
Adaptive Subsets Limit the Anti-Tumoral NK-Cell Activity in Hepatocellular Carcinoma

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Cohort
2.2. PBMC Isolation
2.3. Isolation of Lymphocytes from Tissue Samples
2.4. HCMV Status
2.5. Cell Lines
2.6. Assessment of NK-Cell Function
2.7. Multiparametric Flow Cytometry
2.8. Statistics
3. Results
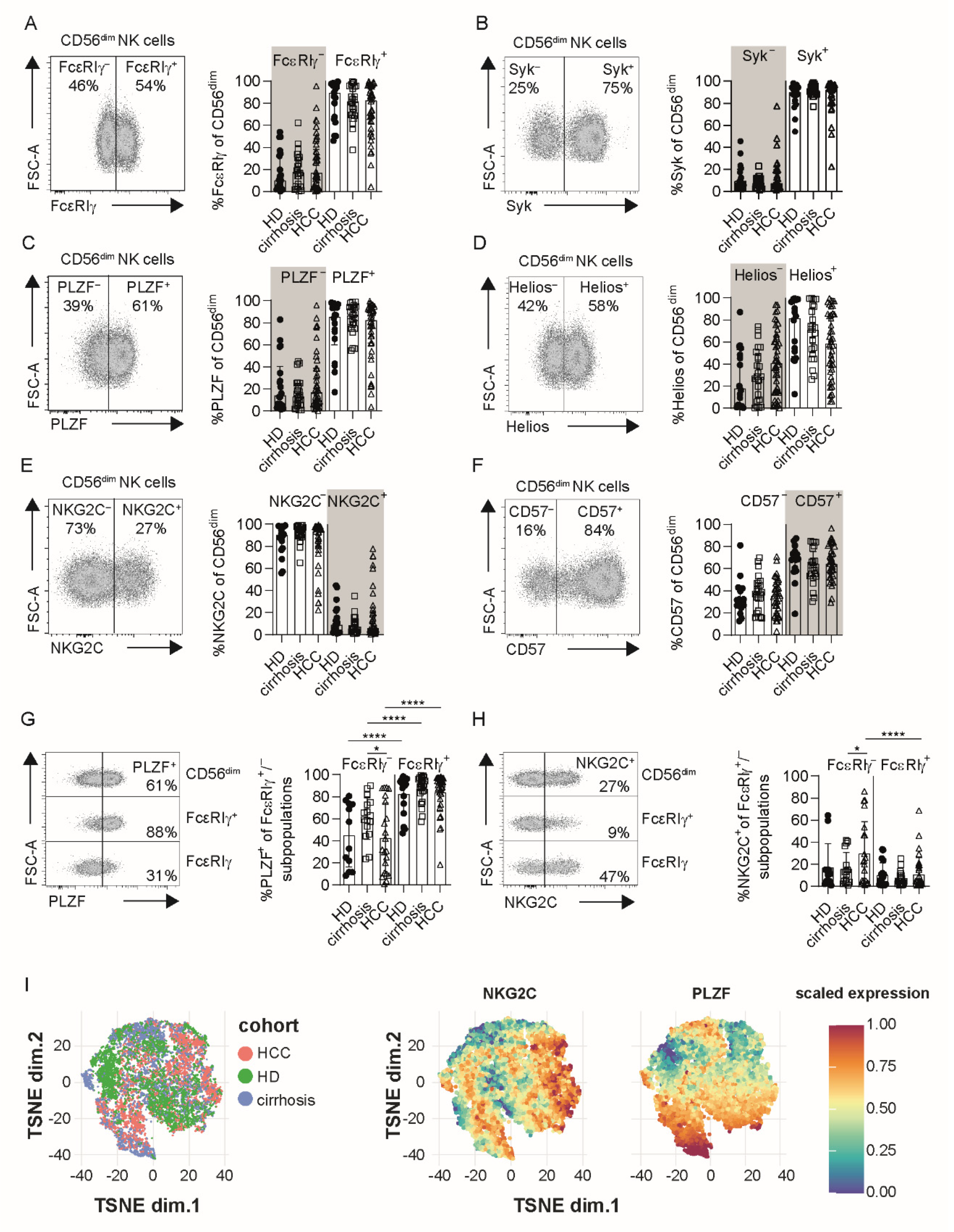
3.1. Profiles of Adaptive NK Cells Are Conserved in HCC
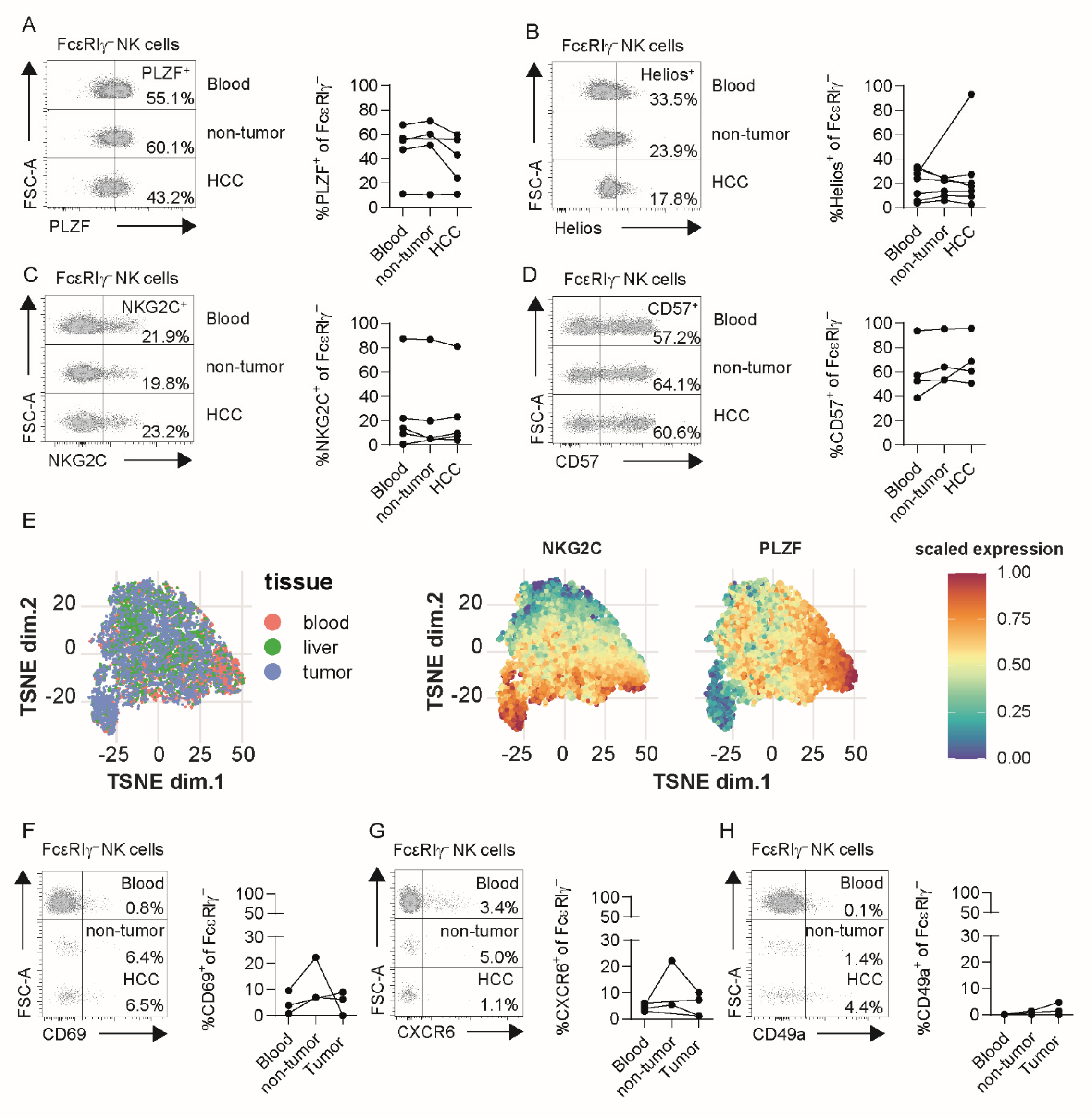
3.2. Adaptive NK Cells in Tumor and Non-Tumor Liver Tissue Exhibit Conserved Profiles
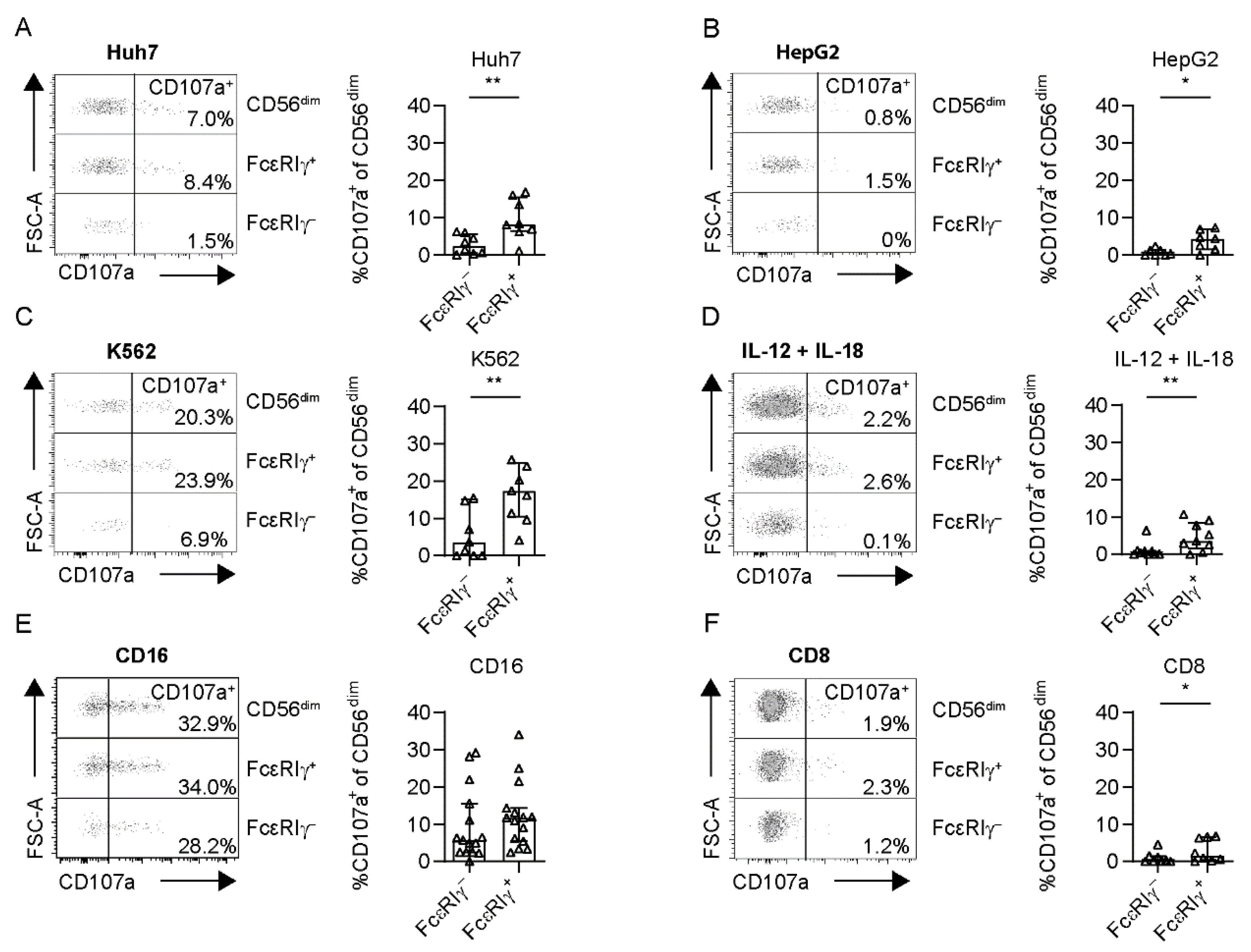
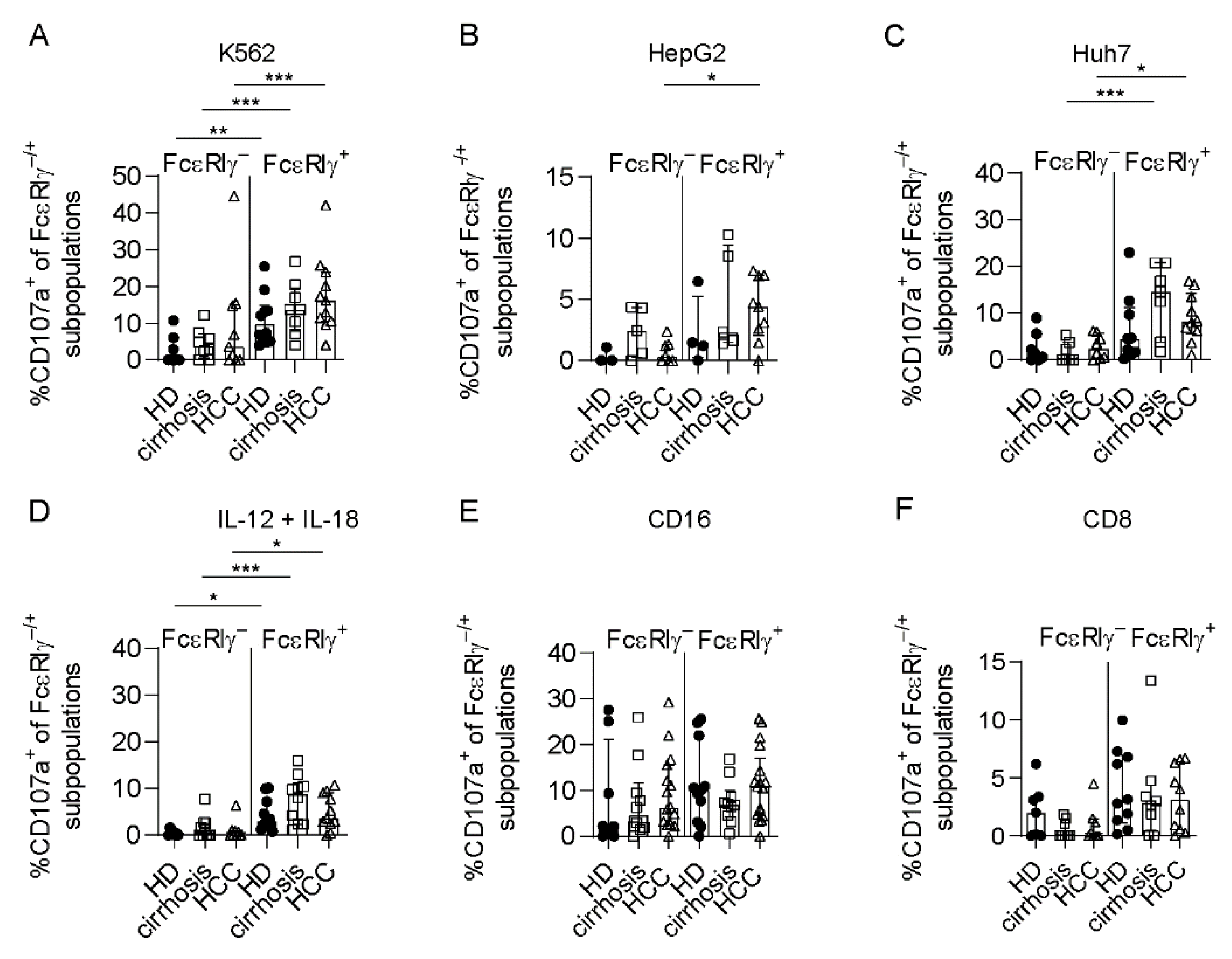
3.3. Limited Anti-Tumoral Activity of FcεRIγ− Adaptive NK Cells
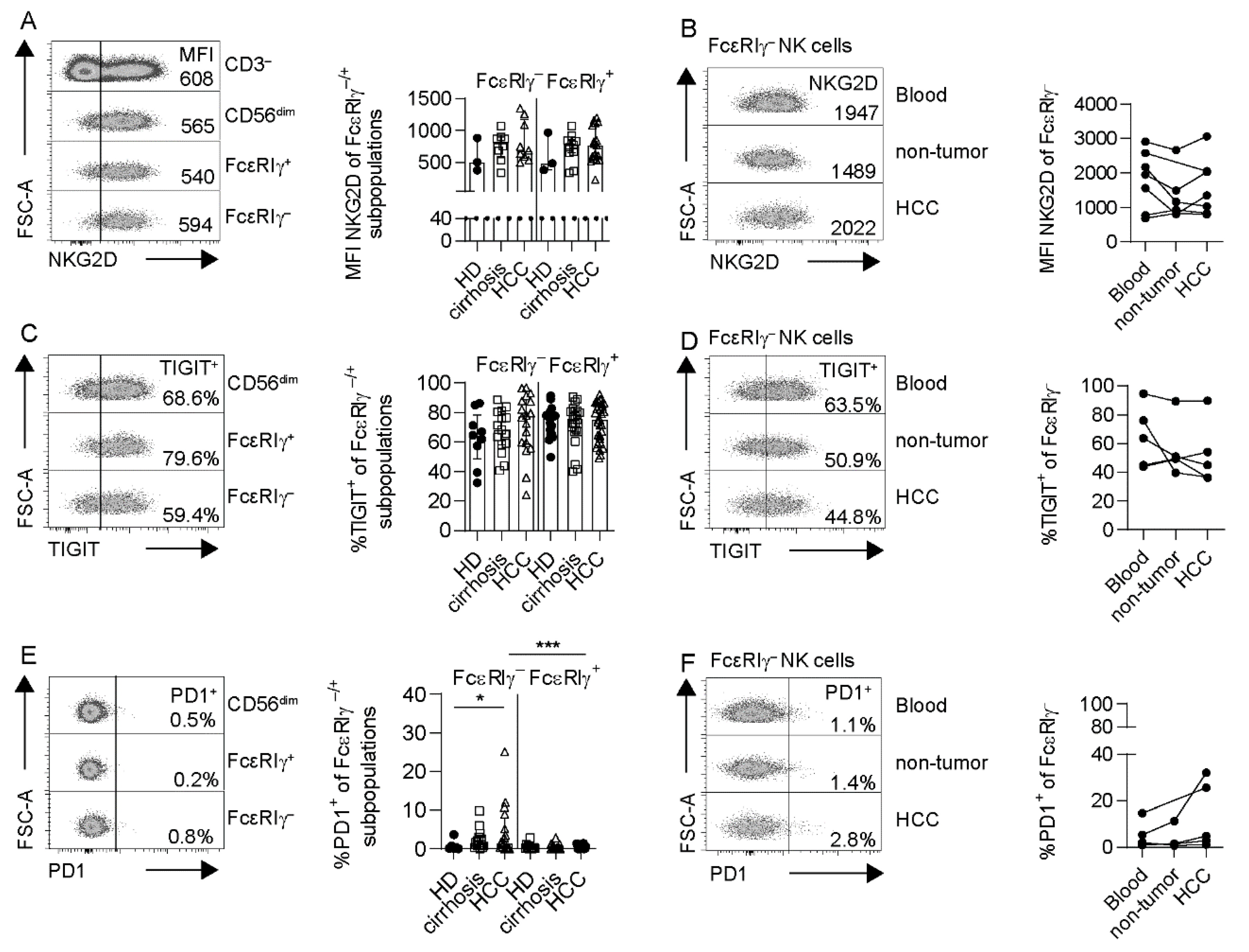
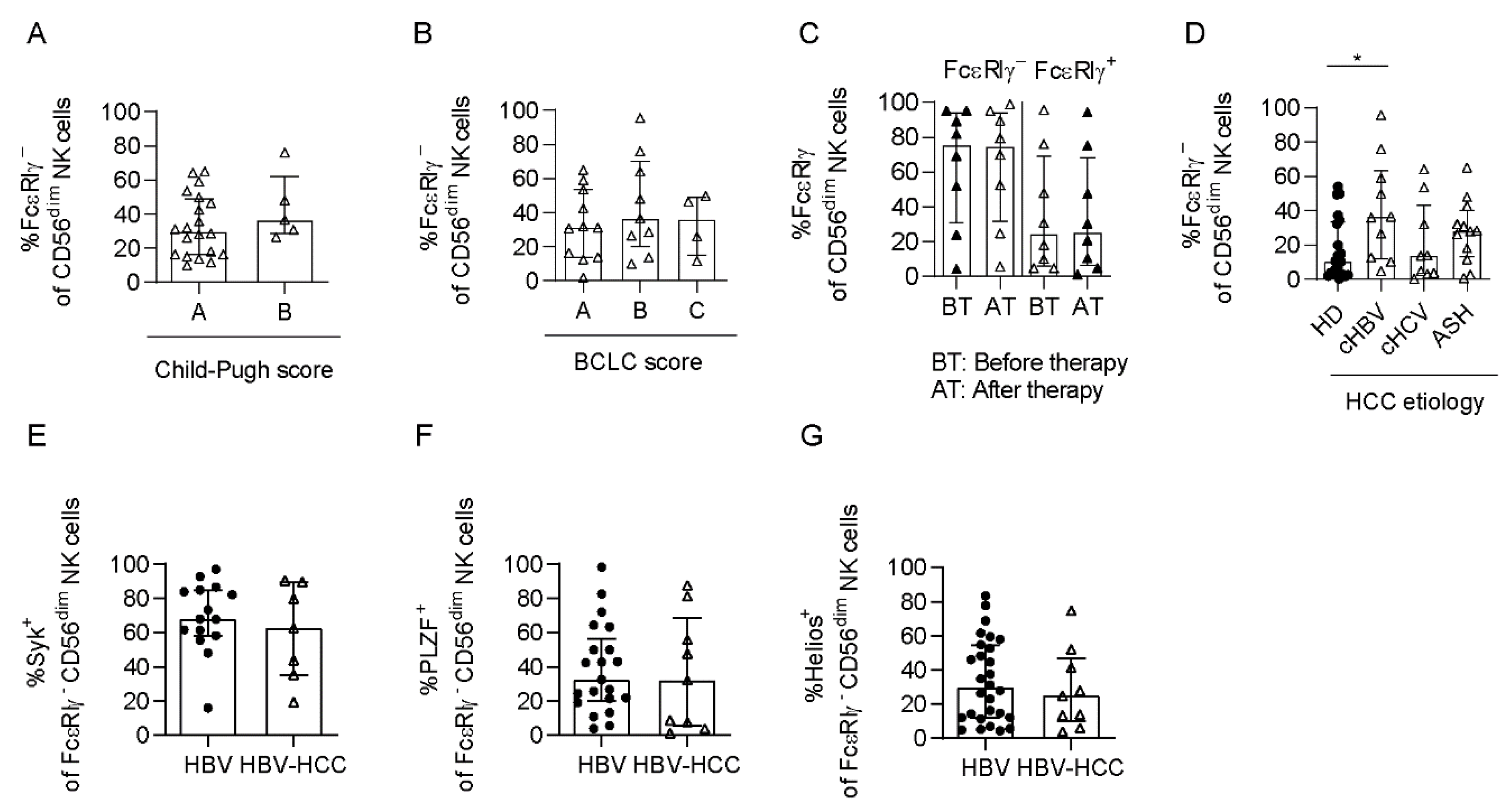
3.4. Adaptive NK Cells Are Not Specifically Impaired in HCC
3.5. Adaptive NK Cells Are Expanded in HCMV+ HBV-Associated HCC Patients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balogh, J.; Victor, D.I.; Asham, E.H.; Burroughs, S.G.; Boktour, M.; Saharia, A.; Li, X.; Ghobrial, R.M.; Monsour, H.P., Jr. Hepatocellular carcinoma: A review. J. Hepatocell Carcinoma 2016, 3, 41–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foerster, F.; Hess, M.; Gerhold-Ay, A.; Marquardt, J.U.; Becker, D.; Galle, P.R.; Schuppan, D.; Binder, H.; Bockamp, E. The immune contexture of hepatocellular carcinoma predicts clinical outcome. Sci. Rep. 2018, 8, 5351. [Google Scholar] [CrossRef] [PubMed]
- Ringelhan, M.; Pfister, D.; O’Connor, T.; Pikarsky, E.; Heikenwalder, M. The immunology of hepatocellular carcinoma. Nat. Immunol. 2018, 19, 222–232. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. EASL clinical practice guidelines: Management of hepatocellular carcinoma. J. Hepatol. 2018, 69, 182–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachdeva, M.; Chawla, Y.K.; Arora, S.K. Immunology of hepatocellular carcinoma. World J. Hepatol. 2015, 7, 2080–2090. [Google Scholar] [CrossRef]
- Flecken, T.; Schmidt, N.; Hild, S.; Gostick, E.; Drognitz, O.; Zeiser, R.; Schemmer, P.; Bruns, H.; Eiermann, T.; Price, D.A.; et al. Immunodominance and functional alterations of tumor-associated antigen-specific cd8+ t-cell responses in hepatocellular carcinoma. Hepatology 2014, 59, 1415–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taketomi, A.; Shimada, M.; Shirabe, K.; Kajiyama, K.; Gion, T.; Sugimachi, K. Natural killer cell activity in patients with hepatocellular carcinoma. Cancer 1998, 83, 58–63. [Google Scholar] [CrossRef]
- Huo, J.; Wu, L.; Zang, Y. Development and validation of a novel immune-gene pairs prognostic model associated with ctnnb1 alteration in hepatocellular carcinoma. Med. Sci. Monit. 2020, 26, e925494. [Google Scholar] [CrossRef]
- Yokoyama, W.M.; Kim, S. How do natural killer cells find self to achieve tolerance? Immunity 2006, 24, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Ljunggren, H.-G.; Kärre, K. In search of the ‘missing self’: Mhc molecules and nk cell recognition. Immunol. Today 1990, 11, 237–244. [Google Scholar] [CrossRef]
- Cerwenka, A.; Lanier, L.L. Nkg2d ligands: Unconventional mhc class i-like molecules exploited by viruses and cancer. Tissue Antigens 2003, 61, 335–343. [Google Scholar] [CrossRef]
- Gasser, S.; Orsulic, S.; Brown, E.J.; Raulet, D.H. The DNA damage pathway regulates innate immune system ligands of the nkg2d receptor. Nature 2005, 436, 1186–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biron, C.A.; Nguyen, K.B.; Pien, G.C.; Cousens, L.P.A.; Salazar-Mather, T.P. Natural killer cells in antiviral defense: Function and regulation by innate cytokines. Annu. Rev. Immunol. 1999, 17, 189–220. [Google Scholar] [CrossRef] [PubMed]
- Fehniger, T.A.; Shah, M.H.; Turner, M.J.; VanDeusen, J.B.; Whitman, S.P.; Cooper, M.A.; Suzuki, K.; Wechser, M.; Goodsaid, F.; Caligiuri, M.A. Differential cytokine and chemokine gene expression by human nk cells following activation with il-18 or il-15 in combination with il-12: Implications for the innate immune response. J. Immunol. 1999, 162, 4511. [Google Scholar]
- Young, H.A.; Ortaldo, J. Cytokines as critical co-stimulatory molecules in modulating the immune response of natural killer cells. Cell Res. 2006, 16, 20–24. [Google Scholar] [CrossRef]
- Cassatella, M.A.; Anegón, I.; Cuturi, M.C.; Griskey, P.; Trinchieri, G.; Perussia, B. Fc gamma r(cd16) interaction with ligand induces ca2+ mobilization and phosphoinositide turnover in human natural killer cells. Role of ca2+ in fc gamma r(cd16)-induced transcription and expression of lymphokine genes. J. Exp. Med. 1989, 169, 549. [Google Scholar] [CrossRef] [Green Version]
- Morvan, M.G.; Lanier, L.L. Nk cells and cancer: You can teach innate cells new tricks. Nat. Rev. Cancer 2015, 16, 7. [Google Scholar] [CrossRef] [PubMed]
- Norris, S.; Collins, C.; Doherty, D.G.; Smith, F.; McEntee, G.; Traynor, O.; Nolan, N.; Hegarty, J.; O’Farrelly, C. Resident human hepatitis lymphocytes are phenotypically different from circulating lymphocytes. J. Hepatol. 1998, 28, 84–90. [Google Scholar] [CrossRef]
- Doherty, D.G.; Norris, S.; Madrigal-Estebas, L.; McEntee, G.; Traynor, O.; Hegarty, J.E.; O’Farrelly, C. The Human Liver Contains Multiple Populations of NK Cells, T Cells, and CD3+CD56+ Natural T Cells with Distinct Cytotoxic Activities and Th1, Th2, and Th0 Cytokine Secretion Patterns. J. Immunol. 1999, 163, 2314. [Google Scholar] [PubMed]
- Björkström, N.K.; Ljunggren, H.-G.; Michaëlsson, J. Emerging insights into natural killer cells in human peripheral tissues. Nat. Rev. Immunol. 2016, 16, 310. [Google Scholar] [CrossRef]
- Harmon, C.; Robinson, M.W.; Fahey, R.; Whelan, S.; Houlihan, D.D.; Geoghegan, J.; O’Farrelly, C. Tissue-resident eomeshi t-betlo cd56bright nk cells with reduced proinflammatory potential are enriched in the adult human liver. Eur. J. Immunol. 2016, 46, 2111–2120. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Scott, J.M.; Hwang, I.; Kim, S. Cutting edge: Antibody-dependent memory-like nk cells distinguished by fcrγ deficiency. J. Immunol. 2013, 190, 1402. [Google Scholar] [CrossRef] [Green Version]
- Easom, N.J.W.; Stegmann, K.A.; Swadling, L.; Pallett, L.J.; Burton, A.R.; Odera, D.; Schmidt, N.; Huang, W.-C.; Fusai, G.; Davidson, B.; et al. Il-15 overcomes hepatocellular carcinoma-induced nk cell dysfunction. Front. Immunol. 2018, 9, 1009. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Zhang, Z.; Zhou, L.; Wang, H.; Fu, J.; Zhang, S.; Shi, M.; Zhang, H.; Yang, Y.; Wu, H.; et al. Functional impairment in circulating and intrahepatic nk cells and relative mechanism in hepatocellular carcinoma patients. Clin. Immunol. 2008, 129, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Cariani, E.; Pilli, M.; Barili, V.; Porro, E.; Biasini, E.; Olivani, A.; Dalla Valle, R.; Trenti, T.; Ferrari, C.; Missale, G. Natural killer cells phenotypic characterization as an outcome predictor of hcv-linked hcc after curative treatments. OncoImmunology 2016, 5, e1154249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horowitz, A.; Strauss-Albee, D.M.; Leipold, M.; Kubo, J.; Nemat-Gorgani, N.; Dogan, O.C.; Dekker, C.L.; Mackey, S.; Maecker, H.; Swan, G.E.; et al. Genetic and environmental determinants of human nk cell diversity revealed by mass cytometry. Sci. Transl. Med. 2013, 5, 208ra145. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, R.; Hintzen, G.; Kemper, A.; Beul, K.; Kempf, S.; Behrens, G.; Sykora, K.-W.; Schmidt, R.E. Cd56bright cells differ in their kir repertoire and cytotoxic features from cd56dim nk cells. Eur. J. Immunol. 2001, 31, 3121–3126. [Google Scholar] [CrossRef]
- Hudspeth, K.; Donadon, M.; Cimino, M.; Pontarini, E.; Tentorio, P.; Preti, M.; Hong, M.; Bertoletti, A.; Bicciato, S.; Invernizzi, P.; et al. Human liver-resident cd56bright/cd16neg nk cells are retained within hepatic sinusoids via the engagement of ccr5 and cxcr6 pathways. J. Autoimmun. 2016, 66, 40–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegmann, K.A.; Robertson, F.; Hansi, N.; Gill, U.; Pallant, C.; Christophides, T.; Pallett, L.J.; Peppa, D.; Dunn, C.; Fusai, G.; et al. Cxcr6 marks a novel subset of t-betloeomeshi natural killer cells residing in human liver. Sci. Rep. 2016, 6, 26157. [Google Scholar] [CrossRef]
- Nagler, A.; Lanier, L.L.; Cwirla, S.; Phillips, J.H. Comparative studies of human fcriii-positive and negative natural killer cells. J. Immunol. 1989, 143, 3183. [Google Scholar]
- Schlums, H.; Cichocki, F.; Tesi, B.; Theorell, J.; Beziat, V.; Holmes, T.D.; Han, H.; Chiang, S.C.; Foley, B.; Mattsson, K.; et al. Cytomegalovirus infection drives adaptive epigenetic diversification of nk cells with altered signaling and effector function. Immunity 2015, 42, 443–456. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Zhang, T.; Hwang, I.; Kim, A.; Nitschke, L.; Kim, M.; Scott, J.M.; Kamimura, Y.; Lanier, L.L.; Kim, S. Epigenetic modification and antibody-dependent expansion of memory-like nk cells in human cytomegalovirus-infected individuals. Immunity 2015, 42, 431–442. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Vergès, S.; Milush, J.M.; Schwartz, B.S.; Pando, M.J.; Jarjoura, J.; York, V.A.; Houchins, J.P.; Miller, S.; Kang, S.-M.; Norris, P.J.; et al. Expansion of a unique cd57+nkg2chi natural killer cell subset during acute human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 14725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumá, M.; Angulo, A.; Vilches, C.; Gómez-Lozano, N.; Malats, N.; López-Botet, M. Imprint of human cytomegalovirus infection on the nk cell receptor repertoire. Blood 2004, 104, 3664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, I.; Zhang, T.; Scott, J.M.; Kim, A.R.; Lee, T.; Kakarla, T.; Kim, A.; Sunwoo, J.B.; Kim, S. Identification of human nk cells that are deficient for signaling adaptor fcrγ and specialized for antibody-dependent immune functions. Int. Immunol. 2012, 24, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Schuch, A.; Zecher, B.F.; Müller, P.A.; Correia, M.P.; Daul, F.; Rennert, C.; Tauber, C.; Schlitt, K.; Boettler, T.; Neumann-Haefelin, C.; et al. Nk-cell responses are biased towards cd16-mediated effector functions in chronic hepatitis b virus infection. J. Hepatol. 2019, 70, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Marquardt, N.; Béziat, V.; Nyström, S.; Hengst, J.; Ivarsson, M.A.; Kekäläinen, E.; Johansson, H.; Mjösberg, J.; Westgren, M.; Lankisch, T.O.; et al. Cutting edge: Identification and characterization of human intrahepatic cd49a+nk cells. J. Immunol. 2015, 194, 1402756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesi, B.; Schlums, H.; Cichocki, F.; Bryceson, Y.T. Epigenetic regulation of adaptive nk cell diversification. Trends Immunol. 2016, 37, 451–461. [Google Scholar] [CrossRef]
- Björkström, N.K.; Lindgren, T.; Stoltz, M.; Fauriat, C.; Braun, M.; Evander, M.; Michaëlsson, J.; Malmberg, K.-J.; Klingström, J.; Ahlm, C.; et al. Rapid expansion and long-term persistence of elevated nk cell numbers in humans infected with hantavirus. J. Exp. Med. 2011, 208, 13. [Google Scholar] [CrossRef]
- Cichocki, F.; Cooley, S.; Davis, Z.; DeFor, T.E.; Schlums, H.; Zhang, B.; Brunstein, C.G.; Blazar, B.R.; Wagner, J.; Diamond, D.J.; et al. Cd56dim cd57+nkg2c+ nk cell expansion is associated with reduced leukemia relapse after reduced intensity hct. Leukemia 2015, 30, 456. [Google Scholar] [CrossRef]
- Lepiller, Q.; Tripathy, M.K.; Di Martino, V.; Kantelip, B.; Herbein, G. Increased hcmv seroprevalence in patients with hepatocellular carcinoma. Virol. J. 2011, 8, 485. [Google Scholar] [CrossRef] [Green Version]
- Lachmann, R.; Loenenbach, A.; Waterboer, T.; Brenner, N.; Pawlita, M.; Michel, A.; Thamm, M.; Poethko-Müller, C.; Wichmann, O.; Wiese-Posselt, M. Cytomegalovirus (cmv) seroprevalence in the adult population of germany. PLoS ONE 2018, 13, e0200267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijaya, R.S.; Read, S.A.; Selvamani, S.P.; Schibeci, S.; Azardaryany, M.K.; Ong, A.; van der Poorten, D.; Lin, R.; Douglas, M.W.; George, J.; et al. Hepatitis c virus eradication with interferon free, daa-based therapy results in klrg1+, hepatitis c virus-specific memory natural killer cells. J. Infect. Dis. 2021, 223, 1183–1195. [Google Scholar] [CrossRef] [PubMed]
- Wijaya, R.S.; Read, S.A.; Schibeci, S.; Eslam, M.; Azardaryany, M.K.; El-Khobar, K.; van der Poorten, D.; Lin, R.; Yuen, L.; Lam, V.; et al. Klrg1+ natural killer cells exert a novel antifibrotic function in chronic hepatitis b. J. Hepatol. 2019, 71, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Wijaya, R.S.; Read, S.A.; Truong, N.R.; Han, S.; Chen, D.; Shahidipour, H.; Fewings, N.L.; Schibeci, S.; Azardaryany, M.K.; Parnell, G.P.; et al. Hbv vaccination and hbv infection induces hbv-specific natural killer cell memory. Gut 2021, 70, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Yu, H.T.; Hwang, I.; Park, S.; Park, S.H.; Kim, S.; Shin, E.C. Phenotypic and functional analysis of human nk cell subpopulations according to the expression of fcepsilonrigamma and nkg2c. Front. Immunol. 2019, 10, 2865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Scott, J.M.; Langguth, E.; Chang, H.; Park, P.H.; Kim, S. Fcrgamma gene editing reprograms conventional nk cells to display key features of adaptive human nk cells. iScience 2020, 23, 101709. [Google Scholar] [CrossRef]
- Franco, A.; Barnaba, V.; Natali, P.; Balsano, C.; Musca, A.; Balsano, F. Expression of class i and class ii major histocompatibility complex antigens on human hepatocytes. Hepatology 1988, 8, 449–454. [Google Scholar] [CrossRef]
- Oh, J.S.; Ali, A.K.; Kim, S.; Corsi, D.J.; Cooper, C.L.; Lee, S.H. Nk cells lacking fcepsilonrigamma are associated with reduced liver damage in chronic hepatitis c virus infection. Eur. J. Immunol. 2016, 46, 1020–1029. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Bi, J.; Zheng, X.; Chen, Y.; Wang, H.; Wu, W.; Wang, Z.; Wu, Q.; Peng, H.; Wei, H.; et al. Blockade of the checkpoint receptor tigit prevents nk cell exhaustion and elicits potent anti-tumor immunity. Nat. Immunol. 2018, 19, 723–732. [Google Scholar] [CrossRef] [PubMed]
- André, P.; Denis, C.; Soulas, C.; Bourbon-Caillet, C.; Lopez, J.; Arnoux, T.; Bléry, M.; Bonnafous, C.; Gauthier, L.; Morel, A.; et al. Anti-nkg2a mab is a checkpoint inhibitor that promotes anti-tumor immunity by unleashing both t and nk cells. Cell 2018, 175, 1731–1743. [Google Scholar] [CrossRef] [Green Version]
- Pesce, S.; Greppi, M.; Tabellini, G.; Rampinelli, F.; Parolini, S.; Olive, D.; Moretta, L.; Moretta, A.; Marcenaro, E. Identification of a subset of human natural killer cells expressing high levels of programmed death 1: A phenotypic and functional characterization. J. Allergy Clin. Immunol. 2017, 139, 335–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Cheng, Y.; Xu, Y.; Wang, Z.; Du, X.; Li, C.; Peng, J.; Gao, L.; Liang, X.; Ma, C. Increased expression of programmed cell death protein 1 on nk cells inhibits nk-cell-mediated anti-tumor function and indicates poor prognosis in digestive cancers. Oncogene 2017, 36, 6143–6153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merino, A.M.; Zhang, B.; Dougherty, P.R.; Luo, X.; Wang, J.; Blazar, B.R.; Miller, J.S.; Cichocki, F. Chronic stimulation drives human nk cell dysfunction and epigenetic reprograming. J. Clin. Investig. 2019, 129, 3770–3785. [Google Scholar] [CrossRef] [Green Version]
- Lang, P.A.; Lang, K.S.; Xu, H.C.; Grusdat, M.; Parish, I.A.; Recher, M.; Elford, A.R.; Dhanji, S.; Shaabani, N.; Tran, C.W.; et al. Natural killer cell activation enhances immune pathology and promotes chronic infection by limiting cd8+ t-cell immunity. Proc. Natl. Acad. Sci. USA 2012, 109, 1210–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duhan, V.; Hamdan, T.A.; Xu, H.C.; Shinde, P.; Bhat, H.; Li, F.; Al-Matary, Y.; Haussinger, D.; Bezgovsek, J.; Friedrich, S.K.; et al. Nk cell-intrinsic fcepsilonrigamma limits cd8+ t-cell expansion and thereby turns an acute into a chronic viral infection. PLoS Pathog. 2019, 15, e1007797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdan, T.A.; Lang, P.A.; Lang, K.S. The diverse functions of the ubiquitous fcgamma receptors and their unique constituent, fcrgamma subunit. Pathogens 2020, 9, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort | Age in Years (Median/Range) | Sex (m/f) | Etiology (HBV/HCV/ASH/NASH/Others 1) | Child Score (A/B/C/No Cirrhosis/n.d.) | BCLC Score (0/A/B/C/n.d.) | HCMV Serostatus (pos/neg/n.d.) |
|---|---|---|---|---|---|---|
| HCC | 67/45–84 | 50/10 | 13/13/19/8/7 | 39/10/3/7/1 | 1/23/26/8/2 | 42/18/0 |
| cirrhosis | 61/45–78 | 20/13 | 3/6/12/8/4 | 14/11/8/0/0 | n.a. | 24/8/1 |
| HD | 58/26–86 | 15/21 | n.a. | n.a. | n.a. | 22/14/0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rennert, C.; Tauber, C.; Fehrenbach, P.; Heim, K.; Bettinger, D.; Sogukpinar, Ö.; Schuch, A.; Zecher, B.F.; Bengsch, B.; Lang, S.A.; et al. Adaptive Subsets Limit the Anti-Tumoral NK-Cell Activity in Hepatocellular Carcinoma. Cells 2021, 10, 1369. https://doi.org/10.3390/cells10061369
Rennert C, Tauber C, Fehrenbach P, Heim K, Bettinger D, Sogukpinar Ö, Schuch A, Zecher BF, Bengsch B, Lang SA, et al. Adaptive Subsets Limit the Anti-Tumoral NK-Cell Activity in Hepatocellular Carcinoma. Cells. 2021; 10(6):1369. https://doi.org/10.3390/cells10061369
Chicago/Turabian StyleRennert, Charlotte, Catrin Tauber, Pia Fehrenbach, Kathrin Heim, Dominik Bettinger, Özlem Sogukpinar, Anita Schuch, Britta Franziska Zecher, Bertram Bengsch, Sven A. Lang, and et al. 2021. "Adaptive Subsets Limit the Anti-Tumoral NK-Cell Activity in Hepatocellular Carcinoma" Cells 10, no. 6: 1369. https://doi.org/10.3390/cells10061369
APA StyleRennert, C., Tauber, C., Fehrenbach, P., Heim, K., Bettinger, D., Sogukpinar, Ö., Schuch, A., Zecher, B. F., Bengsch, B., Lang, S. A., Bronsert, P., Björkström, N. K., Fichtner-Feigl, S., Schultheiss, M., Thimme, R., & Hofmann, M. (2021). Adaptive Subsets Limit the Anti-Tumoral NK-Cell Activity in Hepatocellular Carcinoma. Cells, 10(6), 1369. https://doi.org/10.3390/cells10061369