
Viral MicroRNAs Encoded by Nucleocapsid Gene of SARS-CoV-2 Are Detected during Infection, and Targeting Metabolic Pathways in Host Cells

 , and
, and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Infection of SARS-CoV-2
2.2. Small RNA Library Preparation and Sequencing
2.3. Bioinformatics Analysis
2.4. Reverse Transcription (RT) and Droplet-Digital PCR (ddPCR)
2.5. Transfection of Small RNA Mimics
2.6. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.7. Protein Extraction and Western Blotting
2.8. Patient Samples and Ethics Approval
2.9. Statistical Analyses
3. Results and Discussion
3.1. Expression of Viral MicroRNAs Encoded by SARS-CoV-2 N Gene
3.2. V-miRNAs Derived from the N Gene Are Differentially Expressed in COVID-19 Patients
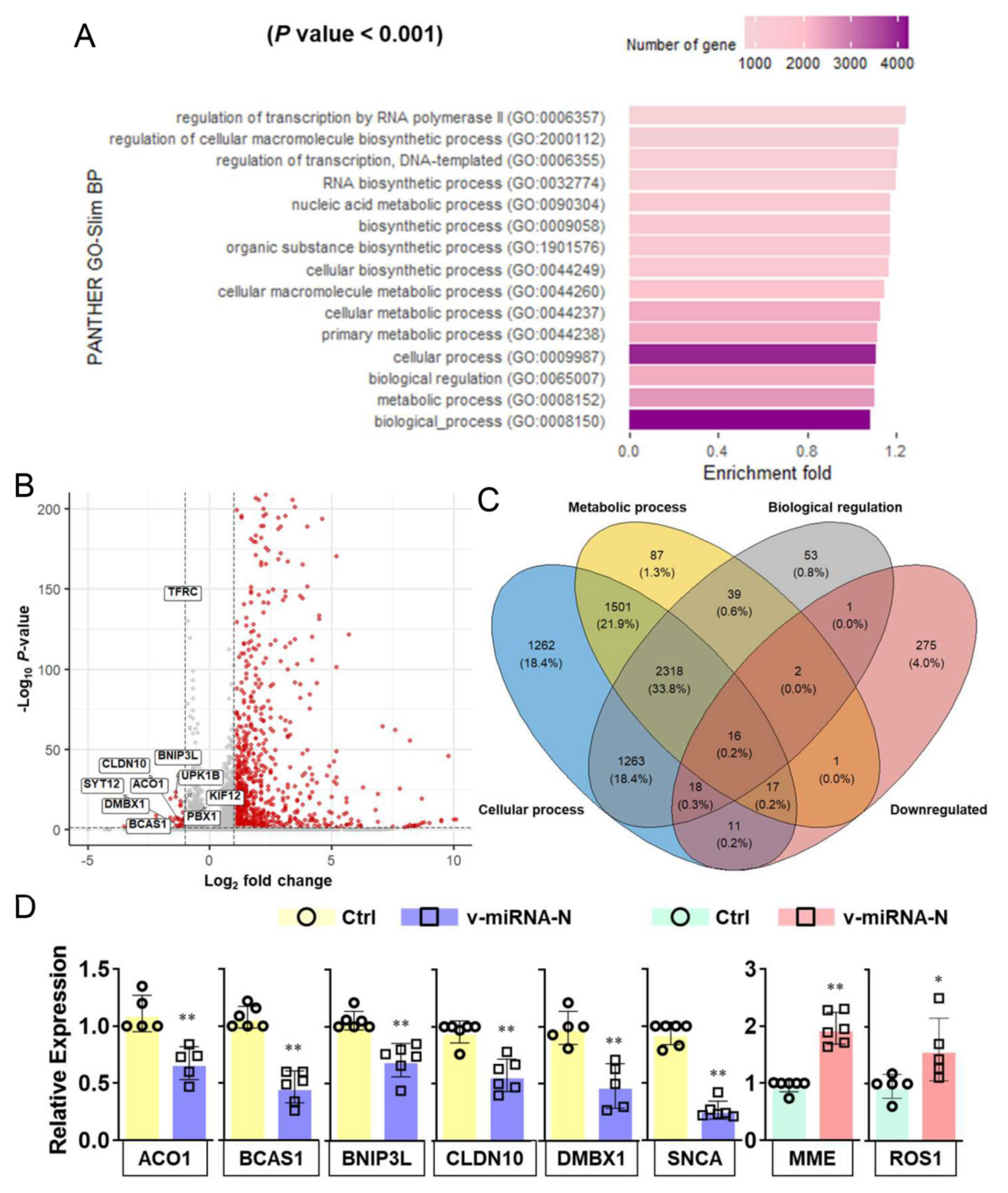
3.3. SARS-CoV-2 V-miRNAs Target Host Genes and Pathways Related to Cellular Metabolic and Biosynthetic Processes
3.4. Increased IL-1β, Caspase 1, and NLRP3 Expressions Were Detected after Transfection of v-miRNA-N Synthetic Mimics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACO1 | Aconitase 1 |
| ATCC | American type culture collection |
| BCAS1 | Brain enriched myelin associated protein 1 |
| BNIP3L | BCL2 interacting protein 3 like |
| CO2 | Carbon dioxide |
| CLDN10 | Claudin 10 |
| COVID-19 | Corona virus disease -19 |
| CXCL10 | C-X-C motif chemokine ligand 10 |
| DEG | Differentially expressed gene |
| DMBX1 | Diencephalon/mesencephalon homeobox 1 |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DNA | Deoxyribonucleic acid |
| FBS | Fetal bovine serum |
| FDR | False discovery rate |
| GO | Gene ontology |
| HK | Hong Kong |
| IL-1β | Interleukin-1 beta |
| IL-6 | Interleukin 6 |
| KIF12 | Kinesin family member 12 |
| LPS | Lipopolysaccharide |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MME | Membrane metalloendopeptidase |
| MOI | Multiplicity of infection |
| NEB | New England biolabs |
| NF-κB | Nuclear factor kappa B |
| NGS | Next-generation sequencing |
| NLRP3 | Nucleotide oligomerization domain-like receptors family pyrin domain containing 3 |
| ORF | Open reading frame |
| PBMC | Peripheral blood mononuclear cell |
| PBX1 | Pre-B-cell leukemia transcription factor 1 |
| PCR | Polymerase chain reaction |
| RNA | Ribonucleic acids |
| ROS 1 | ROS proto-oncogene 1, receptor tyrosine kinase |
| RT | Reverse transcription |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| SD | Standard deviation |
| SNCA | Synuclein alpha |
| SvRNA-N | Small viral RNA nucleocapsid |
| SYT12 | Synaptotagmin 12 |
| TFRC | Transferrin receptor protein 1 |
| USA | United States of America |
| UPK1B | Uroplakin 1B |
| UTR | Untranslated region |
| V-miRNA | Viral microRNAs |
References
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Amahong, K.; Sun, X.; Lian, X.; Liu, J.; Sun, H.; Lou, Y.; Zhu, F.; Qiu, Y. The miRNA: A small but powerful RNA for COVID-19. Brief Bioinform. 2021, 22, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Colpaert, R.M.W.; Calore, M. MicroRNAs in cardiac diseases. Cells 2019, 8, 737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.Z.; Guo, C.; Zou, M.M.; Meng, P.; Zhang, T.T. circRNA-miRNA-mRNA regulatory network in human lung cancer: An update. Cancer Cell Int. 2020, 20, 173. [Google Scholar] [CrossRef] [PubMed]
- Klieser, E.; Mayr, C.; Kiesslich, T.; Wissniowski, T.; Fazio, P.D.; Neureiter, D.; Ocker, M. The crosstalk of miRNA and oxidative stress in the liver: From physiology to pathology and clinical implications. Int. J. Mol. Sci. 2019, 20, 5266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.Q.; Papp, G.; Szodoray, P.; Zeher, M. The role of microRNAs in the pathogenesis of autoimmune diseases. Autoimmun. Rev. 2016, 15, 1171–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier, A.; Sagan, S.M. The diverse roles of microRNAs at the host(-) virus interface. Viruses 2018, 10, 440. [Google Scholar] [CrossRef] [Green Version]
- Kincaid, R.P.; Sullivan, C.S. Virus-encoded microRNAs: An overview and a look to the future. PLoS Pathog. 2012, 8, e1003018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Te Velthuis, A.J.W.; Long, J.C.; Bauer, D.L.V.; Fan, R.L.Y.; Yen, H.L.; Sharps, J.; Siegers, J.Y.; Killip, M.J.; French, H.; Oliva-Martín, M.J.; et al. Mini viral RNAs act as innate immune agonists during influenza virus infection. Nat. Microbiol. Vol. 2018, 3, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Morales, L.; Oliveros, J.C.; Fernandez-Delgado, R.; tenOever, B.R.; Enjuanes, L.; Sola, I. SARS-CoV-encoded small RNAs contribute to infection-associated lung pathology. Cell Host Microbe 2017, 21, 344–355. [Google Scholar] [CrossRef] [Green Version]
- Alkahtani, S.; Alkahtani, M.D.; Alqahtani, A.Q.S.; Alneghery, I.M.; Alqahtani, W.S. A review of comparison study between coronaviruses (SARS-CoV, MERS-CoV) and novel coronavirus (COVID-19). Rev. Mex. de Ing. Química 2020, 19, 201–212. [Google Scholar] [CrossRef]
- Li, J.Y.; You, Z.; Wang, Q.; Zhou, Z.J.; Qiu, Y.; Luo, R.; Ge, X.Y. The epidemic of 2019-novel-coronavirus (2019-nCoV) pneumonia and insights for emerging infectious diseases in the future. Microbes Infect. 2020, 22, 80–85. [Google Scholar] [CrossRef]
- Yoshimoto, F.K. The proteins of severe acute respiratory syndrome coronavirus-2 (SARS CoV-2 or n-COV19), the cause of COVID-19. Protein J. 2020, 39, 198–216. [Google Scholar] [CrossRef] [PubMed]
- Yap, J.K.Y.; Moriyama, M.; Iwasaki, A. Inflammasomes and pyroptosis as therapeutic targets for COVID-19. J. Immunol. 2020, 205, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, Y.; Guan, Z.; Li, H.; Ye, M.; Chen, X.; Shen, J.; Zhou, Y.; Shi, Z.L.; Zhou, P.; et al. SARS-CoV-2 triggers inflammatory responses and cell death through caspase-8 activation. Signal Transduct. Target Ther. 2020, 5, 235. [Google Scholar] [CrossRef]
- Li, C.; Hu, X.; Li, L.; Li, J.H. Differential microRNA expression in the peripheral blood from human patients with COVID-19. J. Clin. Lab. Anal. 2020, 34, e23590. [Google Scholar] [CrossRef] [PubMed]
- Merino, G.A.; Raad, J.; Bugnon, L.A.; Yones, C.; Kamenetzky, L.; Claus, J.; Ariel, F.; Milone, D.H.; Stegmayer, G. Novel SARS-CoV-2 encoded small RNAs in the passage to humans. Bioinformatics 2021, 36, 5571–5581. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, J.; Xu, Y.; Guo, M.; Mi, K.; Xu, R.; Pei, Y.; Zhang, Q.; Luan, X.; Hu, Z.; et al. Implications of the virus-encoded miRNA and host miRNA in the pathogenicity of SARS-CoV-2. arXiv 2020, arXiv:2004.04874. [Google Scholar]
- Gambardella, J.; Sardu, C.; Morelli, M.B.; Messina, V.; Castellanos, V.; Marfella, R.; Maggi, P.; Paolisso, G.; Wang, X.; Santulli, G. Exosomal microRNAs drive thrombosis in COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, R.; Kim, S.H.; Shah, H.; Zhang, S.; Liang, J.H.; Fang, Y.; Gentili, M.; Leary, C.N.O.; Elledge, S.J.; et al. SARS-CoV-2 hijacks folate and one-carbon metabolism for viral replication. Nat. Commun. 2021, 12, 1676. [Google Scholar] [CrossRef]
- Chen, J.; Wang, R.; Wang, M.; Wei, G.W. Mutations strengthened SARS-CoV-2 infectivity. J. Mol. Biol. 2020, 432, 5212–5226. [Google Scholar] [CrossRef]
- Mok, B.W.-Y.; Cremin, C.J.; Lau, S.-Y.; Deng, S.; Chen, P.; Zhang, A.J.; Lee, A.C.-Y.; Liu, H.; Liu, S.; Ng, T.T.-L.; et al. SARS-CoV-2 spike D614G variant exhibits highly efficient replication and transmission in hamsters. bioRxiv 2020. [Google Scholar] [CrossRef]
- Mok, B.W.-Y.; Liu, H.; Lau, S.-Y.; Deng, S.; Liu, S.; Tam, R.C.-Y.; Ng, T.T.-L.; Leung, J.S.-L.; Wang, P.; To, K.K.-W.; et al. Low dose inocula of SARS-CoV-2 B.1.1.7 variant initiate more robust infections in the upper respiratory tract of hamsters than earlier D614G variants. bioRxiv 2021. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, R.; Hofacker, I.L.; Stadler, P.F. RNA folding with hard and soft constraints. Algorithms Mol. Biol. 2016, 11, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerpedjiev, P.; Hammer, S.; Hofacker, I.L. Forna (force-directed RNA): Simple and effective online RNA secondary structure diagrams. Bioinformatics 2015, 31, 3377–3379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [Green Version]
- Reczko, M.; Maragkakis, M.; Alexiou, P.; Grosse, I.; Hatzigeorgiou, A.G. Functional microRNA targets in protein coding sequences. Bioinformatics 2012, 28, 771–776. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gkirtzou, K.; Tsamardinos, I.; Tsakalides, P.; Poirazi, P. MatureBayes: A probabilistic algorithm for identifying the mature miRNA within novel precursors. PLoS ONE 2010, 5, e11843. [Google Scholar] [CrossRef] [PubMed]
- Smits, V.A.J.; Hernandez-Carralero, E.; Paz-Cabrera, M.C.; Cabrera, E.; Hernandez-Reyes, Y.; Hernandez-Fernaud, J.R.; Gillespie, D.A.; Salido, E.; Hernandez-Porto, M.; Freire, R. The nucleocapsid protein triggers the main humoral immune response in COVID-19 patients. Biochem. Biophys. Res. Commun. 2021, 543, 45–49. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, Q.; Chen, J.; Ni, X.; Dai, J. A DNA aptamer based method for detection of SARS-CoV-2 nucleocapsid protein. Virol. Sin. 2020, 35, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; McGrath, J.; Connolly, S.P.; Lambert, J. Partial N gene sequencing for SARS-CoV-2 verification and pathway tracing. Int. Med. Case Rep. J. 2021, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fajnzylber, J.; Regan, J.; Coxen, K.; Corry, H.; Wong, C.; Rosenthal, A.; Worrall, D.; Giguel, F.; Piechocka-Trocha, A.; Atyeo, C.; et al. SARS-CoV-2 viral load is associated with increased disease severity and mortality. Nat. Commun. 2020, 11, 5493. [Google Scholar] [CrossRef] [PubMed]
- Argyropoulos, K.V.; Serrano, A.; Hu, J.; Black, M.; Feng, X.; Shen, G.; Call, M.; Kim, M.J.; Lytle, A.; Belovarac, B.; et al. Association of initial viral load in severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) patients with wutcome and symptoms. Am. J. Pathol. 2020, 190, 1881–1887. [Google Scholar] [CrossRef] [PubMed]
- Hopp, M.-T.; Domingo-Fernández, D.; Gadiya, Y.; Detzel, M.S.; Schmalohr, B.F.; Steinbock, F.; Imhof, D.; Hofmann-Apitius, M. Unravelling the debate on heme effects in COVID-19 infections. bioRxiv 2020. [Google Scholar] [CrossRef]
- Thair, S.A.; He, Y.D.; Hasin-Brumshtein, Y.; Sakaram, S.; Pandya, R.; Toh, J.; Rawling, D.; Remmel, M.; Coyle, S.; Dalekos, G.N.; et al. Transcriptomic similarities and differences in host response between SARS-CoV-2 and other viral infections. iScience 2021, 24, 101947. [Google Scholar] [CrossRef]
- Fard, M.K.; Van der Meer, F.; Sánchez, P.; Cantuti-Castelvetri, L.; Mandad, S.; Jäkel, S.; Fornasiero, E.F.; Schmitt, S.; Ehrlich, M.; Starost, L.; et al. BCAS1 expression defines a population of early myelinating oligodendrocytes in multiple sclerosis lesions. Sci. Transl. Med. 2017, 9. [CrossRef] [Green Version]
- Hadj-Rabia, S.; Brideau, G.; Al-Sarraj, Y.; Maroun, R.C.; Figueres, M.L.; Leclerc-Mercier, S.; Olinger, E.; Baron, S.; Chaussain, C.; Nochy, D.; et al. Multiplex epithelium dysfunction due to CLDN10 mutation: The HELIX syndrome. Genet. Med. 2018, 20, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, S.; Nordqvist, H.; Ambikan, A.T.; Gupta, S.; Sperk, M.; Svensson-Akusjärvi, S.; Mikaeloff, F.; Benfeitas, R.; Saccon, E.; Ponnan, S.M.; et al. Implications of central carbon metabolism in SARS-CoV-2 replication and disease severity. bioRxiv 2021. [Google Scholar] [CrossRef]
- Imig, J.; Brunschweiger, A.; Brummer, A.; Guennewig, B.; Mittal, N.; Kishore, S.; Tsikrika, P.; Gerber, A.P.; Zavolan, M.; Hall, J. miR-CLIP capture of a miRNA targetome uncovers a lincRNA H19-miR-106a interaction. Nat. Chem. Biol. 2015, 11, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, M.N.; Aydemir, H.B.; Korkmaz, E.M.; Budak, M.; Cekin, N.; Pinarbasi, E. Computationally predicted SARS-COV-2 encoded microRNAs target NFKB, JAK/STAT and TGFB signaling pathways. Gene Rep. 2021, 22, 101012. [Google Scholar] [CrossRef]
- Chen, I.Y.; Ichinohe, T. Response of host inflammasomes to viral infection. Trends Microbiol. 2015, 23, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Bryant, C.; Fitzgerald, K.A. Molecular mechanisms involved in inflammasome activation. Trends Cell Biol. 2009, 19, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Lupfer, C.; Kanneganti, T.D. The expanding role of NLRs in antiviral immunity. Immunol. Rev. 2013, 255, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, G.; Leon-Juarez, M.; Garcia-Cordero, J.; Meza-Sanchez, D.E.; Cedillo-Barron, L. Inflammasomes and its importance in viral infections. Immunol. Res. 2016, 64, 1101–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, R.J.; Cao, W.J.; Li, M.H.; Yang, L.; Dai, R.X.; Luo, X.L.; Liu, Y.; Ge, B.X.; Su, X.; Xu, J.F. PM2.5 compromises antiviral immunity in influenza infection by inhibiting activation of NLRP3 inflammasome and expression of interferon-beta. Mol. Immunol. 2020, 125, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Callaway, J.B.; Ting, J.P. Inflammasomes: Mechanism of action, role in disease, and therapeutics. Nat. Med. 2015, 21, 677–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haneklaus, M.; Gerlic, M.; Kurowska-Stolarska, M.; Rainey, A.A.; Pich, D.; McInnes, I.B.; Hammerschmidt, W.; O’Neill, L.A.; Masters, S.L. Cutting edge: miR-223 and EBV miR-BART15 regulate the NLRP3 inflammasome and IL-1beta production. J. Immunol. 2012, 189, 3795–3799. [Google Scholar] [CrossRef] [Green Version]
- McElvaney, O.J.; McEvoy, N.L.; McElvaney, O.F.; Carroll, T.P.; Murphy, M.P.; Dunlea, D.M.; Ni Choileain, O.; Clarke, J.; O’Connor, E.; Hogan, G.; et al. Characterization of the inflammatory response to severe COVID-19 illness. Am. J. Respir. Crit. Care Med. 2020, 202, 812–821. [Google Scholar] [CrossRef]
- Costa, F.F.; Rosario, W.R.; Ribeiro Farias, A.C.; de Souza, R.G.; Duarte Gondim, R.S.; Barroso, W.A. Metabolic syndrome and COVID-19: An update on the associated comorbidities and proposed therapies. Diabetes Metab. Syndr. 2020, 14, 809–814. [Google Scholar] [CrossRef]
- Andrade Silva, M.; da Silva, A.; do Amaral, M.A.; Fragas, M.G.; Camara, N.O.S. Metabolic alterations in SARS-CoV-2 infection and its implication in kidney dysfunction. Front. Physiol. 2021, 12, 624698. [Google Scholar] [CrossRef]
- Nigro, E.; Perrotta, F.; Polito, R.; D’Agnano, V.; Scialo, F.; Bianco, A.; Daniele, A. Metabolic perturbations and severe COVID-19 disease: Implication of molecular pathways. Int. J. Endocrinol. 2020, 2020, 8896536. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, A.; Uhl, S.; Ioannidis, K.; Hofree, M.; tenOever, B.R.; Nahmias, Y. The SARS-CoV-2 transcriptional metabolic signature in lung epithelium. SSRN 2020. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=3650499 (accessed on 27 May 2021).
- Habib, H.M.; Ibrahim, S.; Zaim, A.; Ibrahim, W.H. The role of iron in the pathogenesis of COVID-19 and possible treatment with lactoferrin and other iron chelators. Biomed. Pharmacother. 2021, 136, 111228. [Google Scholar] [CrossRef]
- Moreira, A.C.; Mesquita, G.; Gomes, M.S. Ferritin: An inflammatory player keeping iron at the core of pathogen-host interactions. Microorganisms 2020, 8, 589. [Google Scholar] [CrossRef] [Green Version]
- Hippchen, T.; Altamura, S.; Muckenthaler, M.U.; Merle, U. Hypoferremia is associated with increased hospitalization and oxygen demand in COVID-19 patients. Hemasphere 2020, 4, e492. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Huang, J.; Dai, D.; Feng, Y.; Liu, L.; Nie, S. Serum iron level as a potential predictor of coronavirus disease 2019 severity and mortality: A retrospective study. Open Forum Infect. Dis. 2020, 7, ofaa250. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, F.; Siu, G.K.-H.; Mok, B.W.-Y.; Sun, J.; Fung, K.S.C.; Lam, J.Y.-W.; Wong, N.K.; Gedefaw, L.; Luo, S.; Lee, T.M.H.; et al. Viral MicroRNAs Encoded by Nucleocapsid Gene of SARS-CoV-2 Are Detected during Infection, and Targeting Metabolic Pathways in Host Cells. Cells 2021, 10, 1762. https://doi.org/10.3390/cells10071762
Meng F, Siu GK-H, Mok BW-Y, Sun J, Fung KSC, Lam JY-W, Wong NK, Gedefaw L, Luo S, Lee TMH, et al. Viral MicroRNAs Encoded by Nucleocapsid Gene of SARS-CoV-2 Are Detected during Infection, and Targeting Metabolic Pathways in Host Cells. Cells. 2021; 10(7):1762. https://doi.org/10.3390/cells10071762
Chicago/Turabian StyleMeng, Fei, Gilman Kit-Hang Siu, Bobo Wing-Yee Mok, Jiahong Sun, Kitty S. C. Fung, Jimmy Yiu-Wing Lam, Nonthaphat Kent Wong, Lealem Gedefaw, Shumeng Luo, Thomas M. H. Lee, and et al. 2021. "Viral MicroRNAs Encoded by Nucleocapsid Gene of SARS-CoV-2 Are Detected during Infection, and Targeting Metabolic Pathways in Host Cells" Cells 10, no. 7: 1762. https://doi.org/10.3390/cells10071762
APA StyleMeng, F., Siu, G. K.-H., Mok, B. W.-Y., Sun, J., Fung, K. S. C., Lam, J. Y.-W., Wong, N. K., Gedefaw, L., Luo, S., Lee, T. M. H., Yip, S. P., & Huang, C.-L. (2021). Viral MicroRNAs Encoded by Nucleocapsid Gene of SARS-CoV-2 Are Detected during Infection, and Targeting Metabolic Pathways in Host Cells. Cells, 10(7), 1762. https://doi.org/10.3390/cells10071762