
Serotonin Heteroreceptor Complexes and Their Integration of Signals in Neurons and Astroglia—Relevance for Mental Diseases

 , , ,
, , ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. D2R-5-HT2AR and D2R-5-HT1AR Heterocomplexes and Mental Disease
2.1. D2R-5-HT2AR Heterocomplex
2.2. D2R-5-HT1AR Heterocomplex
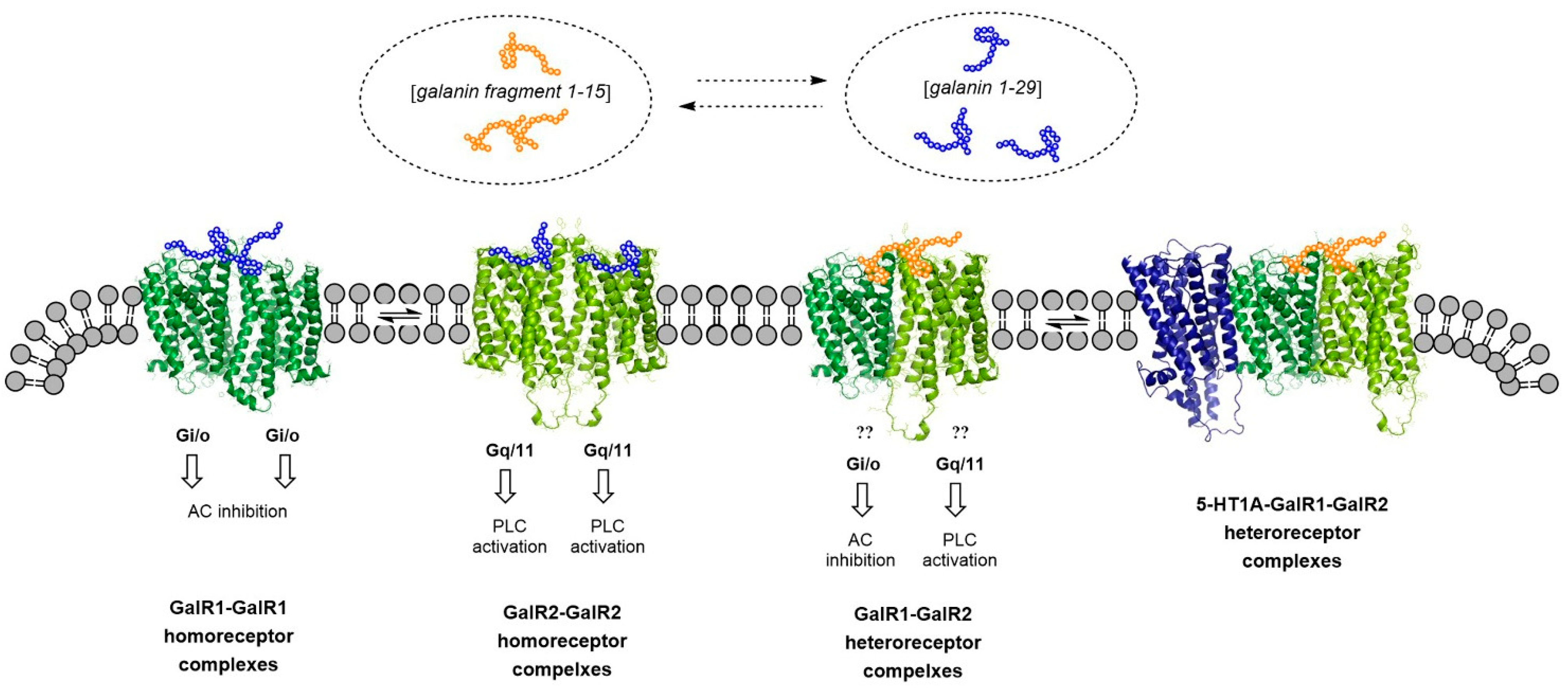
3. The GalR1-GalR2 Heterodimer and the Putative GalR1-GalR2-5-HT1A Heteroreceptor Complexes. Targets for Galanin N-terminal Fragment Gal (1–15), a Modulator of Emotional Networks in Models of Mental Disease
Gal (1–15) and Its Modulation of the Meso-Limbic DA Reward Neurons Causes Anhedonia
4. Serotonin 5-HT1AR-Receptor Tyrosine Kinase (RTK) Heteroreceptor Complexes
5. Other Types of GPCR-RTK Heterocomplexes and Treatment of Depression
5.1. GPCR-TrkB Heterocomplexes
5.2. Muscarinic Acetylcholine Receptor (mAChR)-FGFR1 Hetero Complexes
5.3. GPCR-Epidermal Growth Factor Receptor (EGFR) Heterocomplexes
5.4. Do GPCR-Platelet-Derived Growth Factor Receptor (PDGFR) Heteroreceptor Complexes Exist?
6. Can the Biological Principle of Forming Heteroreceptor Complexes in the Brain Help Understand the Mechanism of Action of Hallucinogens?
6.1. MDMA (3,4-Methylenedioxymethamphetamine)
6.2. Psilocybin
6.3. Ketamine
7. Understanding the Role of Astrocytes in Modulating Forebrain Glutamate Synapses with Impact on Dorsal Raphe-Forebrain Serotonin Neurons Playing a Key Role in Major Depressive Disorder
7.1. Key Features of Astrocytes
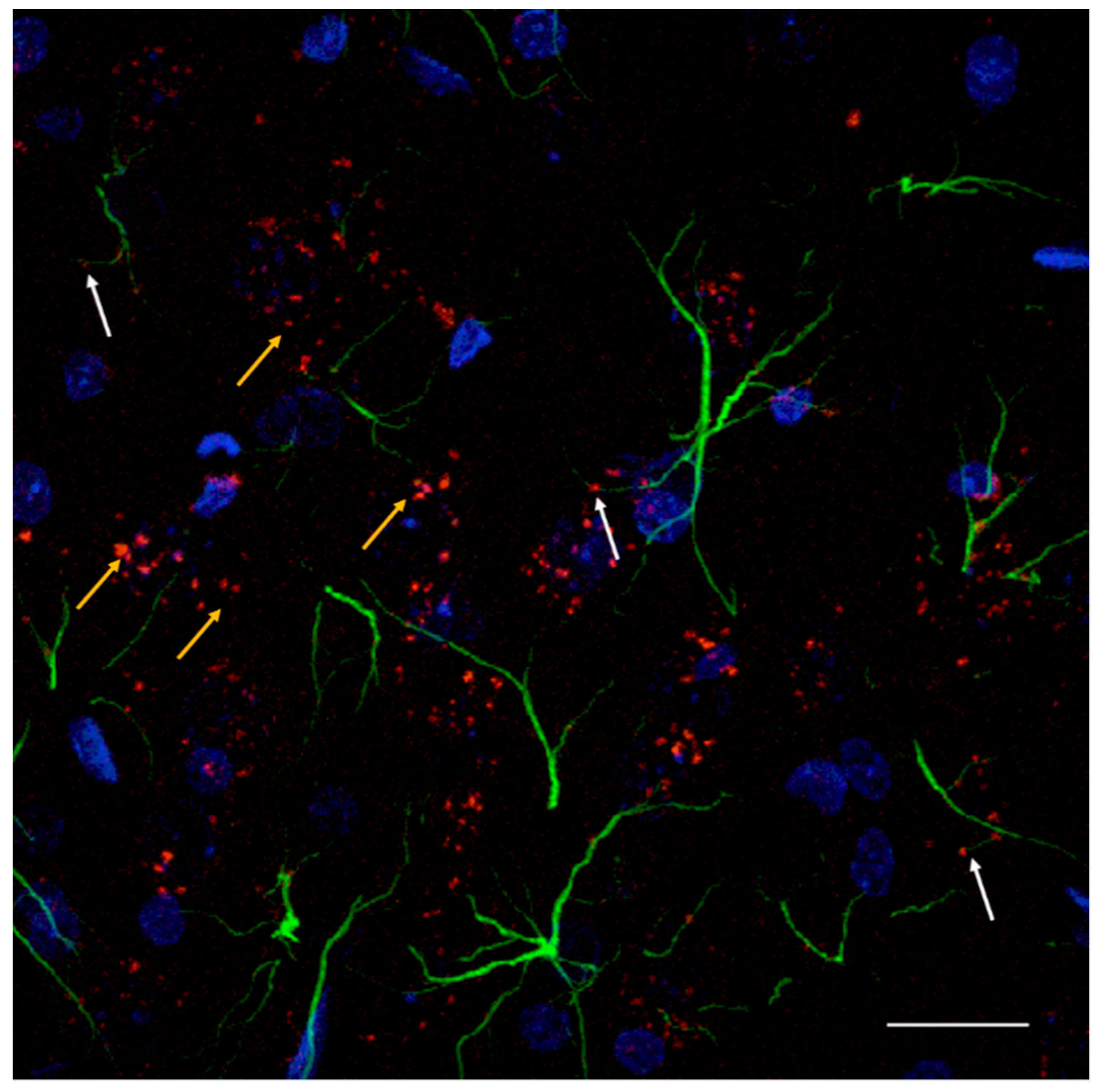
7.2. Astroglial Heteroreceptor Complexes
7.3. On the Role of Astrocytes in the Control of Glutamatergic Activity in the Infralimbic Cortex Modulating the Activity of the Ascending Raphe-Limbic-Cortical Systems
7.4. Selective Knockdown of Astrocytic Glutamate Transporters in the Infralimbic Cortex. Depressive-Like Phenotypes in Mice
8. Conclusions and Future Work
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Borroto-Escuela, D.O.; Fuxe, K. Oligomeric Receptor Complexes and Their Allosteric Receptor-Receptor Interactions in the Plasma Membrane Represent a New Biological Principle for Integration of Signals in the CNS. Front. Mol. Neurosci. 2019, 12, 230. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Narvaez, M.; Ambrogini, P.; Ferraro, L.; Brito, I.; Romero-Fernandez, W.; Andrade-Talavera, Y.; Flores-Burgess, A.; Millón, C.; Gago, B.; et al. Receptor–Receptor Interactions in Multiple 5-HT1A Heteroreceptor Complexes in Raphe-Hippocampal 5-HT Transmission and Their Relevance for Depression and Its Treatment. Molecules 2018, 23, 1341. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.; Ambrogini, P.; Chruścicka, B.; Lindskog, M.; Crespo-Ramirez, M.; Hernández-Mondragón, J.; de la Mora, M.P.; Schellekens, H.; Fuxe, K. The Role of Central Serotonin Neurons and 5-HT Heteroreceptor Complexes in the Pathophysiology of Depression: A Historical Perspective and Future Prospects. Int. J. Mol. Sci. 2021, 22, 1927. [Google Scholar] [CrossRef] [PubMed]
- Artigas, F. Serotonin receptors involved in antidepressant effects. Pharmacol. Ther. 2013, 137, 119–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artigas, F. Developments in the field of antidepressants, where do we go now? Eur. Neuropsychopharmacol. 2015, 25, 657–670. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Tarakanov, A.O.; Fuxe, K. FGFR1–5-HT1A Heteroreceptor Complexes: Implications for Understanding and Treating Major Depression. Trends Neurosci. 2016, 39, 5–15. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Dupont, C.M.; Li, X.; Savelli, D.; Lattanzi, D.; Srivastava, I.; Narváez, M.; Di Palma, M.; Barbieri, E.; Andrade-Talavera, Y.; et al. Disturbances in the FGFR1-5-HT1A Heteroreceptor Complexes in the Raphe-Hippocampal 5-HT System Develop in a Genetic Rat Model of Depression. Front. Cell. Neurosci. 2017, 11, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millón, C.; Flores-Burgess, A.; Narváez, M.; Borroto-Escuela, D.O.; Santin, L.J.; Gago, B.; Narvaez, J.A.; Fuxe, K.; Díaz-Cabiale, Z.; Cabiale, M.Z.D. Galanin (1–15) enhances the antidepressant effects of the 5-HT1A receptor agonist 8-OH-DPAT: Involvement of the raphe-hippocampal 5-HT neuron system. Brain Struct. Funct. 2016, 221, 4491–4504. [Google Scholar] [CrossRef]
- Millón, C.; Flores, A.; Narváez, M.; Escuela, D.O.B.; Gago, B.; Santín, L.; Castilla-Ortega, E.; Narvaez, J.A.; Fuxe, K.; Díaz-Cabiale, Z. The neuropeptides Galanin and Galanin(1–15) in depression-like behaviours. Neuropeptides 2017, 64, 39–45. [Google Scholar] [CrossRef]
- Fuxe, K.; Borroto-Escuela, D.O.; Romero-Fernandez, W.; Tarakanov, A.O.; Calvo, F.; Garriga, P.; Tena, M.; Narvaez, M.; Millón, C.; Parrado, C.; et al. On the existence and function of galanin receptor heteromers in the central nervous system. Front. Endocrinol. 2012, 3, 127. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Tarakanov, A.O.; Marcellino, D.; Ciruela, F.; Agnati, L.F.; Fuxe, K. Dopamine D2 and 5-hydroxytryptamine 5-HT2A receptors assemble into functionally interacting heteromers. Biochem. Biophys. Res. Commun. 2010, 401, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Łukasiewicz, S.; Polit, A.; Kedracka-Krok, S.; Wedzony, K.; Maćkowiak, M.; Dziedzicka-Wasylewska, M. Hetero-dimerization of serotonin 5-HT2A and dopamine D2 receptors. Biochim. Biophys. Acta (BBA)—Bioenerg. 2010, 1803, 1347–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fullana, N.; Gasull-Camós, J.; Gatius, M.T.; Castañé, A.; Bortolozzi, A.; Artigas, F. Astrocyte control of glutamatergic activity: Downstream effects on serotonergic function and emotional behavior. Neuropharmacology 2020, 166, 107914. [Google Scholar] [CrossRef] [PubMed]
- Cervetto, C.; Venturini, A.; Passalacqua, M.; Guidolin, D.; Genedani, S.; Fuxe, K.; Borroto-Esquela, D.O.; Cortelli, P.; Woods, A.; Maura, G.; et al. A2A-D2 receptor-receptor interaction modulates gliotransmitter release from striatal astrocyte processes. J. Neurochem. 2016, 140, 268–279. [Google Scholar] [CrossRef]
- Narváez, M.; Andrade-Talavera, Y.; Valladolid-Acebes, I.; Fredriksson, M.; Siegele, P.; Hernandez-Sosa, A.; Fisahn, A.; Fuxe, K.; Borroto-Escuela, D.O. Existence of FGFR1-5-HT1AR heteroreceptor complexes in hippocampal astrocytes. Putative link to 5-HT and FGF2 modulation of hippocampal gamma oscillations. Neuropharmacology 2020, 170, 108070. [Google Scholar] [CrossRef]
- Matsushita, H.; Latt, H.M.; Koga, Y.; Nishiki, T.; Matsui, H. Oxytocin and Stress: Neural Mechanisms, Stress-Related Disorders, and Therapeutic Approaches. Neuroscience 2019, 417, 1–10. [Google Scholar] [CrossRef]
- Ciruela, F.; Burgueño, J.; Casadó, V.; Canals, M.; Marcellino, D.; Goldberg, S.R.; Bader, M.; Fuxe, K.; Agnati, L.F.; Lluis, C.; et al. Combining Mass Spectrometry and Pull-Down Techniques for the Study of Receptor Heteromerization. Direct Epitope−Epitope Electrostatic Interactions between Adenosine A2Aand Dopamine D2Receptors. Anal. Chem. 2004, 76, 5354–5363. [Google Scholar] [CrossRef]
- Andén, N.-E.; Corrodi, H.; Fuxe, K.; Hökfelt, T. Evidence for a central 5-hydroxytryptamine receptor stimulation by lysergic acid diethylamide. Br. J. Pharmacol. 1968, 34, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Agnati, L.F.; Ungerstedt, U. The effect of mepiprazole on central monoamine neurons. Evidence for increased 5-hydroxytryptamine and dopamine receptor activity. Eur. J. Pharmacol. 1976, 35, 93–107. [Google Scholar] [CrossRef]
- Niimi, K.; Takahashi, E.; Itakura, C. Hallucinogenic 5-Hydroxytryptamine 2A Receptor Agonist Effects in Senescence-Accelerated Mice. Exp. Anim. 2010, 59, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Narvaez, M.; Oflijan, J.; Agnati, L.F.; Fuxe, K. Hallucinogenic 5-HT2AR agonists LSD and DOI enhance dopamine D2R protomer recognition and signaling of D2-5-HT2A heteroreceptor complexes. Biochem. Biophys. Res. Commun. 2014, 443, 278–284. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Pintsuk, J.; Schäfer, T.; Friedland, K.; Ferraro, L.; Tanganelli, S.; Liu, F.; Fuxe, K. Multiple D2 heteroreceptor complexes: New targets for treatment of schizophrenia. Ther. Adv. Psychopharmacol. 2016, 6, 77–94. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, S.; Duncan, G.E.; Marx, C.E.; Lieberman, J.A. Treatments for schizophrenia: A critical review of pharmacology and mechanisms of action of antipsychotic drugs. Mol. Psychiatry 2004, 10, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Meltzer, H.Y.; Massey, B.W.; Horiguchi, M. Serotonin receptors as targets for drugs useful to treat psychosis and cognitive impairment in schizophrenia. Curr. Pharm. Biotechnol. 2012, 13, 1572–1586. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Fuxe, K. Diversity and bias through dopamine D2R heteroreceptor complexes. Curr. Opin. Pharmacol. 2017, 32, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Ferraro, L.; Narvaez, M.; Tanganelli, S.; Beggiato, S.; Liu, F.; Rivera, A.; Fuxe, K. Multiple Adenosine-Dopamine (A2A-D2 Like) Heteroreceptor Complexes in the Brain and Their Role in Schizophrenia. Cells 2020, 9, 1077. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, F.X.; Csomor, P.A.; Knappe, B.; Geyer, M.A.; Quednow, B.B. The Effects of the Preferential 5-HT2A Agonist Psilocybin on Prepulse Inhibition of Startle in Healthy Human Volunteers Depend on Interstimulus Interval. Neuropsychopharmacology 2007, 32, 1876–1887. [Google Scholar] [CrossRef] [Green Version]
- Bankson, M.G.; Cunningham, K.A. 3,4-Methylenedioxymethamphetamine (MDMA) as a unique model of serotonin receptor function and serotonin-dopamine interactions. J. Pharmacol. Exp. Ther. 2001, 297, 846–852. [Google Scholar] [PubMed]
- Mithoefer, M.C.; Wagner, M.T.; Mithoefer, A.T.; Jerome, L.; Doblin, R. The safety and efficacy of ±3,4-methylenedioxymethamphetamine-assisted psychotherapy in subjects with chronic, treatment-resistant posttraumatic stress disorder: The first randomized controlled pilot study. J. Psychopharmacol. 2010, 25, 439–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chruścicka, B.; Fitzsimons, S.E.W.; Borroto-Escuela, D.O.; Druelle, C.; Stamou, P.; Nally, K.; Dinan, T.G.; Cryan, J.F.; Fuxe, K.; Schellekens, H. Attenuation of Oxytocin and Serotonin 2A Receptor Signaling through Novel Heteroreceptor Formation. ACS Chem. Neurosci. 2019, 10, 3225–3240. [Google Scholar] [CrossRef] [PubMed]
- Jurek, B.; Neumann, I.D. The Oxytocin Receptor: From Intracellular Signaling to Behavior. Physiol. Rev. 2018, 98, 1805–1908. [Google Scholar] [CrossRef]
- Łukasiewicz, S.; Błasiak, E.; Szafran-Pilch, K.; Dziedzicka-Wasylewska, M. Dopamine D2 and serotonin 5-HT1A receptor interaction in the context of the effects of antipsychotics—in vitro studies. J. Neurochem. 2016, 137, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Kolasa, M.; Solich, J.; Faron-Górecka, A.; Żurawek, D.; Pabian, P.; Łukasiewicz, S.; Kuśmider, M.; Szafran-Pilch, K.; Szlachta, M.; Dziedzicka-Wasylewska, M. Paroxetine and Low-dose Risperidone Induce Serotonin 5-HT1A and Dopamine D2 Receptor Heteromerization in the Mouse Prefrontal Cortex. Neuroscience 2018, 377, 184–196. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Li, X.; Tarakanov, A.O.; Savelli, D.; Narváez, M.; Shumilov, K.; Andrade-Talavera, Y.; Jimenez-Beristain, A.; Pomierny, B.; Díaz-Cabiale, Z.; et al. Existence of Brain 5-HT1A–5-HT2A Isoreceptor Complexes with Antagonistic Allosteric Receptor–Receptor Interactions Regulating 5-HT1A Receptor Recognition. ACS Omega 2017, 2, 4779–4789. [Google Scholar] [CrossRef] [PubMed]
- Millón, C.; Flores-Burgess, A.; Narváez, M.; Borroto-Escuela, D.O.; Santin, L.J.; Parrado, C.; Narvaez, J.A.; Fuxe, K.; Díaz-Cabiale, Z. A Role for Galanin N-Terminal Fragment (1–15) in Anxiety- and Depression-Related Behaviors in Rats. Int. J. Neuropsychopharmacol. 2015, 18, pyu064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedlund, P.B.; Yanaihara, N.; Fuxe, K. Evidence for specific N-terminal galanin fragment binding sites in the rat brain. Eur. J. Pharmacol. 1992, 224, 203–205. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Narvaez, M.; Di Palma, M.; Calvo, F.; Rodriguez, D.; Millón, C.; Carlsson, J.; Agnati, L.F.; Garriga, P.; Cabiale, M.Z.D.; et al. Preferential activation by galanin 1–15 fragment of the GalR1 protomer of a GalR1–GalR2 heteroreceptor complex. Biochem. Biophys. Res. Commun. 2014, 452, 347–353. [Google Scholar] [CrossRef]
- Diaz-Cabiale, Z.; Narvaez, J.A.; Finnman, U.B.; Bellido, I.; Ogren, S.O.; Fuxe, K. Galanin-(1-16) modulates 5-HT1A receptors in the ventral limbic cortex of the rat. Neuroreport 2000, 11, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Cabiale, Z.; Parrado, C.; Narváez, M.; Millón, C.; Puigcerver, A.; Fuxe, K.; Narvaez, J.A. The Galanin N-terminal fragment (1–15) interacts with neuropeptide Y in central cardiovascular control: Involvement of the NPY Y2 receptor subtype. Regul. Pept. 2010, 163, 130–136. [Google Scholar] [CrossRef]
- Hedlund, P.B.; Finnman, U.-B.; Yanaihara, N.; Fuxe, K. Galanin-(1–15), but not galanin-(1–29), modulates 5-HT1A receptors in the dorsal hippocampus of the rat brain: Possible existence of galanin receptor subtypes. Brain Res. 1994, 634, 163–167. [Google Scholar] [CrossRef]
- Flores-Burgess, A.; Millón, C.; Gago, B.; Narváez, M.; Borroto-Escuela, D.O.; Mengod, G.; Narvaez, J.A.; Fuxe, K.; Santín, L.; Díaz-Cabiale, Z.; et al. Galanin (1-15) enhancement of the behavioral effects of Fluoxetine in the forced swimming test gives a new therapeutic strategy against depression. Neuropharmacology 2017, 118, 233–241. [Google Scholar] [CrossRef]
- Flores-Burgess, A.; Millon, C.; Gago, B.; Narvaez, J.A.; Fuxe, K.; Diaz-Cabiale, Z. Small interference RNA knockdown rats in behavioral functions. GalR1-GalR2 heteroreceptor in anxiety and depression-like behaviour. In Receptor-Receptor Interactions in the Central Nervous System; Fuxe, K., Borroto-Escuela, D.O., Eds.; Humana Press: New York, NY, USA, 2018; Volume 140, pp. 133–149. [Google Scholar]
- Flores-Burgess, A.; Millón, C.; Gago, B.; García-Durán, L.; Cantero-García, N.; Coveñas, R.; Narváez, J.A.; Fuxe, K.; Santín, L.; Díaz-Cabiale, Z. Galanin (1–15)-fluoxetine interaction in the novel object recognition test. Involvement of 5-HT1A receptors in the prefrontal cortex of the rats. Neuropharmacology 2019, 155, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Carlsson, J.; Ambrogini, P.; Narváez, M.; Wydra, K.; Tarakanov, A.O.; Li, X.; Millón, C.; Ferraro, L.; Cuppini, R.; et al. Understanding the Role of GPCR Heteroreceptor Complexes in Modulating the Brain Networks in Health and Disease. Front. Cell. Neurosci. 2017, 11, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuxe, K.; Agnati, L.F.; Borroto-Escuela, D.O. The impact of receptor–receptor interactions in heteroreceptor complexes on brain plasticity. Expert Rev. Neurother. 2014, 14, 719–721. [Google Scholar] [CrossRef] [Green Version]
- Millón, C.; Flores, A.; Gago, B.; Alén, F.; Orio, L.; García-Durán, L.; Narváez, J.A.; Fuxe, K.; Santín, L.; Díaz-Cabiale, Z. Role of the galanin N-terminal fragment (1-15) in anhedonia: Involvement of the dopaminergic mesolimbic system. J. Psychopharmacol. 2019, 33, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.; Shen, W.; Graves, S.M.; Surmeier, D.J. Dopaminergic modulation of striatal function and Parkinson’s disease. J. Neural Transm. 2019, 126, 411–422. [Google Scholar] [CrossRef]
- Fuxe, K.; Canals, M.; Torvinen, M.; Marcellino, D.; Terasmaa, A.; Genedani, S.; Leo, G.; Guidolin, D.; Cabiale, M.Z.D.; Rivera, A.; et al. Intramembrane receptor–receptor interactions: A novel principle in molecular medicine. J. Neural Transm. 2006, 114, 49–75. [Google Scholar] [CrossRef] [PubMed]
- Di Liberto, V.; Mudò, G.; Belluardo, N. Crosstalk between receptor tyrosine kinases (RTKs) and G protein-coupled receptors (GPCR) in the brain: Focus on heteroreceptor complexes and related functional neurotrophic effects. Neuropharmacology 2019, 152, 67–77. [Google Scholar] [CrossRef]
- Kitayama, I.; Cintra, A.; Janson, A.M.; Fuxe, K.; Agnati, L.F.; Eneroth, P.; Aronsson, M.; Härfstrand, A.; Steinbush, H.W.M.; Visser, T.J.; et al. Chronic immobilization stress: Evidence for decreases of 5-hydroxy-tryptamine immunoreactivity and for increases of glucocorticoid receptor immunoreactivity in various brain regions of the male rat. J. Neural Transm. 1989, 77, 93–130. [Google Scholar] [CrossRef]
- Di Palma, M.; Sartini, S.; Lattanzi, D.; Cuppini, R.; Pita-Rodriguez, M.; Diaz-Carmenate, Y.; Narvaez, M.; Fuxe, K.; Borroto-Escuela, D.O.; Ambrogini, P. Evidence for the existence of A2AR-TrkB heteroreceptor complexes in the dorsal hippocampus of the rat brain: Potential implications of A2AR and TrkB interplay upon ageing. Mech. Ageing Dev. 2020, 190, 111289. [Google Scholar] [CrossRef]
- Flajolet, M.; Wang, Z.; Futter, M.; Shen, W.; Nuangchamnong, N.; Bendor, J.; Wallach, I.; Nairn, A.C.; Surmeier, D.J.; Greengard, P.; et al. FGF acts as a co-transmitter through adenosine A2A receptor to regulate synaptic plasticity. Nat. Neurosci. 2008, 11, 1402–1409. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Mudo’, G.; Pérez-Alea, M.; Ciruela, F.; Tarakanov, A.O.; Narvaez, M.; Di Liberto, V.; Agnati, L.F.; Belluardo, N.; et al. Fibroblast Growth Factor Receptor 1– 5-Hydroxytryptamine 1A Heteroreceptor Complexes and Their Enhancement of Hippocampal Plasticity. Biol. Psychiatry 2012, 71, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Narvaez, M.; Pérez-Alea, M.; Tarakanov, A.O.; Jiménez-Beristain, A.; Mudo’, G.; Agnati, L.F.; Ciruela, F.; Belluardo, N.; Fuxe, K. Evidence for the existence of FGFR1–5-HT1A heteroreceptor complexes in the midbrain raphe 5-HT system. Biochem. Biophys. Res. Commun. 2015, 456, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Pérez-Alea, M.; Narvaez, M.; Tarakanov, A.O.; Mudo’, G.; Jiménez-Beristain, A.; Agnati, L.F.; Ciruela, F.; Belluardo, N.; Fuxe, K. Enhancement of the FGFR1 signaling in the FGFR1-5-HT1A heteroreceptor complex in midbrain raphe 5-HT neuron systems. Relevance for neuroplasticity and depression. Biochem. Biophys. Res. Commun. 2015, 463, 180–186. [Google Scholar] [CrossRef] [Green Version]
- Casarotto, P.C.; Girych, M.; Fred, S.M.; Kovaleva, V.; Moliner, R.; Enkavi, G.; Biojone, C.; Cannarozzo, C.; Sahu, M.P.; Kaurinkoski, K.; et al. Antidepressant drugs act by directly binding to TRKB neurotrophin receptors. Cell 2021, 184, 1299–1313. [Google Scholar] [CrossRef]
- Duman, R.S.; Monteggia, L.M. A Neurotrophic Model for Stress-Related Mood Disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Castrén, E.; Antila, H. Neuronal plasticity and neurotrophic factors in drug responses. Mol. Psychiatry 2017, 22, 1085–1095. [Google Scholar] [CrossRef]
- Suzuki, S.; Kiyosue, K.; Hazama, S.; Ogura, A.; Kashihara, M.; Hara, T.; Koshimizu, H.; Kojima, M. Brain-Derived Neurotrophic Factor Regulates Cholesterol Metabolism for Synapse Development. J. Neurosci. 2007, 27, 6417–6427. [Google Scholar] [CrossRef] [PubMed]
- Sahu, M.P.; Pazos-Boubeta, Y.; Steinzeig, A.; Kaurinkoski, K.; Palmisano, M.; Borowecki, O.; Piepponen, T.P.; Castrén, E. Depletion of TrkB Receptors From Adult Serotonergic Neurons Increases Brain Serotonin Levels, Enhances Energy Metabolism and Impairs Learning and Memory. Front. Mol. Neurosci. 2021, 14, 616178. [Google Scholar] [CrossRef]
- Diniz, C.R.; Casarotto, P.C.; Fred, S.M.; Biojone, C.; Castrén, E.; Joca, S.R. Antidepressant-like effect of losartan involves TRKB transactivation from angiotensin receptor type 2 (AGTR2) and recruitment of FYN. Neuropharmacology 2018, 135, 163–171. [Google Scholar] [CrossRef]
- Samarajeewa, A.; Goldemann, L.; Vasefi, M.S.; Ahmed, N.; Gondora, N.; Khanderia, C.; Mielke, J.G.; Beazely, M.A. 5-HT7 receptor activation promotes an increase in TrkB receptor expression and phosphorylation. Front. Behav. Neurosci. 2014, 8, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rantamäki, T.; Vesa, L.; Antila, H.; Di Lieto, A.; Tammela, P.; Schmitt, A.; Lesch, K.-P.; Rios, M.; Castrén, E. Antidepressant Drugs Transactivate TrkB Neurotrophin Receptors in the Adult Rodent Brain Independently of BDNF and Monoamine Transporter Blockade. PLoS ONE 2011, 6, e20567. [Google Scholar] [CrossRef] [Green Version]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Garriga, P.; Ciruela, F.; Narvaez, M.; Tarakanov, A.O.; Palkovits, M.; Agnati, L.F.; Fuxe, K. G Protein–Coupled Receptor Heterodimerization in the Brain. Methods Enzymol. 2013, 521, 281–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Liberto, V.; Borroto-Escuela, D.; Frinchi, M.; Verdi, V.; Fuxe, K.; Belluardo, N.; Mudò, G. Existence of muscarinic acetylcholine receptor (mAChR) and fibroblast growth factor receptor (FGFR) heteroreceptor complexes and their enhancement of neurite outgrowth in neural hippocampal cultures. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2017, 1861, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Stachowiak, E.K.; Fang, X.; Myers, J.; Dunham, S.; Stachowiak, M.K. cAMP-induced differentiation of human neuronal progenitor cells is mediated by nuclear fibroblast growth factor receptor-1 (FGFR1). J. Neurochem. 2003, 84, 1296–1312. [Google Scholar] [CrossRef] [PubMed]
- Veena, J.; Srikumar, B.N.; Mahati, K.; Raju, T.R.; Rao, B.S.S. Oxotremorine treatment restores hippocampal neurogenesis and ameliorates depression-like behaviour in chronically stressed rats. Psychopharmacology 2011, 217, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Wydra, K.; Pintsuk, J.; Narvaez, M.; Corrales, F.; Zaniewska, M.; Agnati, L.F.; Franco, R.; Tanganelli, S.; Ferraro, L.; et al. Understanding the Functional Plasticity in Neural Networks of the Basal Ganglia in Cocaine Use Disorder: A Role for Allosteric Receptor-Receptor Interactions in A2A-D2 Heteroreceptor Complexes. Neural Plast. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Schlessinger, J. Ligand-Induced, Receptor-Mediated Dimerization and Activation of EGF Receptor. Cell 2002, 110, 669–672. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.-Q.; Zhang, L.-M.; Cao, Y.; Zhu, J.; Feng, L.-Y. Adenosine A1 receptor-mediated transactivation of the EGF receptor produces a neuroprotective effect on cortical neurons in vitro. Acta Pharmacol. Sin. 2009, 30, 889–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Buck, D.C.; Yang, R.; Macey, T.A.; Neve, K.A. Dopamine D2 receptor stimulation of mitogen-activated protein kinases mediated by cell type-dependent transactivation of receptor tyrosine kinases. J. Neurochem. 2005, 93, 899–909. [Google Scholar] [CrossRef]
- Li, B.; Zhang, S.; Zhang, H.; Nu, W.; Cai, L.; Hertz, L.; Peng, L. Fluoxetine-mediated 5-HT2B receptor stimulation in astrocytes causes EGF receptor transactivation and ERK phosphorylation. Psychopharmacology 2008, 201, 443–458. [Google Scholar] [CrossRef]
- Tsuchioka, M.; Takebayashi, M.; Hisaoka, K.; Maeda, N.; Nakata, Y. Serotonin (5-HT) induces glial cell line-derived neurotrophic factor (GDNF) mRNA expression via the transactivation of fibroblast growth factor receptor 2 (FGFR2) in rat C6 glioma cells. J. Neurochem. 2008, 106, 244–257. [Google Scholar] [CrossRef]
- Funa, K.; Sasahara, M. The Roles of PDGF in Development and During Neurogenesis in the Normal and Diseased Nervous System. J. Neuroimmune Pharmacol. 2013, 9, 168–181. [Google Scholar] [CrossRef] [Green Version]
- Kruk, J.S.; Vasefi, M.S.; Heikkila, J.J.; Beazely, M.A. Reactive Oxygen Species Are Required for 5-HT-Induced Transactivation of Neuronal Platelet-Derived Growth Factor and TrkB Receptors, but Not for ERK1/2 Activation. PLoS ONE 2013, 8, e77027. [Google Scholar] [CrossRef]
- Kruk, J.S.; Vasefi, M.S.; Liu, H.; Heikkila, J.J.; Beazely, M.A. 5-HT1A receptors transactivate the platelet-derived growth factor receptor type beta in neuronal cells. Cell. Signal. 2013, 25, 133–143. [Google Scholar] [CrossRef]
- Kruk, J.S.; Vasefi, M.S.; Gondora, N.; Ahmed, N.; Heikkila, J.J.; Beazely, M.A. Fluoxetine-induced transactivation of the platelet-derived growth factor type β receptor reveals a novel heterologous desensitization process. Mol. Cell. Neurosci. 2015, 65, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Kotecha, S.A.; Oak, J.N.; Jackson, M.F.; Perez, Y.; Orser, B.A.; Van Tol, H.H.; MacDonald, J.F. A D2 Class Dopamine Receptor Transactivates a Receptor Tyrosine Kinase to Inhibit NMDA Receptor Transmission. Neuron 2002, 35, 1111–1122. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Chu, X.-P.; Mao, L.-M.; Wang, M.; Lan, H.-X.; Li, M.-H.; Zhang, G.-C.; Parelkar, N.K.; Fibuch, E.E.; Haines, M.; et al. Modulation of D2R-NR2B Interactions in Response to Cocaine. Neuron 2006, 52, 897–909. [Google Scholar] [CrossRef] [Green Version]
- Heifets, B.D.; Malenka, R.C. MDMA as a Probe and Treatment for Social Behaviors. Cell 2016, 166, 269–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamilar-Britt, P.; Bedi, G. The prosocial effects of 3,4-methylenedioxymethamphetamine (MDMA): Controlled studies in humans and laboratory animals. Neurosci. Biobehav. Rev. 2015, 57, 433–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gregorio, D.; Aguilar-Valles, A.; Preller, K.H.; Heifets, B.D.; Hibicke, M.; Mitchell, J.; Gobbi, G. Hallucinogens in Mental Health: Preclinical and Clinical Studies on LSD, Psilocybin, MDMA, and Ketamine. J. Neurosci. 2020, 41, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Andén, N.E.; Corrodi, H.; Fuxe, K. Hallucinogenic drugs of the indolealkylamine type and central monoamine neurons. J. Pharmacol. Exp. Ther. 1971, 179, 236–249. [Google Scholar]
- Harfstrand, A.; Fuxe, K.; Cintra, A.; Agnati, L.F.; Zini, I.; Wikstrom, A.C.; Okret, S.; Yu, Z.Y.; Goldstein, M.; Steinbusch, H. Glucocorticoid receptor immunoreactivity in monoaminergic neurons of rat brain. Proc. Natl. Acad. Sci. USA 1986, 83, 9779–9783. [Google Scholar] [CrossRef] [Green Version]
- Tylš, F.; Páleníček, T.; Horacek, J. Psilocybin—Summary of knowledge and new perspectives. Eur. Neuropsychopharmacol. 2014, 24, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.C.; Marques, J.G. What is the clinical evidence on psilocybin for the treatment of psychiatric disorders? A systematic review. Porto Biomed. J. 2021, 6, e128. [Google Scholar] [CrossRef] [PubMed]
- Zanos, P.; Gould, T.D. Mechanisms of ketamine action as an antidepressant. Mol. Psychiatry 2018, 23, 801–811. [Google Scholar] [CrossRef]
- Saarelainen, T.; Hendolin, P.; Lucas, G.; Koponen, E.; Sairanen, M.; Macdonald, E.; Agerman, K.; Haapasalo, A.; Nawa, H.; Aloyz, R.; et al. Activation of the TrkB Neurotrophin Receptor Is Induced by Antidepressant Drugs and Is Required for Antidepressant-Induced Behavioral Effects. J. Neurosci. 2003, 23, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Kettenmann, H. Calcium signalling in glial cells. Trends Neurosci. 1996, 19, 346–352. [Google Scholar] [CrossRef]
- Fuxe, K.; Agnati, L.F.; Marcoli, M.; Escuela, D.O.B. Volume Transmission in Central Dopamine and Noradrenaline Neurons and Its Astroglial Targets. Neurochem. Res. 2015, 40, 2600–2614. [Google Scholar] [CrossRef]
- Volterra, A.; Meldolesi, J. Astrocytes, from brain glue to communication elements: The revolution continues. Nat. Rev. Neurosci. 2005, 6, 626–640. [Google Scholar] [CrossRef]
- Fuxe, K.; Dahlström, A.B.; Jonsson, G.; Marcellino, D.; Guescini, M.; Dam, M.; Manger, P.; Agnati, L. The discovery of central monoamine neurons gave volume transmission to the wired brain. Prog. Neurobiol. 2010, 90, 82–100. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Borroto-Escuela, D.O. Volume transmission and receptor-receptor interactions in heteroreceptor complexes: Understanding the role of new concepts for brain communication. Neural Regen. Res. 2016, 11, 1220–1223. [Google Scholar] [CrossRef] [PubMed]
- Parpura, V.; Basarsky, T.A.; Liu, F.; Jeftinija, K.; Jeftinija, S.; Haydon, P.G. Glutamate-mediated astrocyte–neuron signalling. Nat. Cell Biol. 1994, 369, 744–747. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, M. Direct signaling from astrocytes to neurons in cultures of mammalian brain cells. Science 1994, 263, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Parpura, V.; Haydon, P.G. Physiological astrocytic calcium levels stimulate glutamate release to modulate adjacent neurons. Proc. Natl. Acad. Sci. USA 2000, 97, 8629–8634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.-F.; Zhou, C.; Tang, A.-H.; Wang, S.-Q.; Chai, Z. Cellular mechanism for spontaneous calcium oscillations in astrocytes. Acta Pharmacol. Sin. 2006, 27, 861–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guthrie, P.B.; Knappenberger, J.; Segal, M.; Bennett, M.V.L.; Charles, A.C.; Kater, S.B. ATP Released from Astrocytes Mediates Glial Calcium Waves. J. Neurosci. 1999, 19, 520–528. [Google Scholar] [CrossRef]
- Mothet, J.-P.; Parent, A.T.; Wolosker, H.; Brady, R.O.; Linden, D.J.; Ferris, C.D.; Rogawski, M.; Snyder, S.H. D-Serine is an endogenous ligand for the glycine site of the N-methyl-D-aspartate receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 4926–4931. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Liu, Y.; Ye, Z.-R. Growth enhancement effect of BzATP on primary cultured astrocytes from rat brain. Neurosci. Bull. 2006, 22, 151–158. [Google Scholar]
- Bélanger, M.; Allaman, I.; Magistretti, P.J. Differential effects of pro- and anti-inflammatory cytokines alone or in combinations on the metabolic profile of astrocytes. J. Neurochem. 2011, 116, 564–576. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Allaman, I. Lactate in the brain: From metabolic end-product to signalling molecule. Nat. Rev. Neurosci. 2018, 19, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Pelassa, S.; Guidolin, D.; Venturini, A.; Averna, M.; Frumento, G.; Campanini, L.; Bernardi, R.; Cortelli, P.; Buonaura, G.C.; Maura, G.; et al. A2A-D2 Heteromers on Striatal Astrocytes: Biochemical and Biophysical Evidence. Int. J. Mol. Sci. 2019, 20, 2457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasull-Camós, J.; Gatius, M.T.; Artigas, F.; Castañé, A. Glial GLT-1 blockade in infralimbic cortex as a new strategy to evoke rapid antidepressant-like effects in rats. Transl. Psychiatry 2017, 7, e1038. [Google Scholar] [CrossRef] [Green Version]
- Zanos, P.; Moaddel, R.; Morris, P.; Georgiou, P.; Fischell, J.; Elmer, G.; Alkondon, M.; Yuan, P.; Pribut, H.J.; Singh, N.S.; et al. NMDAR inhibition-independent antidepressant actions of ketamine metabolites. Nat. Cell Biol. 2016, 533, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Artigas, F.; Bortolozzi, A.; Celada, P. Can we increase speed and efficacy of antidepressant treatments? Part I: General aspects and monoamine-based strategies. Eur. Neuropsychopharmacol. 2018, 28, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Gasull-Camós, J.; Martínez-Torres, S.; Gatius, M.T.; Ozaita, A.; Artigas, F.; Castañé, A. Serotonergic mechanisms involved in antidepressant-like responses evoked by GLT-1 blockade in rat infralimbic cortex. Neuropharmacology 2018, 139, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fullana, M.N.; Covelo, A.; Bortolozzi, A.; Araque, A.; Artigas, F. In vivo knockdown of astroglial glutamate transporters GLT-1 and GLAST increases excitatory neurotransmission in mouse infralimbic cortex: Relevance for depressive-like phenotypes. Eur. Neuropsychopharmacol. 2019, 29, 1288–1294. [Google Scholar] [CrossRef]
- Fullana, M.N.; Ruiz-Bronchal, E.; Ferrés-Coy, A.; Juarez-Escoto, E.; Artigas, F.; Bortolozzi, A. Regionally selective knockdown of astroglial glutamate transporters in infralimbic cortex induces a depressive phenotype in mice. Glia 2019, 67, 1122–1137. [Google Scholar] [CrossRef]
- Steinbusch, H.W.; Nieuwenhuys, R.; Verhofstad, A.A.; Van Der Kooy, D. The nucleus raphe dorsalis of the rat and its projection upon the caudatoputamen. A combined cytoarchitectonic, immunohistochemical and retrograde transport study. J. Physiol. 1981, 77, 157–174. [Google Scholar]
- Mo, J.; Kim, C.-H.; Lee, D.; Sun, W.; Lee, H.W.; Kim, H. Early growth response 1 (Egr-1) directly regulates GABAAreceptor α2, α4, and θ subunits in the hippocampus. J. Neurochem. 2015, 133, 489–500. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borroto-Escuela, D.O.; Ambrogini, P.; Narvaez, M.; Di Liberto, V.; Beggiato, S.; Ferraro, L.; Fores-Pons, R.; Alvarez-Contino, J.E.; Lopez-Salas, A.; Mudò, G.; et al. Serotonin Heteroreceptor Complexes and Their Integration of Signals in Neurons and Astroglia—Relevance for Mental Diseases. Cells 2021, 10, 1902. https://doi.org/10.3390/cells10081902
Borroto-Escuela DO, Ambrogini P, Narvaez M, Di Liberto V, Beggiato S, Ferraro L, Fores-Pons R, Alvarez-Contino JE, Lopez-Salas A, Mudò G, et al. Serotonin Heteroreceptor Complexes and Their Integration of Signals in Neurons and Astroglia—Relevance for Mental Diseases. Cells. 2021; 10(8):1902. https://doi.org/10.3390/cells10081902
Chicago/Turabian StyleBorroto-Escuela, Dasiel O., Patrizia Ambrogini, Manuel Narvaez, Valentina Di Liberto, Sarah Beggiato, Luca Ferraro, Ramon Fores-Pons, Jose E. Alvarez-Contino, Alexander Lopez-Salas, Giuseppa Mudò, and et al. 2021. "Serotonin Heteroreceptor Complexes and Their Integration of Signals in Neurons and Astroglia—Relevance for Mental Diseases" Cells 10, no. 8: 1902. https://doi.org/10.3390/cells10081902