ping, X.-L.; Lai, W.-Y.; et al. Nuclear m 6 A Reader YTHDC1 Regulates mRNA Splicing. Mol. Cell 2016, 61, 507–519. [Google Scholar] [CrossRef] [Green Version]
- David, C.J.; Manley, J.L. Alternative pre-mRNA splicing regulation in cancer: Pathways and programs unhinged. Genes Dev. 2010, 24, 2343–2364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoss, O.; Schwaiger, F.-W.; Cooper, T.A.; Stamm, S. Alternative Splicing Determines the Intracellular Localization of the Novel Nuclear Protein Nop30 and Its Interaction with the Splicing Factor SRp30c. J. Biol. Chem. 1999, 274, 10951–10962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raffetseder, U.; Frye, B.; Rauen, T.; Jürchott, K.; Royer, H.-D.; Jansen, P.L.; Mertens, P.R. Splicing Factor SRp30c Interaction with Y-box Protein-1 Confers Nuclear YB-1 Shuttling and Alternative Splice Site Selection. J. Biol. Chem. 2003, 278, 18241–18248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiede, B.; Dimmler, C.; Siejak, F.; Rudel, T. Predominant Identification of RNA-binding Proteins in Fas-induced Apoptosis by Proteome Analysis. J. Biol. Chem. 2001, 276, 26044–26050. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha, J.; Jang, H.; Choi, N.; Oh, J.; Min, C.; Pradella, D.; Jung, D.-W.; Williams, D.R.; Park, D.; Ghigna, C.; et al. SRSF9 Regulates Cassette Exon Splicing of Caspase-2 by Interacting with Its Downstream Exon. Cells 2021, 10, 679. https://doi.org/10.3390/cells10030679
Ha J, Jang H, Choi N, Oh J, Min C, Pradella D, Jung D-W, Williams DR, Park D, Ghigna C, et al. SRSF9 Regulates Cassette Exon Splicing of Caspase-2 by Interacting with Its Downstream Exon. Cells. 2021; 10(3):679. https://doi.org/10.3390/cells10030679
Chicago/Turabian StyleHa, Jiyeon, Hana Jang, Namjeong Choi, Jagyeong Oh, Chanhyuk Min, Davide Pradella, Da-Woon Jung, Darren R. Williams, Daeho Park, Claudia Ghigna, and et al. 2021. "SRSF9 Regulates Cassette Exon Splicing of Caspase-2 by Interacting with Its Downstream Exon" Cells 10, no. 3: 679. https://doi.org/10.3390/cells10030679