
Deletion of the Actin-Associated Tropomyosin Tpm3 Leads to Reduced Cell Complexity in Cultured Hippocampal Neurons—New Insights into the Role of the C-Terminal Region of Tpm3.1

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture and Transfection of NIH3T3 Cells
2.2. Plasmids
2.3. Transgenic Mice Used in the Study
2.4. Primary Culture of Mouse Hippocampal Neurons
2.5. Transfection of Mouse Primary Hippocampal Neurons
3. Immunocytochemistry
4. Imaging
5. Morphological and Statistical Analysis
6. Western Blotting
7. Results
7.1. Knock-Out of Tpm3 Gene Shows Decreased Length and Complexity of Both Axons and Dendrites of Primary Mouse Hippocampal Neurons
7.2. The C-Terminus of the Tpm3 Isoform Tpm3.1 is Important for Incorporation of Tropomyosin into Actin Filaments
7.3. C-Terminal Truncation of hTpm3.1 Impacts the Effect of hTpm3.1 Expression on Neuronal Morphology
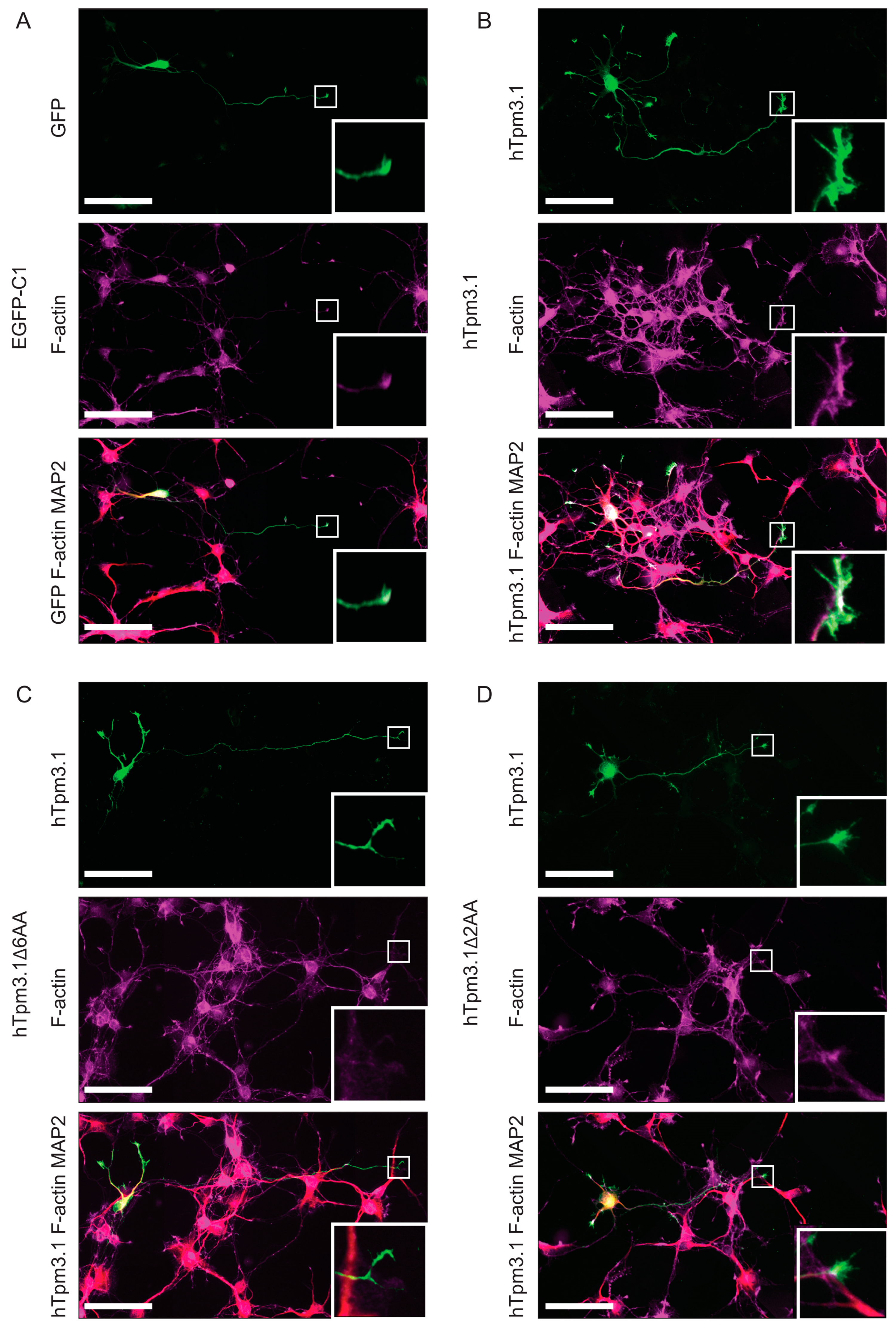
7.4. C-Terminal Truncation of Tpm3.1 Impacts the Segregation of hTpm3.1 to the Tips of Neurites
7.5. C-Terminal Deletion of hTpm3.1 Significantly Impacts the Amount of F-Actin Pool at the Growth Cone
8. Discussion
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bryce, N.S.; Schevzov, G.; Ferguson, V.; Percival, J.M.; Lin, J.J.-C.; Matsumura, F.; Bamburg, J.R.; Jeffrey, P.L.; Hardeman, E.C.; Gunning, P.; et al. Specification of Actin Filament Function and Molecular Composition by Tropomyosin Isoforms. Mol. Biol. Cell 2003, 14, 1002–1016. [Google Scholar] [CrossRef] [Green Version]
- Schevzov, G.; Bryce, N.S.; Almonte-Baldonado, R.; Joya, J.; Lin, J.J.-C.; Hardeman, E.; Weinberger, R.; Gunning, P. Specific Features of Neuronal Size and Shape Are Regulated by Tropomyosin Isoforms. Mol. Biol. Cell 2005, 16, 3425–3437. [Google Scholar] [CrossRef] [Green Version]
- Fath, T.; Chan, Y.-K.A.; Vrhovski, B.; Clarke, H.; Curthoys, N.; Hook, J.; Lemckert, F.; Schevzov, G.; Tam, P.; Watson, C.M.; et al. New aspects of tropomyosin-regulated neuritogenesis revealed by the deletion of Tm5NM1 and 2. Eur. J. Cell Biol. 2010, 89, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Guven, K.L.; Gunning, P.W.; Fath, T. TPM3 and TPM4 gene products segregate to the postsynaptic region of central nervous system synapses. Bioarchitecture 2011, 1, 284–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchowerska, A.K.; Fok, S.; Stefen, H.; Gunning, P.W.; Hardeman, E.C.; Power, J.; Fath, T. Developmental Profiling of Tropomyosin Expression in Mouse Brain Reveals Tpm4.2 as the Major Post-synaptic Tropomyosin in the Mature Brain. Front. Cell. Neurosci. 2017, 11, 421. [Google Scholar] [CrossRef] [PubMed]
- Abouelezz, A.; Stefen, H.; Segerstrale, M.; Micinski, D.; Minkeviciene, R.; Lahti, L.; Hardeman, E.C.; Gunning, P.W.; Hoogenraad, C.C.; Taira, T.; et al. Tropomyosin Tpm3.1 Is Required to Maintain the Structure and Function of the Axon Initial Segment. iScience 2020, 23, 101053. [Google Scholar] [CrossRef]
- Weinberger, R.; Schevzov, G.; Jeffrey, P.; Gordon, K.; Hill, M.; Gunning, P. The molecular composition of neuronal microfilaments is spatially and temporally regulated. J. Neurosci. 1996, 16, 238–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrhovski, B.; Schevzov, G.; Dingle, S.; Lessard, J.L.; Gunning, P.; Weinberger, R.P. Tropomyosin isoforms from the gamma gene differing at the C-terminus are spatially and developmentally regulated in the brain. J. Neurosci. Res. 2003, 72, 373–383. [Google Scholar] [CrossRef]
- Hook, J.; Lemckert, F.; Qin, H.; Schevzov, G.; Gunning, P. Gamma Tropomyosin Gene Products Are Required for Embryonic Development. Mol. Cell. Biol. 2004, 24, 2318–2323. [Google Scholar] [CrossRef] [Green Version]
- Curthoys, N.M.; Gunning, P.W.; Fath, T. Tropomyosins in neuronal morphogenesis and development. In Cytoskeleton of the Nervous System; Nixon, R.A., Yuan, A., Eds.; Springer: New York, NY, USA, 2011; pp. 411–445. [Google Scholar]
- Hitchcock-DeGregori, S.E.; Barua, B. Tropomyosin Structure, Function, and Interactions: A Dynamic Regulator. Sub. Biol. 2017, 82, 253–284. [Google Scholar] [CrossRef]
- Martin, C.; Gunning, P. Isoform sorting of tropomyosins. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2008; Volume 644, pp. 187–200. [Google Scholar]
- Schevzov, G.; Whittaker, S.P.; Fath, T.; Lin, J.J.-C.; Gunning, P.W. Tropomyosin isoforms and reagents. BioArchitecture 2011, 1, 135–164. [Google Scholar] [CrossRef] [Green Version]
- Geeves, M.A.; Hitchcock-DeGregori, S.E.; Gunning, P.W. A systematic nomenclature for mammalian tropomyosin isoforms. J. Muscle Res. Cell Motil. 2015, 36, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Jainchill, J.L.; Aaronson, S.A.; Todaro, G.J. Murine sarcoma and leukemia viruses: Assay using clonal lines of contact-inhibited mouse cells. J. Virol. 1969, 4, 549–553. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Gunning, P. The 3’-end of the human beta-actin gene enhances activity of the beta-actin expression vector system: Construction of improved vectors. J. Biochem. Biophys. Methods 1997, 36, 63–72. [Google Scholar] [CrossRef]
- Fath, T.; Ke, Y.D.; Gunning, P.; Götz, J.; Ittner, L.M. Primary support cultures of hippocampal and substantia nigra neurons. Nat. Protoc. 2008, 4, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Schevzov, G.; Vrhovski, B.; Bryce, N.S.; Elmir, S.; Qiu, M.R.; O’Neill, G.M.; Yang, N.; Verrills, N.M.; Kavallaris, M.; Gunning, P.W. Tissue-specific Tropomyosin Isoform Composition. J. Histochem. Cytochem. 2005, 53, 557–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schevzov, G.; Curthoys, N.M.; Gunning, P.W.; Fath, T. Functional Diversity of Actin Cytoskeleton in Neurons and its Regulation by Tropomyosin. Int. Rev. Cell Mol. Biol. Vol. 2012, 298, 33–94. [Google Scholar] [CrossRef]
- Gomez, T.M.; Letourneau, P.C. Actin dynamics in growth cone motility and navigation. J. Neurochem. 2014, 129, 221–234. [Google Scholar] [CrossRef]
- Blanchoin, L.; Pollard, T.D.; Hitchcock-DeGregori, S.E. Inhibition of the Arp2/3 complex-nucleated actin polymerization and branch formation by tropomyosin. Curr. Biol. 2001, 11, 1300–1304. [Google Scholar] [CrossRef] [Green Version]
- Ono, S.; Ono, K. Tropomyosin inhibits ADF/cofilin-dependent actin filament dynamics. J. Cell Biol. 2002, 156, 1065–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broschat, K.O.; Burgess, D.R. Low Mr tropomyosin isoforms from chicken brain and intestinal epithelium have distinct actin-binding properties. J. Biol. Chem. 1986, 261, 13350–13359. [Google Scholar] [CrossRef]
- Novy, R.E.; Lin, J.L.; Lin, C.S.; Lin, J.J. Human fibroblast tropomyosin isoforms: Characterization of cDNA clones and analysis of tropomyosin isoform expression in human tissues and in normal and transformed cells. Cell Motil. Cytoskelet. 1993, 25, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Moraczewska, J.; Nicholson-Flynn, K.; Hitchcock-DeGregori, S.E. The Ends of Tropomyosin Are Major Determinants of Actin Affinity and Myosin Subfragment 1-Induced Binding to F-Actin in the Open State. Biochemistry 1999, 38, 15885–15892. [Google Scholar] [CrossRef]
- Michelot, A.; Drubin, D.G. Building Distinct Actin Filament Networks in a Common Cytoplasm. Curr. Biol. 2011, 21, R560–R569. [Google Scholar] [CrossRef] [Green Version]
- Skau, C.T.; Neidt, E.M.; Kovar, D.R. Role of Tropomyosin in Formin-mediated Contractile Ring Assembly in Fission Yeast. Mol. Biol. Cell 2009, 20, 2160–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houle, F.; Poirier, A.; Dumaresq, J.; Huot, J. DAP kinase mediates the phosphorylation of tropomyosin-1 downstream of the ERK pathway, which regulates the formation of stress fibers in response to oxidative stress. J. Cell Sci. 2007, 120, 3666–3677. [Google Scholar] [CrossRef] [Green Version]
- Coulton, A.T.; East, D.A.; Galinska-Rakoczy, A.; Lehman, W.; Mulvihill, D.P. The recruitment of acetylated and unacetylated tropomyosin to distinct actin polymers permits the discrete regulation of specific myosins in fission yeast. J. Cell Sci. 2010, 123, 3235–3243. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, N.; Ishii, I.; Habara, Y.; Allen, C.B.; Chun, J. Dual Regulation of Actin Rearrangement through Lysophosphatidic Acid Receptor in Neuroblast Cell Lines: Actin Depolymerization by Ca2+-α-Actinin and Polymerization by Rho. Mol. Biol. Cell 2002, 13, 2692–2705. [Google Scholar] [CrossRef] [Green Version]
- Sobue, K.; Kanda, K. α-Actinins, Calspectin (Brain Spectrin or Fodrin), and Actin Participate in Adhesion and Movement of Growth Cones. Neuron 1989, 3, 311–319. [Google Scholar] [CrossRef]
- Hall, A. Rho GTPases and the Actin Cytoskeleton. Science 1998, 279, 509–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, D.S.; Gaspar, C.J.; Domingos, P.M. Ire1 Mediated mRNA Splicing in a C-Terminus Deletion Mutant of Drosophila Xbp1. PLoS ONE 2014, 9, e105588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiina, N.; Shinkura, K.; Tokunaga, M. A Novel RNA-Binding Protein in Neuronal RNA Granules: Regulatory Machinery for Local Translation. J. Neurosci. 2005, 25, 4420–4434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alami, N.H.; Smith, R.B.; Carrasco, M.A.; Williams, L.A.; Winborn, C.S.; Han, S.S.; Kiskinis, E.; Winborn, B.; Freibaum, B.D.; Kanagaraj, A.; et al. Axonal Transport of TDP-43 mRNA Granules Is Impaired by ALS-Causing Mutations. Neuron 2014, 81, 536–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, K.C.; Pak, C.W.; Shaw, A.E.; Bradke, F.; Bamburg, J.R. Growth cone-like waves transport actin and promote axonogenesis and neurite branching. Dev. Neurobiol. 2009, 69, 761–779. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomanić, T.; Martin, C.; Stefen, H.; Parić, E.; Gunning, P.; Fath, T. Deletion of the Actin-Associated Tropomyosin Tpm3 Leads to Reduced Cell Complexity in Cultured Hippocampal Neurons—New Insights into the Role of the C-Terminal Region of Tpm3.1. Cells 2021, 10, 715. https://doi.org/10.3390/cells10030715
Tomanić T, Martin C, Stefen H, Parić E, Gunning P, Fath T. Deletion of the Actin-Associated Tropomyosin Tpm3 Leads to Reduced Cell Complexity in Cultured Hippocampal Neurons—New Insights into the Role of the C-Terminal Region of Tpm3.1. Cells. 2021; 10(3):715. https://doi.org/10.3390/cells10030715
Chicago/Turabian StyleTomanić, Tamara, Claire Martin, Holly Stefen, Esmeralda Parić, Peter Gunning, and Thomas Fath. 2021. "Deletion of the Actin-Associated Tropomyosin Tpm3 Leads to Reduced Cell Complexity in Cultured Hippocampal Neurons—New Insights into the Role of the C-Terminal Region of Tpm3.1" Cells 10, no. 3: 715. https://doi.org/10.3390/cells10030715