
A Histological and Morphometric Assessment of the Adult and Juvenile Rat Livers after Mild Traumatic Brain Injury

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Conditions
2.2. Induction of a Mild Traumatic Brain Injury
2.3. Experiment Design
- Group I (n = 70; sexually mature males):
- -
- Group IA (n = 10; control group),
- -
- Group IB (n = 60; animals with model mild traumatic brain injury)
- Group II (n = 70; young males):
- -
- Group IIA (n = 10; control group),
- -
- Group IIB (n = 60; animals with a model mild traumatic brain injury)
2.4. Histomorphometric Analysis
2.5. Statistical Analyses
3. Results
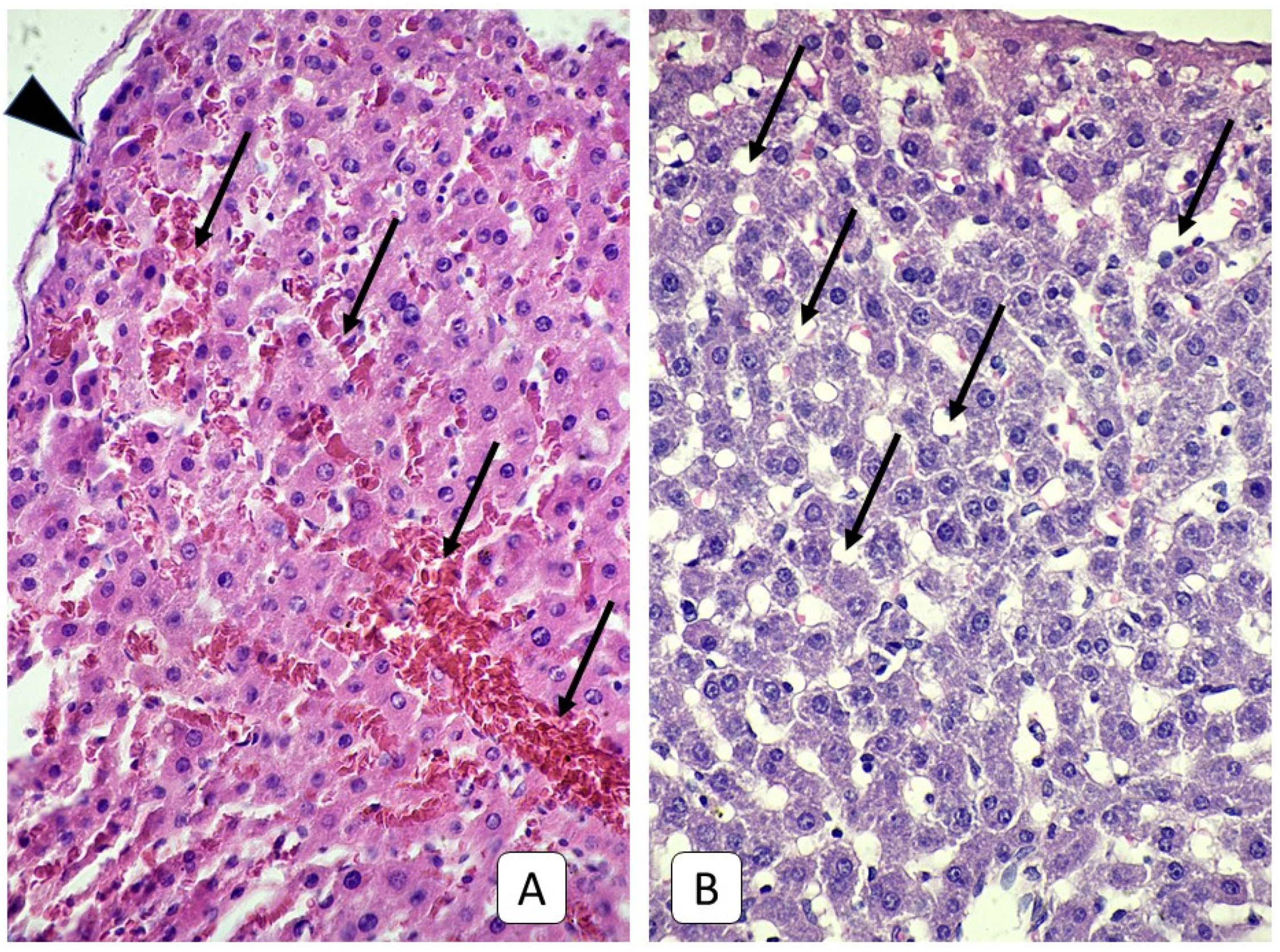
3.1. Histological Examinations of Rat Livers on the First Day after Injury
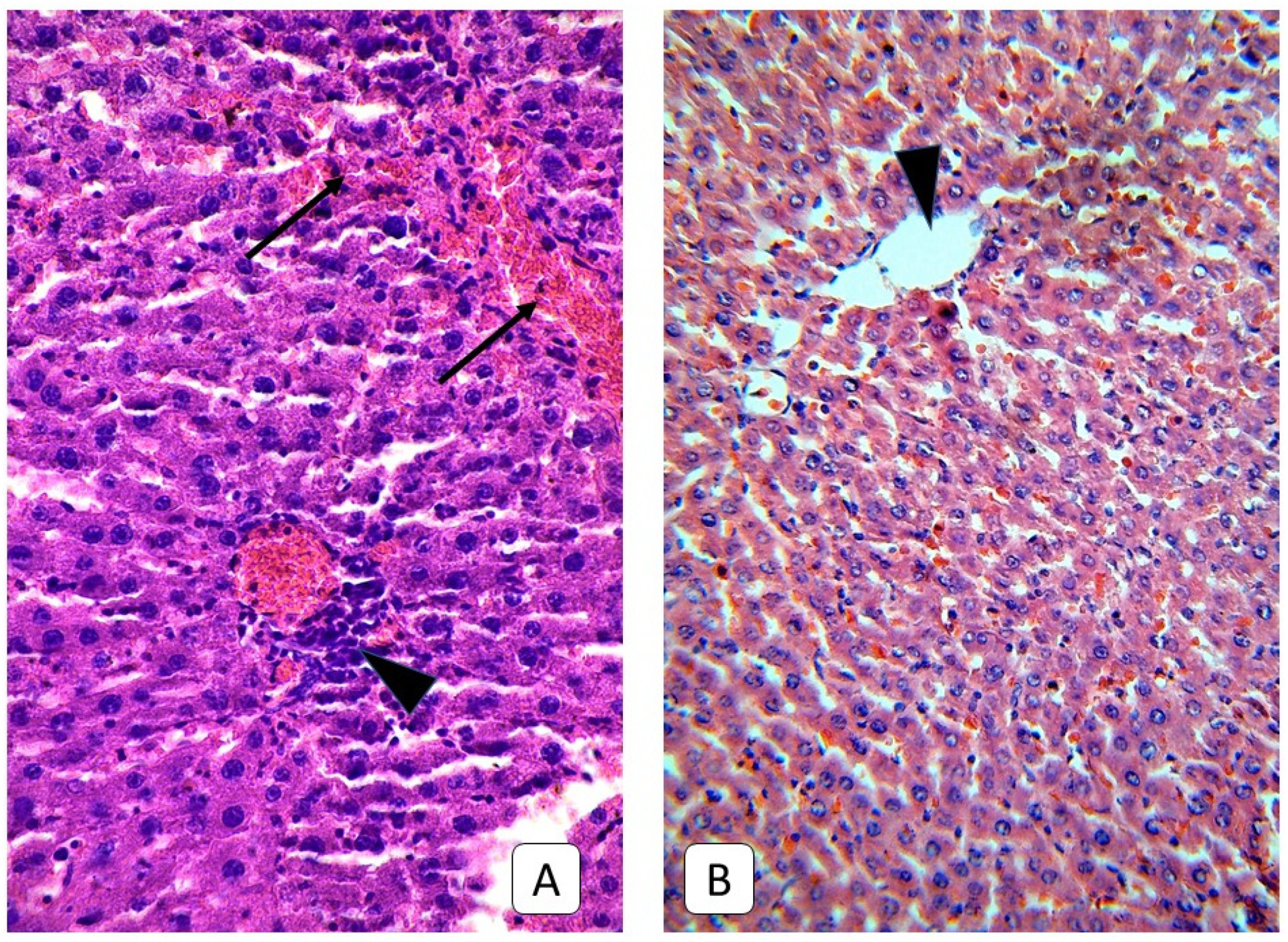
3.2. Histological Examinations of Rat Livers on the Third Day after Injury
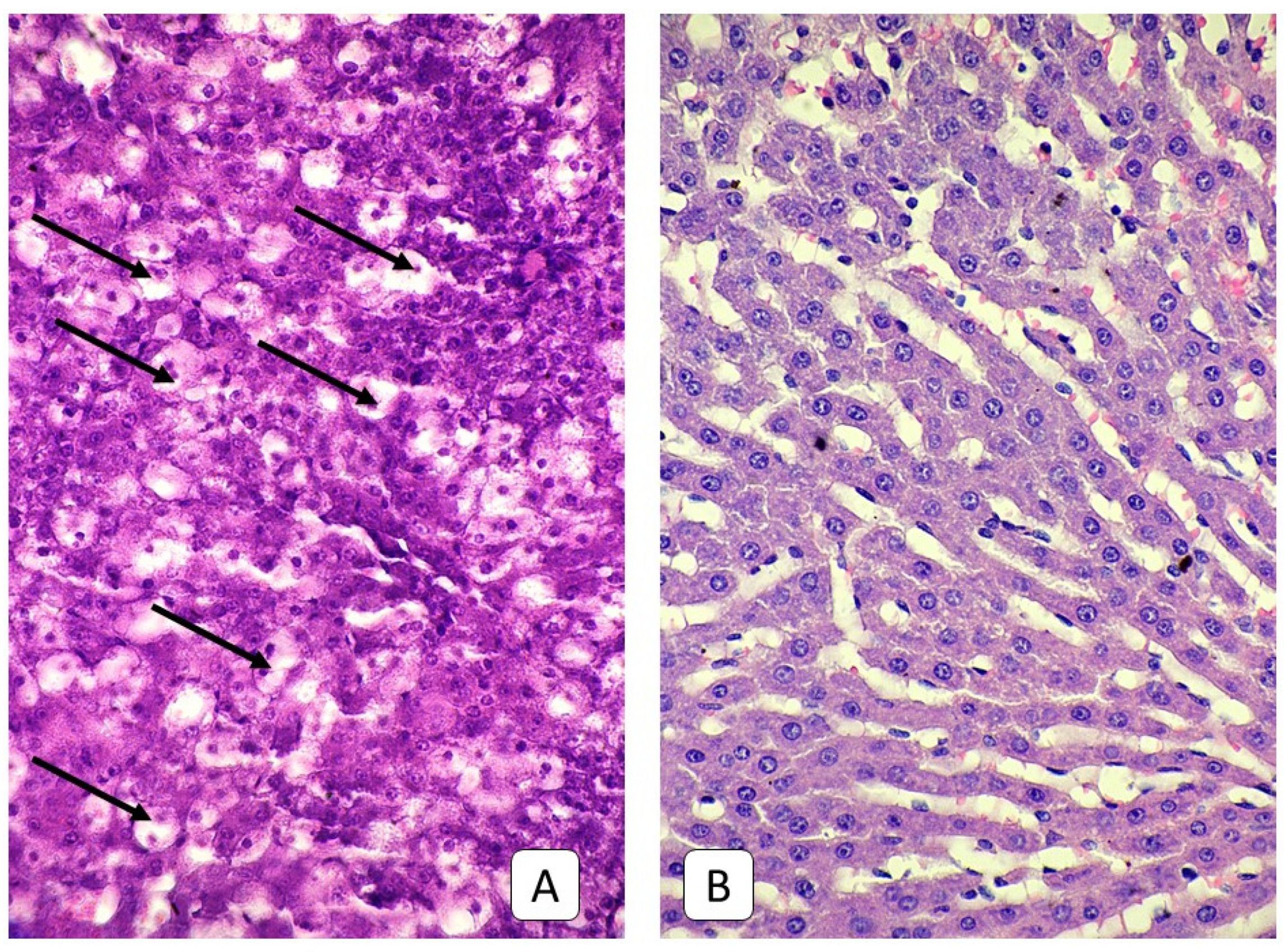
3.3. Histological Examinations of Rat Livers on the Fifth Day after Injury
3.4. Histological Examinations of Rats’ Liver on the Seventh Day after Trauma
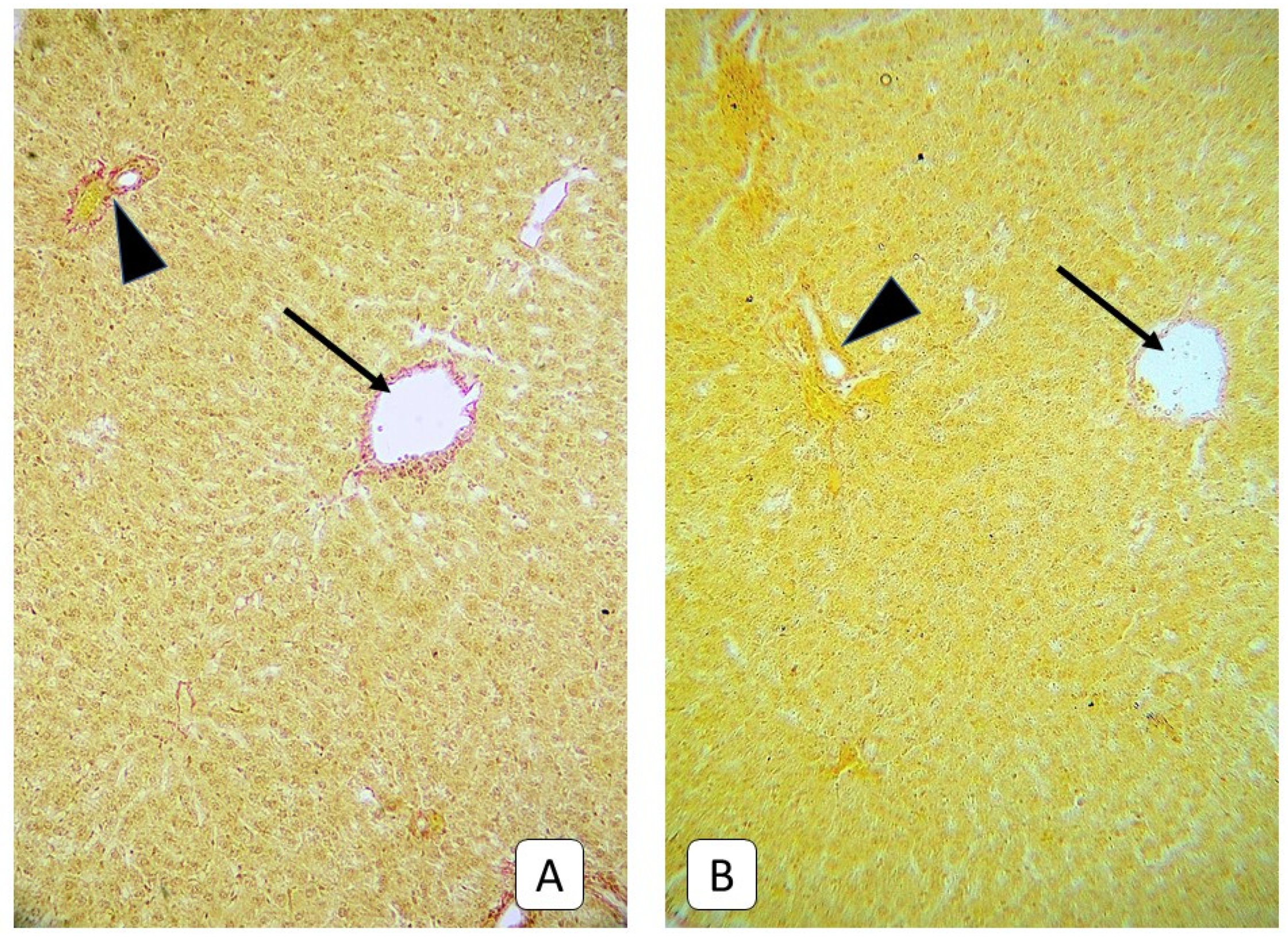
3.5. Histological Examinations of the Livers of Rats on the Fourteenth and Twenty-First Days after Trauma
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lumba-Brown, A.; Yeates, K.O.; Sarmiento, K.; Breiding, M.J.; Haegerich, T.M.; Gioia, G.A.; Turner, M.; Benzel, E.C.; Suskauer, S.J.; Giza, C.C.; et al. Centers for Disease Control and Prevention Guideline on the Diagnosis and Management of Mild Traumatic Brain Injury Among Children. JAMA Pediatr. 2018, 172, e182853. [Google Scholar] [CrossRef]
- Olsen, M.; Vik, A.; Lund Nilsen, T.I.; Uleberg, O.; Moen, K.G.; Fredriksli, O.; Lien, E.; Finnanger, T.G.; Skandsen, T. Incidence and mortality of moderate and severe traumatic brain injury in children: A ten year population-based cohort study in Norway. Eur. J. Paediatr. Neurol. 2019, 23, 500–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyder, A.A.; Wunderlich, C.A.; Puvanachandra, P.; Gururaj, G.; Kobusingye, O.C. The impact of traumatic brain injuries: A global perspective. Neuro. Rehabil. 2007, 22, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Maas, A.I.R.; Menon, D.K.; Adelson, P.D.; Angelic, N.; Bell, M.J.; Belli, A.; Bragge, P.; Brazinova, A.; Büki, A.; Chesnut, R.M.; et al. Traumatic brain injury: Integrated approaches to improve prevention, clinical care, and research. Lancet Neurol. 2017, 16, 987–1048. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.A.; Bell, J.M.; Breiding, M.J.; Xu, L. Traumatic brain injury-related Emergency department visits, hospitalizations, and deaths—United States, 2007 and 2013. MMWR SurveillSumm. 2017, 66, 1–16. [Google Scholar] [CrossRef]
- Araki, T.; Yokota, H.; Morita, A. Pediatric traumatic brain injury: Characteristic features, diagnosis, and management. Neurol. Med. Chir. (Tokyo) 2017, 57, 82–93. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-C.; Hsieh, P.-C.; Chen, C.P.C.; Hsieh, Y.-W.; Chung, C.-Y.; Lin, K.-L. Prevention, Protection Against Child Abuse, Neglect Study Group. Clinical characteristics and predictors of poor hospital discharge outcome for young children with abusive head trauma. J. Clin. Med. 2019, 8, 390. [Google Scholar] [CrossRef] [Green Version]
- Stefan, L.; Houston, R.; Pearson, G.; Edwards, R.; Hyde, P.; Maconochie, I.; Parslow, R.C.; Kemp, A. Epidemiology of children with head injury: A national overview. Arch. Dis. Child. 2016, 101, 527–532. [Google Scholar]
- Cassidy, J.D.; Carroll, L.J.; Peloso, P.M.; Borg, J.; von Holst, H.; Holm, L.; Kraus, J.; Coronado, V.G.; WHO Collaborating Centre Task Force on Mild Traumatic Brain Injury. Incidence, risk factors and prevention of mild traumatic brain injury: Results of the WHO Collaborating Centre Task Force on Mild Traumatic Brain Injury. J. Rehabil. Med. 2004, 43, 28–60. [Google Scholar] [CrossRef] [Green Version]
- Mild Traumatic Brain Injury Committee, American Congress of Rehabilitation Medicine, Head Injury Interdisciplinary Special Interest Group. Definition of mild traumatic brain injury. J. Head Trauma Rehabil. 1993, 8, 86–87. [Google Scholar] [CrossRef]
- Stuss, D.T. Traumatic brain injury: Relation to executive dysfunction and the frontal lobes. Curr. Opin. Neurol. 2011, 24, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.J.; Duka, T.; Stimpson, C.D.; Schapiro, S.J.; Baze, W.B.; McArthur, M.J.; Fobbs, A.J.; Sousa, A.M.; Sestan, N.; Wildman, D.E.; et al. Prolonged myelination in human neocortical evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 16480–16485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLegge, M.H.; Smoke, A. Neurodegeneration and inflammation. Nutr. Clin. Pract. 2008, 23, 35–41. [Google Scholar] [CrossRef]
- Mayer, C.L.; Huber, B.R.; Peskind, E. Traumatic brain injury, neuroinflammation, and posttraumatic headaches. Headache 2013, 53, 1523–1530. [Google Scholar] [PubMed] [Green Version]
- Anthony, D.C.; Couch, Y.; Losey, P.; Evans, M.C. The systemic response to brain injury and disease. Brain Behav. Immun. 2012, 26, 534–540. [Google Scholar] [CrossRef]
- Tobin, R.P.; Mukherjee, S.; Kain, J.M.; Rogers, S.K.; Henderson, S.K.; Motal, H.L.; Newell Rogers, M.K.; Shapiro, L.A. Traumatic brain injury causes selective, CD74-dependent peripheral lymphocyte activation that exacerbates neurodegeneration. Acta Neuropathol. Commun. 2014, 20, 143. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.J.; Gracon, A.S.; Ripsch, M.S.; Fisher, A.J.; Cheon, B.M.; Pandya, P.H.; Vittal, R.; Capitano, M.L.; Kim, Y.; Allette, Y.M.; et al. The HMGB1-RAGE axis mediates traumatic brain injury-induced pulmonary dysfunction in lung transplantation. Sci. Transl. Med. 2014, 6, 252ra124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodcock, T.; Morganti-Kossmann, M.C. The role of markers of inflammation in traumatic brain injury. Front. Neurol. 2013, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Kossmann, T.; Hans, V.H.; Imhof, H.G.; Stocker, R.; Grob, P.; Trentz, O.; Morganti-Kossmann, C. Intrathecal and serum interleukin-6 and the acute-phase response in patients with severe traumatic brain injuries. Shock 1995, 4, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Ramadori, G.; Christ, B. Cytokines and the hepatic acute-phase response. Semin. Liver Dis. 1999, 19, 141–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, S.J.; Anthony, D.C.; Oakley, F.; Carlsen, H.; Elsharkawy, A.M.; Blomhoff, R.; Mann, D.A. Hepatic nuclear factor kappa B regulates neutrophil recruitment to the injured brain. J. Neuropathol. Exp. Neurol. 2008, 67, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Villapol, S. Consequences of hepatic damage after traumatic brain injury: Current outlook and potential therapeutic targets. Neural. Regen. Res. 2016, 11, 226–227. [Google Scholar] [CrossRef]
- Meretskyi, V.M. Method for Simulation of Traumatic Brain Injury. Ukrainian Patent No. 74935, 12 November 2012. Available online: http://uapatents.com/5-74935-sposib-modelyuvannya-cherepno-mozkovo-travmi.html (accessed on 30 January 2019).
- Yelsky, V.N.; Zyablitsev, S.V. Simulation of Traumatic Brain Injury; New Worls: Donetsk, Ukrain, 2008; p. 139. [Google Scholar]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. BiophotonicsInt. 2004, 11, 36–42. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 28, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Jensen, E.B.; Gundersen, H.J.; Osterby, R. Determination of membrane thickness distribution from orthogonal intercepts. J. Microsc. 1979, 115, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Coan, P.M.; Ferguson-Smith, A.C.; Burton, G.J. Developmental dynamics of the definitive mouse placenta assessed by stereology. Biol. Reprod. 2004, 70, 1806–1813. [Google Scholar] [CrossRef]
- Howard, V.; Reed, M. Unbiased Stereology: Three-Dimensional Measurement in Microscopy; Garland Science: London, UK, 2004; p. 296. [Google Scholar]
- Santarsiero, M.; Kumar, R.G.; Kochanek, P.M.; Berga, S.; Wagner, A.K. Variable neuroendocrine-immune dysfunction in individuals with unfavorable outcome after severe traumatic brain injury. Brain Behav. Immun. 2015, 45, 15–27. [Google Scholar] [CrossRef] [Green Version]
- McKee, C.A.; Lukens, J.R. Emerging roles for the immune system in traumatic brain injury. Front. Immunol. 2016, 7, 556. [Google Scholar] [CrossRef] [Green Version]
- Hazeldine, J.; Lord, J.M.; Belli, A. Traumatic brain injury and peripheral immune suppression: Primer and prospectus. Front. Neurol. 2015, 6, 235. [Google Scholar] [CrossRef] [Green Version]
- Algattas, H.; Huang, J.H. Traumatic brain injury pathophysiology and treatments: Early, intermediate, and late phases post-injury. Int. J. Mol. Sci. 2013, 15, 309–341. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, K. Traumatic brain injury: Pathophysiology for neurocritical care. J. Intensive Care 2016, 4, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, L.; McClain, C.J.; Gillespie, M.; Young, B. Cytokines and metabolic dysfunction after severe head injury. J. Neurotrauma 1994, 11, 447–472. [Google Scholar] [CrossRef] [PubMed]
- Meretskyi, V. Structural and morphological disorders of internal organs of rats with craniocerebral injury and diabetes mellitus. Hosp. Surg. 2012, 4, 56–60. [Google Scholar]
- Brienza, N.; Delfino, L.; Cannella, G.; Diele, C.; Bruno, F.; Fiore, T. Jaundice in critical illness: Promoting factors of a concealed reality. Intensive. Care Med. 2006, 32, 267–274. [Google Scholar] [CrossRef]
- Gruber, A.; Reinprecht, A.; Illievich, U.M.; Fitzgerald, R.; Dietrich, W.; Czech, T.; Richling, B. Extracerebral organ dysfunction and neurologic outcome after aneurysmal subarachnoid hemorrhage. Crit. Care Med. 1999, 27, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Harbrecht, B.G.; Zenati, M.S.; Doyle, H.R.; McMichael, J.; Townsend, R.N.; Clancy, K.D.; Peitzman, A.B. Hepatic dysfunction increases length of stay and risk of death after injury. J. Trauma 2002, 53, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Romeo, R.D. The metamorphosis of adolescent hormonal stress reactivity: A focus on animal models. Front. Neuroendocrinol. 2018, 49, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, D.M. Stress and the developing limbic-hypothalamic-pituitary-adrenal axis. Psychoneuroendocrinology 1998, 23, 663–700. [Google Scholar] [CrossRef]
- De Kloet, E.R.; Rosenfeld, P.; Van Eekelen, J.A.; Sutanto, W.; Levine, S. Stress, glucocorticoids and development. Prog. Brain. Res. 1988, 73, 101–120. [Google Scholar]
- Espelid, S.; Løkken, G.B.; Steiro, K.; Bøgwald, J. Effects of cortisoland stress on the immune system in Atlantic Salmon (Salmo salar L.). Fish Shellfish Immunol. 1996, 6, 95–110. [Google Scholar] [CrossRef]
- Roth, J.A. Cortisol as mediator of stress-associated immunosuppression in cattle. In Animal Stress; Moberg, G.P., Ed.; Springer: New York, NY, USA, 1985; pp. 225–2433. [Google Scholar]
- Ortega, E. Neuroendocrine mediators in the modulation of phagocytosis by exercise: Physiological implications. Exerc. Immunol. Rev. 2003, 9, 70–93. [Google Scholar] [PubMed]
- Greenstein, A.; Munster, P.N.; Sachdev, J.; Fleming, G.; Grauer, A.; Shepherd, S. Impact of relacorilant, a selective glucocorticoid receptor antagonist, on the immunosuppressive effects of endogenous cortisol. J. Clin. Oncol. 2020, 38, 3091. [Google Scholar] [CrossRef]
- Cottrell, E.C.; Seckl, J.R. Prenatal stress, glucocorticoids and the programming of adult disease. Front. Behav. Neurosci. 2009, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Tsoory, M.; Richter-Levin, G. Learning under stress in the adult rat is differentially affected by ‘juvenile’ or ‘adolescent’ stress. Int. J. Neuropsychopharmacol. 2006, 9, 713–728. [Google Scholar] [CrossRef] [Green Version]
- Semenova, Z.B.; Melnikov, A.V.; Savin, I.A.; Lekmanov, A.U.; Khachatryan, V.A.; Gorelyshev, S.K. Recommendations for treatment of children with craniocerebral trauma. Russ. J. Pediatric Surg. Anesth. Intensive Care 2016, 6, 112–127. [Google Scholar]
- Robba, C.; Bonatti, G.; Pelosi, P.; Citerio, G. Extracranial complications after traumatic brain injury: Targeting the brain and the body. Curr. Opin. Crit. Care 2020, 26, 137–146. [Google Scholar] [CrossRef]
- Gundappa, P. Extracranial complications of traumatic brain injury: Pathophysiology—A review. J. Neuroanaesth. Crit. Care 2019, 6, 200–212. [Google Scholar] [CrossRef] [Green Version]
- Kemp, C.D.; Johnson, J.C.; Riordan, W.P.; Cotton, B.A. How we die: The impact of nonneurologic organ dysfunction after severe traumatic brain injury. Am. Surg. 2008, 74, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Narad, M.E.; Riemersma, J.; Wade, S.L.; Smith-Paine, J.; Morrison, P.; Taylor, H.G.; Yeates, K.O.; Kurowski, B.G. Impact of secondary adhd on long-term outcomes after early childhood traumatic brain injury. J. Head Trauma Rehabil. 2020, 35, E271–E279. [Google Scholar] [CrossRef]
- Carroll, E.L.; Manktelow, A.E.; Outtrim, J.G.; Chatfield, D.; Forsyth, F.; Hutchinson, P.J.A.; Tenovuo, O.; Posti, J.P.; Wilson, L.; Sahakian, B.J.; et al. Influence of concomitant extracranial injury on functional and cognitive recovery from mild versus moderateto severe traumatic brain injury. J. Head Trauma Rehabil. 2020, 35, E513–E523. [Google Scholar] [CrossRef] [PubMed]
- Landre, N.; Poppe, C.J.; Davis, N.; Schmaus, B.; Hobbs, S.E. Cognitive functioning and postconcussive symptoms in trauma patients with and without mild TBI. Arch. Clin. Neuropsychol. 2006, 21, 255–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, H.S.; Hanten, G.; Roberson, G.; Li, X.; Ewing-Cobbs, L.; Dennis, M.; Chapman, S.; Max, J.E.; Hunter, J.; Schachar, R.; et al. Prediction of cognitive sequelae based on abnormal computed tomography findings in children following mild traumatic brain injury. J. Neurosurg. Pediatr. 2008, 1, 461–470. [Google Scholar] [CrossRef]
- Vos, P.E.; Alekseenko, Y.; Battistin, L.; Ehler, E.; Gerstenbrand, F.; Muresanu, D.F.; Potapov, A.; Stepan, C.A.; Traubner, P.; Vecsei, L.; et al. Mild traumatic brain injury. Eur. J. Neurol. 2012, 19, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, P. The laboratory rat: Relating its age with human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group IA [μm] | Numbers of Measurements | p | Group IIA [μm] | Numbers of Measurements | p | |
|---|---|---|---|---|---|---|
| Control group | 4.07, (3.56–4.68) | 208 | 3.43, (3.06–4.09) | 131 | ||
| Study group | Group IB [μm] | Group IIB [μm] | ||||
| Day 1 | 6.03, (4.44–8.69) | 275 | <0.001 * | 3.72, (2.88–4.90) | 188 | 0.61 * |
| Day 3 | 9.40, (7.62–11.82) | 321 | <0.001 * <0.001 # | 2.38, (1.92–2.85) | 142 | <0.001 * <0.001 # |
| Day 5 | 10.17, (7.91–12.67) | 246 | <0.001 * 0.53 # | 2.64, (2.12–3.67) | 126 | <0.001 * 0.01 # |
| Day 7 | 8.37, (6.16–11.01) | 244 | <0.001 * <0.001 # | 2.60, (2.14–3.44) | 111 | <0.001 * 0.99 # |
| Day 14 | 9.53, (7.29–12.71) | 277 | <0.001 * 0.001 # | 3.16, (2.61–3.93) | 111 | 0.052 * 0.052 # |
| Day 21 | 7.78, (5.81–9.17) | 205 | <0.001 * <0.001 # | 3.19, (2.42–3.98) | 111 | 0.052 * 0.99 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prus, R.; Appelhans, O.; Logash, M.; Pokotylo, P.; Nowicki, G.J.; Ślusarska, B. A Histological and Morphometric Assessment of the Adult and Juvenile Rat Livers after Mild Traumatic Brain Injury. Cells 2021, 10, 1121. https://doi.org/10.3390/cells10051121
Prus R, Appelhans O, Logash M, Pokotylo P, Nowicki GJ, Ślusarska B. A Histological and Morphometric Assessment of the Adult and Juvenile Rat Livers after Mild Traumatic Brain Injury. Cells. 2021; 10(5):1121. https://doi.org/10.3390/cells10051121
Chicago/Turabian StylePrus, Ruslan, Olena Appelhans, Maksim Logash, Petro Pokotylo, Grzegorz Józef Nowicki, and Barbara Ślusarska. 2021. "A Histological and Morphometric Assessment of the Adult and Juvenile Rat Livers after Mild Traumatic Brain Injury" Cells 10, no. 5: 1121. https://doi.org/10.3390/cells10051121