
The Transformation of the Centrosome into the Basal Body: Similarities and Dissimilarities between Somatic and Male Germ Cells and Their Relevance for Male Fertility


Abstract
:1. Introduction
2. The Centrosome in Somatic Cells and Its Transformation into the Basal Body
3. Building the Connecting Piece and Its Ultrastructure
4. The Protein Components of the Connecting Piece and Their Relevance for the Head-to-Tail Linkage and Male Fertility
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oakberg, E.F. A Description of Spermiogenesis in the Mouse and Its Use in Analysis of the Cycle of the Seminiferous Epithelium and Germ Cell Renewal. Am. J. Anat. 1956, 99, 391–413. [Google Scholar] [CrossRef] [PubMed]
- Clermont, Y. Kinetics of Spermatogenesis in Mammals: Seminiferous Epithelium Cycle and Spermatogonial Renewal. Physiol. Rev. 1972, 52, 198–236. [Google Scholar] [CrossRef]
- Russell, L.D. Histological and Histopathological Evaluation of the Testis; Cache River Press, 1990; ISBN 9780962742200. [Google Scholar]
- de Rooij, D.G.; Grootegoed, J.A. Spermatogonial Stem Cells. Curr. Opin. Cell Biol. 1998, 10, 694–701. [Google Scholar] [CrossRef]
- Hermo, L.; Pelletier, R.-M.; Cyr, D.G.; Smith, C.E. Surfing the Wave, Cycle, Life History, and Genes/Proteins Expressed by Testicular Germ Cells. Part 1: Background to Spermatogenesis, Spermatogonia, and Spermatocytes. Microsc. Res. Tech. 2010, 73, 241–278. [Google Scholar] [CrossRef]
- Mortimer, D. The Functional Anatomy of the Human Spermatozoon: Relating Ultrastructure and Function. Mhr Basic Sci. Reprod. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, D.W. The Mammalian Spermatozoon. Dev. Biol. 1975, 44, 394–436. [Google Scholar] [CrossRef]
- Chemes, H.E.; Rawe, V.Y. The Making of Abnormal Spermatozoa: Cellular and Molecular Mechanisms Underlying Pathological Spermiogenesis. Cell Tissue Res. 2010, 341, 349–357. [Google Scholar] [CrossRef]
- Kierszenbaum, A.L. Intramanchette Transport (IMT): Managing the Making of the Spermatid Head, Centrosome, and Tail. Mol. Reprod. Dev. 2002, 63, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, J.L.; Witman, G.B. Intraflagellar Transport. Nat. Rev. Mol. Cell Biol. 2002, 3, 813–825. [Google Scholar] [CrossRef]
- Pedersen, L.B.; Rosenbaum, J.L. Intraflagellar Transport (IFT) Role in Ciliary Assembly, Resorption and Signalling. Curr. Top. Dev. Biol. 2008, 85, 23–61. [Google Scholar] [CrossRef]
- Kierszenbaum, A.L.; Rivkin, E.; Tres, L.L. Cytoskeletal Track Selection during Cargo Transport in Spermatids Is Relevant to Male Fertility. Spermatogenesis 2011, 1, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, L.; O’Bryan, M.K. Microtubules and Spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 45–54. [Google Scholar] [CrossRef]
- Rattner, J.B.; Olson, G. Observations on the Fine Structure of the Nuclear Ring of the Mammalian Spermatid. J. Ultrastruct. Res. 1973, 43, 438–444. [Google Scholar] [CrossRef]
- Baltz, J.M.; Oneeka Williams, P.; Cone, R.A. Dense Fibers Protect Mammalian Sperm Against Damage. Biol. Reprod. 1990, 43, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindemann, C.B. Functional Significance of the Outer Dense Fibers of Mammalian Sperm Examined by Computer Simulations with the Geometric Clutch Model. Cell Motil. Cytoskelet. 1996, 34, 258–270. [Google Scholar] [CrossRef]
- Chemes, H.E.; Alvarez Sedo, C. Tales of the Tail and Sperm Head AchesChanging Concepts on the Prognostic Significance of Sperm Pathologies Affecting the Head, Neck and Tail. Asian J. 2012, 14, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Hoyer-Fender, S. Centrosomes in Fertilization, Early Embryonic Development, Stem Cell Division, and Cancer. Atlas Genet. Cytogenet. Oncol. Haematol. 2012. [Google Scholar] [CrossRef]
- Van Blerkom, J. Sperm Centrosome Dysfunction: A Possible New Class of Male Factor Infertility in the Human. Mol. Hum. Reprod. 1996, 2, 349–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, Z.P. Sperm Centriole Disfunction and Sperm Immotility. Mol. Cell. Endocrinol. 2000, 166, 59–62. [Google Scholar] [CrossRef]
- Terada, Y.; Schatten, G.; Hasegawa, H.; Yaegashi, N. Essential Roles of the Sperm Centrosome in Human Fertilization: Developing the Therapy for Fertilization Failure Due to Sperm Centrosomal Dysfunction. Tohoku J. Exp. Med. 2010, 220, 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorobjev, I.A.; Chentsov, Y.S. The Ultrastructure of Centriole in Mammalian Tissue Culture Cells. Cell Biol. Int. Rep. 1980, 4, 1037–1044. [Google Scholar] [CrossRef]
- Kimble, M.; Kuriyama, R. Functional Components Of Microtubule-Organizing Centers. In International Review of Cytology; Jeon, K.W., Friedlander, M., Eds.; Academic Press: Cambridge, MA, USA, 1992; Volume 136, pp. 1–50. [Google Scholar]
- Kalt, A.; Schliwa, M. Molecular Components of the Centrosome. Trends Cell Biol. 1993, 3, 118–128. [Google Scholar] [CrossRef]
- Kellogg, D.R.; Moritz, M.; Alberts, B.M. The Centrosome and Cellular Organization. Annu. Rev. Biochem. 1994, 63, 639–674. [Google Scholar] [CrossRef]
- Zheng, Y.; Wong, M.L.; Alberts, B.; Mitchison, T. Nucleation of Microtubule Assembly by a γ-Tubulin-Containing Ring Complex. Nature 1995, 378, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Moritz, M.; Braunfeld, M.B.; Sedat, J.W.; Alberts, B.; Agard, D.A. Microtubule Nucleation by Gamma-Tubulin-Containing Rings in the Centrosome. Nature 1995, 378, 638–640. [Google Scholar] [CrossRef] [PubMed]
- Stearns, T.; Evans, L.; Kirschner, M. γ-Tubulin Is a Highly Conserved Component of the Centrosome. Cell 1991, 65, 825–836. [Google Scholar] [CrossRef]
- Joshi, H.C.; Palacios, M.J.; McNamara, L.; Cleveland, D.W. γ-Tubulin Is a Centrosomal Protein Required for Cell Cycle-Dependent Microtubule Nucleation. Nature 1992, 356, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Stearns, T.; Kirschner, M. In Vitro Reconstitution of Centrosome Assembly and Function: The Central Role of γ-Tubulin. Cell 1994, 76, 623–637. [Google Scholar] [CrossRef]
- Doxsey, S.J.; Stein, P.; Evans, L.; Calarco, P.D.; Kirschner, M. Pericentrin, a Highly Conserved Centrosome Protein Involved in Microtubule Organization. Cell 1994, 76, 639–650. [Google Scholar] [CrossRef]
- Dictenberg, J.B.; Zimmerman, W.; Sparks, C.A.; Young, A.; Vidair, C.; Zheng, Y.; Carrington, W.; Fay, F.S.; Doxsey, S.J. Pericentrin and γ-Tubulin Form a Protein Complex and Are Organized into a Novel Lattice at the Centrosome. J. Cell Biol. 1998, 141, 12. [Google Scholar] [CrossRef]
- Ou, Y.; Rattner, J.B. A Subset of Centrosomal Proteins Are Arranged in a Tubular Conformation That Is Reproduced during Centrosome Duplication. Cell Motil. Cytoskelet. 2000, 47, 13–24. [Google Scholar] [CrossRef]
- Ou, Y.Y.; Zhang, M.; Chi, S.; Matyas, J.R.; Rattner, J.B. Higher Order Structure of the PCM Adjacent to the Centriole. Cell Motil. Cytoskelet. 2003, 55, 125–133. [Google Scholar] [CrossRef]
- Mennella, V.; Keszthelyi, B.; McDonald, K.L.; Chhun, B.; Kan, F.; Rogers, G.C.; Huang, B.; Agard, D.A. Subdiffraction-Resolution Fluorescence Microscopy Reveals a Domain of the Centrosome Critical for Pericentriolar Material Organization. Nat. Cell Biol. 2012, 14, 1159–1168. [Google Scholar] [CrossRef] [Green Version]
- Lawo, S.; Hasegan, M.; Gupta, G.D.; Pelletier, L. Subdiffraction Imaging of Centrosomes Reveals Higher-Order Organizational Features of Pericentriolar Material. Nat. Cell Biol. 2012, 14, 1148–1158. [Google Scholar] [CrossRef] [PubMed]
- Tsou, M.-F.B.; Stearns, T. Controlling Centrosome Number: Licenses and Blocks. Curr. Opin. Cell Biol. 2006, 18, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Tsou, M.-F.B.; Stearns, T. Mechanism Limiting Centrosome Duplication to Once per Cell Cycle. Nature 2006, 442, 947–951. [Google Scholar] [CrossRef]
- Nigg, E.A.; Stearns, T. The Centrosome Cycle: Centriole Biogenesis, Duplication and Inherent Asymmetries. Nat. Cell Biol. 2011, 13, 1154–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigg, E.A.; Holland, A.J. Once and Only Once: Mechanisms of Centriole Duplication and Their Deregulation in Disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Paintrand, M.; Moudjou, M.; Delacroix, H.; Bornens, M. Centrosome Organization and Centriole Architecture: Their Sensitivity to Divalent Cations. J. Struct. Biol. 1992, 108, 107–128. [Google Scholar] [CrossRef]
- Piel, M.; Meyer, P.; Khodjakov, A.; Rieder, C.L.; Bornens, M. The Respective Contributions of the Mother and Daughter Centrioles to Centrosome Activity and Behavior in Vertebrate Cells. J. Cell Biol. 2000, 149, 317–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, R.; Messaoudi, C.; Chichon, F.J.; Celati, C.; Marco, S. Electron Tomography Study of Isolated Human Centrioles. Microsc. Res. Tech. 2009, 72, 42–48. [Google Scholar] [CrossRef]
- Dawe, H.R.; Farr, H.; Gull, K. Centriole/Basal Body Morphogenesis and Migration during Ciliogenesis in Animal Cells. J. Cell Sci. 2006, 120, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenney, J.; Karsenti, E.; Gowen, B.; Fuller, S.D. Three-Dimensional Reconstruction of the Mammalian Centriole from Cryoelectron Micrographs: The Use of Common Lines for Orientation and Alignment. J. Struct. Biol. 1997, 120, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Tylkowski, M.A.; Hüber, D.; Contreras, C.T.; Hoyer-Fender, S. ODF2 Maintains Centrosome Cohesion by Restricting β-Catenin Accumulation. J. Cell Sci. 2018, 131, jcs220954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowler, M.; Kong, D.; Sun, S.; Nanjundappa, R.; Evans, L.; Farmer, V.; Holland, A.; Mahjoub, M.R.; Sui, H.; Loncarek, J. High-Resolution Characterization of Centriole Distal Appendage Morphology and Dynamics by Correlative STORM and Electron Microscopy. Nat. Commun. 2019, 10, 993. [Google Scholar] [CrossRef]
- Kodani, A.; Salomé Sirerol-Piquer, M.; Seol, A.; Manuel Garcia-Verdugo, J.; Reiter, J.F. Kif3a Interacts with Dynactin Subunit P150 Glued to Organize Centriole Subdistal Appendages. Embo J. 2013, 32, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Garcia, G.; Reiter, J.F. A Primer on the Mouse Basal Body. Cilia 2016, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Reiter, J. How the Centriole Builds Its Cilium: Of Mothers, Daughters, and the Acquisition of Appendages. Curr. Opin. Struct. Biol. 2021, 66, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Hoyer-Fender, S.P.; Motile, C. Their Ultrastructure and Ciliogenesis. In Cilia and Nervous System Development and Function; Tucker, K.L., Caspary, T., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 1–53. ISBN 9789400758087. [Google Scholar]
- Uzbekov, R.; Alieva, I. Who Are You, Subdistal Appendages of Centriole? Open Biol. 2018, 8, 180062. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, I.R. The relationship between the fine structure and direction of beat in gill cilia of a lamellibranch mollusc. J. Biophys. Biochem. Biol. 1961, 11, 179–205. [Google Scholar] [CrossRef] [Green Version]
- Tateishi, K.; Yamazaki, Y.; Nishida, T.; Watanabe, S.; Kunimoto, K.; Ishikawa, H.; Tsukita, S. Two Appendages Homologous between Basal Bodies and Centrioles Are Formed Using Distinct Odf2 Domains. J. Cell Biol. 2013, 203, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Kunimoto, K.; Yamazaki, Y.; Nishida, T.; Shinohara, K.; Ishikawa, H.; Hasegawa, T.; Okanoue, T.; Hamada, H.; Noda, T.; Tamura, A.; et al. Coordinated Ciliary Beating Requires Odf2-Mediated Polarization of Basal Bodies via Basal Feet. Cell 2012, 148, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, H.; Kano, A.; Aoki, T.; Ohwada, N. Immunocytochemistry of the Striated Rootlets Associated with Solitary Cilia in Human Oviductal Secretory Cells. Histochem. Cell Biol. 2000, 114, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, T.W. Zur Anatomie und Physiologie der Flimmerzellen. Pflüg. Arch. Für Gesammte Physiol. Menschen Thiere 1880, 23, 505–535. [Google Scholar] [CrossRef]
- Sjöstrand, F.S. The Ultrastructure of the Inner Segments of the Retinal Rods of the Guinea Pig Eye as Revealed by Electron Microscopy: Ultrastructure of rod inner segments. J. Cell. Comp. Physiol. 1953, 42, 45–70. [Google Scholar] [CrossRef]
- Fawcett, D.W.; Porter, K.R. A Study of the Fine Structure of Ciliated Epithelia. J. Morphol. 1954, 94, 221–281. [Google Scholar] [CrossRef]
- Spira, A.W.; Milman, G.E. The Structure and Distribution of the Cross-Striated Fibril and Associated Membranes in Guinea Pig Photoreceptors. Am. J. Anat. 1979, 155, 319–337. [Google Scholar] [CrossRef] [PubMed]
- Sandoz, D.; Chailley, B.; Boisvieux-Ulrich, E.; Lemullois, M.; Laine, M.-C.; Bautista-Harris, G. Organization and Functions of Cytoskeleton in Metazoan Ciliated Cells. Biol. Cell 1988, 63, 183–193. [Google Scholar] [CrossRef]
- Hagiwara, H.; Aoki, T.; Ohwada, N.; Fujimoto, T. Development of Striated Rootlets during Ciliogenesis in the Human Oviduct Epithelium. Cell Tissue Res. 1997, 290, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, X.; Yue, G.; Adamian, M.; Bulgakov, O.; Li, T. Rootletin, a Novel Coiled-Coil Protein, Is a Structural Component of the Ciliary Rootlet. J. Cell Biol. 2002, 159, 431–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, J.S.; Wilkinson, C.J.; Mayor, T.; Mortensen, P.; Nigg, E.A.; Mann, M. Proteomic Characterization of the Human Centrosome by Protein Correlation Profiling. Nature 2003, 426, 570–574. [Google Scholar] [CrossRef]
- Fry, A.M.; Mayor, T.; Meraldi, P.; Stierhof, Y.-D.; Tanaka, K.; Nigg, E.A. C-Nap1, a Novel Centrosomal Coiled-Coil Protein and Candidate Substrate of the Cell Cycle–Regulated Protein Kinase Nek2. J. Cell Biol. 1998, 141, 1563–1574. [Google Scholar] [CrossRef] [Green Version]
- Mayor, T.; Stierhof, Y.-D.; Tanaka, K.; Fry, A.M.; Nigg, E.A. The Centrosomal Protein C-Nap1 Is Required for Cell Cycle–Regulated Centrosome Cohesion. J. Cell Biol. 2000, 151, 837–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornens, M. Centrosome Composition and Microtubule Anchoring Mechanisms. Curr. Opin. Cell Biol. 2002, 14, 25–34. [Google Scholar] [CrossRef]
- Bahe, S.; Stierhof, Y.-D.; Wilkinson, C.J.; Leiss, F.; Nigg, E.A. Rootletin Forms Centriole-Associated Filaments and Functions in Centrosome Cohesion. J. Cell Biol. 2005, 171, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Adamian, M.; Li, T. Rootletin Interacts with C-Nap1 and May Function as a Physical Linker between the Pair of Centrioles/Basal Bodies in Cells. Mol. Biol. Cell 2006, 17, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Flory, M.R.; Moser, M.J.; Monnat, R.J.; Davis, T.N. Identification of a Human Centrosomal Calmodulin-Binding Protein That Shares Homology with Pericentrin. Proc. Natl. Acad. Sci. USA 2000, 97, 5919–5923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flory, M.R.; Davis, T.N. The Centrosomal Proteins Pericentrin and Kendrin Are Encoded by Alternatively Spliced Products of One Gene. Genomics 2003, 82, 401–405. [Google Scholar] [CrossRef]
- Dammermann, A.; Merdes, A. Assembly of Centrosomal Proteins and Microtubule Organization Depends on PCM-1. J. Cell Biol. 2002, 159, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graser, S.; Stierhof, Y.-D.; Lavoie, S.B.; Gassner, O.S.; Lamla, S.; Le Clech, M.; Nigg, E.A. Cep164, a Novel Centriole Appendage Protein Required for Primary Cilium Formation. J. Cell Biol. 2007, 179, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Ferreria, M.A.; Rath, U.; Buster, D.W.; Chanda, S.K.; Caldwell, J.S.; Rines, D.R.; Sharp, D.J. Human Cep192 Is Required for Mitotic Centrosome and Spindle Assembly. Curr. Biol. 2007, 17, 1960–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Lawo, S.; Bird, A.; Pinchev, D.; Ralph, A.; Richter, C.; Müller-Reichert, T.; Kittler, R.; Hyman, A.A.; Pelletier, L. The Mammalian SPD-2 Ortholog Cep192 Regulates Centrosome Biogenesis. Curr. Biol. 2008, 18, 136–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, K.-W.; Choi, Y.-K.; Rattner, J.B.; Qi, R.Z. CDK5RAP2 Is a Pericentriolar Protein That Functions in Centrosomal Attachment of the γ-Tubulin Ring Complex. Mol. Biol. Cell 2008, 19, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, V.D.; Huang, B. Molecular Cloning and Centrosomal Localization of Human Caltractin. Proc. Natl. Acad. Sci. USA 1993, 90, 11039–11043. [Google Scholar] [CrossRef] [Green Version]
- Satisbury, J.L. Centrin, Centrosomes, and Mitotic Spindle Poles. Curr. Opin. Cell Biol. 1995, 7, 39–45. [Google Scholar] [CrossRef]
- Paoletti, A.; Moudjou, M.; Paintrand, M.; Salisbury, J.L.; Bornens, M. Most of Centrin in Animal Cells Is Not Centrosome-Associated and Centrosomal Centrin Is Confined to the Distal Lumen of Centrioles. J. Cell Sci. 1996, 109, 3089–3102. [Google Scholar] [CrossRef]
- Dantas, T.J.; Daly, O.M.; Morrison, C.G. Such Small Hands: The Roles of Centrins/Caltractins in the Centriole and in Genome Maintenance. Cell. Mol. Life Sci. 2012, 69, 2979–2997. [Google Scholar] [CrossRef] [PubMed]
- Le Guennec, M.; Klena, N.; Gambarotto, D.; Laporte, M.H.; Tassin, A.-M.; van den Hoek, H.; Erdmann, P.S.; Schaffer, M.; Kovacik, L.; Borgers, S.; et al. A Helical Inner Scaffold Provides a Structural Basis for Centriole Cohesion. Sci. Adv. 2020, 6, eaaz4137. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, L.; Vanselow, K.; Skogs, M.; Toyoda, Y.; Lundberg, E.; Poser, I.; Falkenby, L.G.; Bennetzen, M.; Westendorf, J.; Nigg, E.A.; et al. Novel Asymmetrically Localizing Components of Human Centrosomes Identified by Complementary Proteomics Methods. Embo J. 2011, 30, 1520–1535. [Google Scholar] [CrossRef]
- Nogales-Cadenas, R.; Abascal, F.; Díez-Pérez, J.; Carazo, J.M.; Pascual-Montano, A. CentrosomeDB: A Human Centrosomal Proteins Database. Nucleic Acids Res. 2009, 37, 175–180. [Google Scholar] [CrossRef]
- Alves-Cruzeiro, J.M.d.C.; Nogales-Cadenas, R.; Pascual-Montano, A.D. CentrosomeDB: A New Generation of the Centrosomal Proteins Database for Human and Drosophila Melanogaster. Nucleic Acids Res. 2014, 42, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Rajendran, V.; Sethumadhavan, R.; Purohit, R. CEP Proteins: The Knights of Centrosome Dynasty. Protoplasma 2013, 250, 965–983. [Google Scholar] [CrossRef]
- Sullenberger, C.; Vasquez-Limeta, A.; Kong, D.; Loncarek, J. With Age Comes Maturity: Biochemical and Structural Transformation of a Human Centriole in the Making. Cells 2020, 9, 1429. [Google Scholar] [CrossRef]
- Zou, C.; Li, J.; Bai, Y.; Gunning, W.T.; Wazer, D.E.; Band, V.; Gao, Q. Centrobin: A Novel Daughter Centriole–Associated Protein That Is Required for Centriole Duplication. J. Cell Biol. 2005, 171, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Mahjoub, M.R.; Xie, Z.; Stearns, T. Cep120 Is Asymmetrically Localized to the Daughter Centriole and Is Essential for Centriole Assembly. J. Cell Biol. 2010, 191, 331–346. [Google Scholar] [CrossRef] [Green Version]
- Gudi, R.; Zou, C.; Li, J.; Gao, Q. Centrobin–Tubulin Interaction Is Required for Centriole Elongation and Stability. J. Cell Biol. 2011, 193, 711–725. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.-J.C.; Lin, S.-Y.; Hsu, W.-B.; Lin, Y.-N.; Wu, C.-T.; Lin, Y.-C.; Chang, C.-W.; Wu, K.-S.; Tang, T.K. The Human Microcephaly Protein STIL Interacts with CPAP and Is Required for Procentriole Formation: STIL Is Required for Procentriole Formation. Embo J. 2011, 30, 4790–4804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, W.Y.; Bossard, C.; Khanna, H.; Peränen, J.; Swaroop, A.; Malhotra, V.; Dynlacht, B.D. CP110 Suppresses Primary Cilia Formation through Its Interaction with CEP290, a Protein Deficient in Human Ciliary Disease. Dev. Cell 2008, 15, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Spektor, A.; Tsang, W.Y.; Khoo, D.; Dynlacht, B.D. Cep97 and CP110 Suppress a Cilia Assembly Program. Cell 2007, 130, 678–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojarad, B.A.; Gupta, G.D.; Hasegan, M.; Goudiam, O.; Basto, R.; Gingras, A.-C.; Pelletier, L. CEP19 Cooperates with FOP and CEP350 to Drive Early Steps in the Ciliogenesis Programme. Open Biol. 2017, 7, 170114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Zeng, H.; Ning, G.; Reiter, J.F.; Liu, A. C2cd3 Is Critical for Centriolar Distal Appendage Assembly and Ciliary Vesicle Docking in Mammals. Proc. Natl. Acad. Sci. USA 2014, 111, 2164–2169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.L.; Santé, J.; Comerci, C.J.; Cyge, B.; Menezes, L.F.; Li, F.-Q.; Germino, G.G.; Moerner, W.E.; Takemaru, K.-I.; Stearns, T. Cby1 Promotes Ahi1 Recruitment to a Ring-Shaped Domain at the Centriole–Cilium Interface and Facilitates Proper Cilium Formation and Function. Mol. Biol. Cell 2014, 25, 2919–2933. [Google Scholar] [CrossRef]
- Singla, V.; Romaguera-Ros, M.; Garcia-Verdugo, J.M.; Reiter, J.F. Ofd1, a Human Disease Gene, Regulates the Length and Distal Structure of Centrioles. Dev. Cell 2010, 18, 410–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sillibourne, J.E.; Hurbain, I.; Grand-Perret, T.; Goud, B.; Tran, P.; Bornens, M. Primary Ciliogenesis Requires the Distal Appendage Component Cep123. Biol. Open 2013, 2, 535–545. [Google Scholar] [CrossRef] [Green Version]
- Tanos, B.E.; Yang, H.-J.; Soni, R.; Wang, W.-J.; Macaluso, F.P.; Asara, J.M.; Tsou, M.-F.B. Centriole Distal Appendages Promote Membrane Docking, Leading to Cilia Initiation. Genes Dev. 2013, 27, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Joo, K.; Kim, C.G.; Lee, M.-S.; Moon, H.-Y.; Lee, S.-H.; Kim, M.J.; Kweon, H.-S.; Park, W.-Y.; Kim, C.-H.; Gleeson, J.G.; et al. CCDC41 Is Required for Ciliary Vesicle Docking to the Mother Centriole. Proc. Natl. Acad. Sci. USA 2013, 110, 5987–5992. [Google Scholar] [CrossRef] [Green Version]
- Guarguaglini, G.; Duncan, P.I.; Stierhof, Y.D.; Holmström, T.; Duensing, S.; Nigg, E.A. The Forkhead-Associated Domain Protein Cep170 Interacts with Polo-like Kinase 1 and Serves as a Marker for Mature Centrioles. Mol. Biol. Cell 2005, 16, 1095–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, B.M.; Gull, K. A Molecular Marker for Centriole Maturation in the Mammalian Cell Cycle. J. Cell Biol. 1995, 130, 919–927. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, Y.; Yamane, Y.; Okanoue, T.; Tsukita, S.; Tsukita, S. Outer Dense Fiber 2 Is a Widespread Centrosome Scaffold Component Preferentially Associated with Mother Centrioles: Its Identification from Isolated Centrosomes. Mol. Biol. Cell 2001, 12, 1687–1697. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, H.; Kubo, A.; Tsukita, S.; Tsukita, S. Odf2-Deficient Mother Centrioles Lack Distal/Subdistal Appendages and the Ability to Generate Primary Cilia. Nat. Cell Biol. 2005, 7, 517–524. [Google Scholar] [CrossRef]
- Mönnich, M.; Borgeskov, L.; Breslin, L.; Jakobsen, L.; Rogowski, M.; Doganli, C.; Schrøder, J.M.; Mogensen, J.B.; Blinkenkjær, L.; Harder, L.M.; et al. CEP128 Localizes to the Subdistal Appendages of the Mother Centriole and Regulates TGF-β/BMP Signaling at the Primary Cilium. Cell Rep. 2018, 22, 2584–2592. [Google Scholar] [CrossRef] [Green Version]
- Gromley, A.; Jurczyk, A.; Sillibourne, J.; Halilovic, E.; Mogensen, M.; Groisman, I.; Blomberg, M.; Doxsey, S. A Novel Human Protein of the Maternal Centriole Is Required for the Final Stages of Cytokinesis and Entry into S Phase. J. Cell Biol. 2003, 161, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.; Stearns, T. Delta-Tubulin and Epsilon-Tubulin: Two New Human Centrosomal Tubulins Reveal New Aspects of Centrosome Structure and Function. Nat. Cell Biol. 2000, 2, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.; Giddings, T.H.; Winey, M.; Stearns, T. ε-Tubulin Is Required for Centriole Duplication and Microtubule Organization. Nat. Cell Biol. 2003, 5, 71–76. [Google Scholar] [CrossRef]
- Bouckson-Castaing, V.; Moudjou, M.; Ferguson, D.J.; Mucklow, S.; Belkaid, Y.; Milon, G.; Crocker, P.R. Molecular Characterisation of Ninein, a New Coiled-Coil Protein of the Centrosome. J. Cell Sci. 1996, 109, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.R.; Chen, C.H.; Chang, J.H.; Wang, S.; Sy, W.D.; Chou, C.K.; Howng, S.L. Cloning and Characterization of a Novel Human Ninein Protein That Interacts with the Glycogen Synthase Kinase 3beta. Biochim. Biophys. Acta 2000, 1492, 513–516. [Google Scholar] [CrossRef]
- Mogensen, M.M.; Malik, A.; Piel, M.; Bouckson-Castaing, V.; Bornens, M. Microtubule Minus-End Anchorage at Centrosomal and Non-Centrosomal Sites: The Role of Ninein. J. Cell Sci. 2000, 113, 3013–3023. [Google Scholar] [CrossRef]
- Delgehyr, N.; Sillibourne, J.; Bornens, M. Microtubule Nucleation and Anchoring at the Centrosome Are Independent Processes Linked by Ninein Function. J. Cell Sci. 2005, 118, 1565–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashihara, H.; Chiba, S.; Kanno, S.; Suzuki, K.; Yano, T.; Tsukita, S. Cep128 Associates with Odf2 to Form the Subdistal Appendage of the Centriole. Genes Cells 2019, 24, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Chong, W.M.; Wang, W.-J.; Lo, C.-H.; Chiu, T.-Y.; Chang, T.-J.; Liu, Y.-P.; Tanos, B.; Mazo, G.; Tsou, M.-F.B.; Jane, W.-N.; et al. Super-Resolution Microscopy Reveals Coupling between Mammalian Centriole Subdistal Appendages and Distal Appendages. eLife 2020, 9, e53580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzbekov, E.; Avidor-Reiss, T. Principal postulates of centrosomal biology. Version 2020. Cells 2020, 9, 2156. [Google Scholar] [CrossRef]
- Breslow, D.K.; Holland, A.J. Mechanism and Regulation of Centriole and Cilium Biogenesis. Annu. Rev. Biochem. 2019, 88, 691–724. [Google Scholar] [CrossRef] [PubMed]
- Reiter, J.F.; Leroux, M.R. Genes and Molecular Pathways Underpinning Ciliopathies. Nat. Rev. Mol. Cell Biol. 2017, 18, 533–547. [Google Scholar] [CrossRef]
- Fawcett, D.W. The Anatomy of the Mammalian Spermatozoon with Particular Reference to the Guinea Pig. Z. Für Zellforsch. Mikrosk. Anat. 1965, 67, 279–296. [Google Scholar] [CrossRef]
- Fawcett, D.W.; Ito, S. The Fine Structure of Bat Spermatozoa. Am. J. Anat. 1965, 116, 567–609. [Google Scholar] [CrossRef]
- Fawcett, D.W.; Phillips, D.M. The Fine Structure and Development of the Neck Region of the Mammalian Spermatozoon. Anat. Rec. 1969, 165, 153–183. [Google Scholar] [CrossRef]
- de Kretser, D.M. Ultrastructural Features of Human Spermiogenesis. Z. Für Zellforsch. Mikrosk. Anat. 1969, 98, 477–505. [Google Scholar] [CrossRef]
- Nicander, L.; Bane, A. Fine Structure of Boar Spermatozoa. Z. Für Zellforsch. Mikrosk. Anat. 1962, 57, 390–405. [Google Scholar] [CrossRef] [PubMed]
- Aire, T.A. Spermiogenesis in Birds. Spermatogenesis 2014, 4, e959392. [Google Scholar] [CrossRef]
- Zamboni, L.; Stefanini, M. The Fine Structure of the Neck of Mammalian Spermatozoa. Anat. Rec. 1971, 169, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M. The Distal Centriole in Guinea Pig Spermiogenesis. J. Ultrastruct. Res. 1972, 39, 364–388. [Google Scholar] [CrossRef]
- Irons, M.J. Synthesis and Assembly of Connecting-Piece Proteins as Revealed by Radioautography. J. Ultrastruct. Res. 1983, 82, 27–34. [Google Scholar] [CrossRef]
- Nagano, T. Observations On The Fine Structure Of The Developing Spermatid In The Domestic Chicken. J. Cell Biol. 1962, 14, 193–205. [Google Scholar] [CrossRef]
- Garanina, A.S.; Alieva, I.B.; Bragina, E.E.; Blanchard, E.; Arbeille, B.; Guerif, F.; Uzbekova, S.V.; Uzbekov, R.E. The centriolar adjunct-appearance and disassembly in spermiogenesis and the potential impact on fertility. Cells 2019, 8, 180. [Google Scholar] [CrossRef] [Green Version]
- Avidor-Reiss, T. Rapid Evolution of Sperm Produces Diverse Centriole Structures That Reveal the Most Rudimentary Structure Needed for Function. Cells 2018, 7, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolley, D.M.; Fawcett, D.W. The Degeneration and Disappearance of the Centrioles during the Development of the Rat Spermatozoon. Anat. Rec. 1973, 177, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, G.; Sutovsky, P.; Joshi, H.C.; Stearns, T.; Schatten, G. Centrosome Reduction during Mouse Spermiogenesis. Dev. Biol. 1998, 203, 424–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manandhar, G. Highly Degenerated Distal Centrioles in Rhesus and Human Spermatozoa. Hum. Reprod. 2000, 15, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Fishman, E.L.; Jo, K.; Nguyen, Q.P.H.; Kong, D.; Royfman, R.; Cekic, A.R.; Khanal, S.; Miller, A.L.; Simerly, C.; Schatten, G.; et al. A Novel Atypical Sperm Centriole Is Functional during Human Fertilization. Nat. Commun. 2018, 9, 2210. [Google Scholar] [CrossRef]
- Saacke, R.G.; Almquist, J.O. Ultrastructure of Bovine Spermatozoa. II. The Neck and Tail of Normal, Ejaculated Sperm. Am. J. Anat. 1964, 115, 163–183. [Google Scholar] [CrossRef]
- Pedersen, H. Ultrastructure of the Ejaculated Human Sperm. Z. Für Zellforsch. Mikrosk. Anat. 1969, 94, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Ounjai, P.; Kim, K.D.; Lishko, P.V.; Downing, K.H. Three-Dimensional Structure of the Bovine Sperm Connecting Piece Revealed by Electron Cryotomography. Biol. Reprod. 2012, 87, 73. [Google Scholar] [CrossRef] [PubMed]
- Baccetti, B. Genetic Sperm Defects and Consanguinity. Hum. Reprod. 2001, 16, 1365–1371. [Google Scholar] [CrossRef] [Green Version]
- Perotti, M.E.; Giarola, A.; Gioria, M. Ultrastructural Study of the Decapitated Sperm Defect in an Infertile Man. J. Reprod. Fertil. 1981, 63, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Baccetti, B.; Selmi, M.G.; Soldani, P. Morphogenesis of “decapitated” Spermatozoa in a Man. Reproduction 1984, 70, 395–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chemes, H.; Carizza, C.; Scarinci, F. Lack of a Head in Human Spermatozoa from Sterile Patients: A Syndrome Associated with Impaired Fertilization. Int. J. Gynecol. Obs. 1988, 26, 165. [Google Scholar] [CrossRef]
- Baccetti, B.; Burrini, A.G.; Collodel, G.; Magnano, A.R.; Piomboni, P.; Renieri, T.; Sensini, C. Morphogenesis of the Decapitated and Decaudated Sperm Defect in Two Brothers. Gamete Res. 1989, 23, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Chemes, H.E.; Puigdomenech, E.T.; Carizza, C.; Olmedo, S.B.; Zanchetti, F.; Hermes, R. Acephalic Spermatozoa and Abnormal Development of the Head–Neck Attachment: A Human Syndrome of Genetic Origin. Hum. Reprod. 1999, 14, 1811–1818. [Google Scholar] [CrossRef] [Green Version]
- Toyama, Y.; Iwamoto, T.; Yajima, M.; Baba, K.; Yuasa, S. Decapitated and Decaudated Spermatozoa in Man, and Pathogenesis Based on the Ultrastructure. Int. J. 2000, 23, 109–115. [Google Scholar] [CrossRef]
- Holstein, A.F.; Schill, W.B.; Breucker, H. Dissociated Centriole Development as a Cause of Spermatid Malformation in Man. J. Reprod. Fertil. 1986, 78, 719–725. [Google Scholar] [CrossRef] [Green Version]
- Schalles, U.; Shao, X.; van der Hoorn, F.A.; Oko, R. Developmental Expression of the 84-KDa ODF Sperm Protein: Localization to Both the Cortex and Medulla of Outer Dense Fibers and to the Connecting Piece. Dev. Biol. 1998, 199, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Meinhardt, A.; Zhang, B.; Grzmil, P.; Adham, I.M.; Hoyer-Fender, S. The Small Heat Shock Protein ODF1/HSPB10 Is Essential for Tight Linkage of Sperm Head to Tail and Male Fertility in Mice. Mol. Cell. Biol. 2012, 32, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Grzmil, P.; Meinhardt, A.; Hoyer-Fender, S. Haplo-Deficiency of ODF1/HSPB10 in Mouse Sperm Causes Relaxation of Head-to-Tail Linkage. Reproduction 2014, 148, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Hetherington, L.; Schneider, E.K.; Scott, C.; DeKretser, D.; Muller, C.H.; Hondermarck, H.; Velkov, T.; Baker, M.A. Deficiency in Outer Dense Fiber 1 Is a Marker and Potential Driver of Idiopathic Male Infertility. Mol. Cell. Proteom. Mcp 2016, 15, 3685–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia Contreras, C.; Hoyer-Fender, S. CCDC42 Localizes to Manchette, HTCA and Tail and Interacts With ODF1 and ODF2 in the Formation of the Male Germ Cell Cytoskeleton. Front. Cell Dev. Biol. 2019, 7, 151. [Google Scholar] [CrossRef] [Green Version]
- Brohmann, H.; Pinnecke, S.; Hoyer-Fender, S. Identification and Characterization of New cDNAs Encoding Outer Dense Fiber Proteins of Rat Sperm. J. Biol. Chem. 1997, 272, 10327–10332. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C. Outer Dense Fibre Proteins from Human Sperm Tail: Molecular Cloning and Expression Analyses of Two cDNA Transcripts Encoding Proteins of 70 kDa. Mol. Hum. Reprod. 1999, 5, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Salmon, N.A.; Reijo Pera, R.A.; Xu, E.Y. A Gene Trap Knockout of the Abundant Sperm Tail Protein, Outer Dense Fiber 2, Results in Preimplantation Lethality. Genesis 2006, 44, 515–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarnasky, H.; Cheng, M.; Ou, Y.; Thundathil, J.C.; Oko, R.; van der Hoorn, F.A. Gene Trap Mutation of Murine Outer Dense Fiber Protein-2 Gene Can Result in Sperm Tail Abnormalities in Mice with High Percentage Chimaerism. Bmc Dev. Biol. 2010, 10, 67. [Google Scholar] [CrossRef] [Green Version]
- Ito, C.; Akutsu, H.; Yao, R.; Yoshida, K.; Yamatoya, K.; Mutoh, T.; Makino, T.; Aoyama, K.; Ishikawa, H.; Kunimoto, K.; et al. Odf2 Haploinsufficiency Causes a New Type of Decapitated and Decaudated Spermatozoa, Odf2-DDS, in Mice. Sci. Rep. 2019, 9, 14249. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C.; Aumüller, G.; Bahrami, M.; Hoyer-Fender, S. Molecular Cloning of Odf3 Encoding a Novel Coiled-Coil Protein of Sperm Tail Outer Dense Fibers. Mol. Reprod. Dev. 2002, 61, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Liu, C.; Wang, F.; Yang, X.; Zhang, J.; Wu, H.; Zhang, Z.; He, X.; Zhang, Z.; Zhou, P.; et al. Mutations in PMFBP1 Cause Acephalic Spermatozoa Syndrome. Am. J. Hum. Genet. 2018, 103, 188–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sha, Y.-W.; Wang, X.; Xu, X.; Ding, L.; Liu, W.-S.; Li, P.; Su, Z.-Y.; Chen, J.; Mei, L.-B.; Zheng, L.-K.; et al. Biallelic Mutations in PMFBP1 Cause Acephalic Spermatozoa. Clin. Genet. 2019, 95, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wang, N.; Zhang, H.; Yin, S.; Dai, H.; Lin, G.; Li, W. Novel Mutations in PMFBP1, TSGA10 and SUN5: Expanding the Spectrum of Mutations That May Cause Acephalic Spermatozoa. Clin. Genet. 2020, 97, 938–939. [Google Scholar] [CrossRef]
- Pasek, R.C.; Malarkey, E.; Berbari, N.F.; Sharma, N.; Kesterson, R.A.; Tres, L.L.; Kierszenbaum, A.L.; Yoder, B.K. Coiled-Coil Domain Containing 42 (Ccdc 42) Is Necessary for Proper Sperm Development and Male Fertility in the Mouse. Dev. Biol. 2016, 412, 208–218. [Google Scholar] [CrossRef]
- Shao, X.; Tarnasky, H.A.; Lee, J.P.; Oko, R.; van der Hoorn, F.A. Spag4, a Novel Sperm Protein, Binds Outer Dense-Fiber Protein Odf1 and Localizes to Microtubules of Manchette and Axoneme. Dev. Biol. 1999, 211, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Pasch, E.; Link, J.; Beck, C.; Scheuerle, S.; Alsheimer, M. The LINC Complex Component Sun4 Plays a Crucial Role in Sperm Head Formation and Fertility. Biol. Open 2015, 4, 1792–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvi, A.; Wong, A.S.W.; Wright, G.; Wong, E.S.M.; Loo, T.H.; Stewart, C.L.; Burke, B. SUN4 Is Essential for Nuclear Remodeling during Mammalian Spermiogenesis. Dev. Biol. 2015, 407, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Adham, I.M.; Meinhardt, A.; Hoyer-Fender, S. Ultra-Structure of the Sperm Head-to-Tail Linkage Complex in the Absence of the Spermatid-Specific LINC Component SPAG4. Histochem. Cell Biol. 2018, 150, 49–59. [Google Scholar] [CrossRef]
- Shang, Y.; Zhu, F.; Wang, L.; Ouyang, Y.-C.; Dong, M.-Z.; Liu, C.; Zhao, H.; Cui, X.; Ma, D.; Zhang, Z.; et al. Essential Role for SUN5 in Anchoring Sperm Head to the Tail. eLife 2017, 6, e28199. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Wang, F.; Yang, X.; Zhang, J.; Wu, H.; Zhang, Z.; Zhang, Z.; He, X.; Zhou, P.; Wei, Z.; et al. Biallelic SUN5 Mutations Cause Autosomal-Recessive Acephalic Spermatozoa Syndrome. Am. J. Hum. Genet. 2016, 99, 942–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkhatib, R.A.; Paci, M.; Longepied, G.; Saias-Magnan, J.; Courbiere, B.; Guichaoua, M.-R.; Lévy, N.; Metzler-Guillemain, C.; Mitchell, M. Homozygous Deletion of SUN5 in Three Men with Decapitated Spermatozoa. Hum. Mol. Genet. 2017, 26, 3167–3171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.; Yan, J.; Tang, W.; Liu, C.; Xiao, S.; Guo, Y.; Yuan, L.; Chen, L.; Jiang, H.; Guo, X.; et al. Mechanistic Insights into Acephalic Spermatozoa Syndrome–Associated Mutations in the Human SUN5 Gene. J. Biol. Chem. 2018, 293, 2395–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yang, L.; Huang, L.; Liu, G.; Nie, X.; Zhang, X.; Xing, X. SUN5 Interacting With Nesprin3 Plays an Essential Role in Sperm Head-to-Tail Linkage: Research on Sun5 Gene Knockout Mice. Front. Cell Dev. Biol. 2021, 9, 684826. [Google Scholar] [CrossRef] [PubMed]
- Tokuhiro, K.; Isotani, A.; Yokota, S.; Yano, Y.; Oshio, S.; Hirose, M.; Wada, M.; Fujita, K.; Ogawa, Y.; Okabe, M.; et al. OAZ-t/OAZ3 Is Essential for Rigid Connection of Sperm Tails to Heads in Mouse. PLoS Genet. 2009, 5, e1000712. [Google Scholar] [CrossRef] [Green Version]
- Goto, M.; Eddy, E.M. Speriolin Is a Novel Spermatogenic Cell-Specific Centrosomal Protein Associated with the Seventh WD Motif of Cdc20. J. Biol. Chem. 2004, 279, 42128–42138. [Google Scholar] [CrossRef] [Green Version]
- Goto, M.; O’Brien, D.A.; Eddy, E.M. Speriolin Is a Novel Human and Mouse Sperm Centrosome Protein. Hum. Reprod. 2010, 25, 1884–1894. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kwon, J.T.; Jeong, J.; Kim, J.; Hong, S.H.; Kim, J.; Park, Z.Y.; Chung, K.H.; Eddy, E.M.; Cho, C. SPATC1L Maintains the Integrity of the Sperm Head-tail Junction. Embo Rep. 2018, 19, e45991. [Google Scholar] [CrossRef]
- Li, Y.-Z.; Li, N.; Liu, W.-S.; Sha, Y.-W.; Wu, R.-F.; Tang, Y.-L.; Zhu, X.-S.; Wei, X.-L.; Zhang, X.-Y.; Wang, Y.-F.; et al. Biallelic Mutations in Spermatogenesis and Centriole-Associated 1 like (SPATC1L) Cause Acephalic Spermatozoa Syndrome and Male Infertility. Asian J. 2021. [Google Scholar] [CrossRef]
- Yuan, S.; Stratton, C.J.; Bao, J.; Zheng, H.; Bhetwal, B.P.; Yanagimachi, R.; Yan, W. Spata6 Is Required for Normal Assembly of the Sperm Connecting Piece and Tight Head–Tail Conjunction. Proc. Natl. Acad. Sci. USA 2015, 112, E430–E439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netzel-Arnett, S.; Bugge, T.H.; Hess, R.A.; Carnes, K.; Stringer, B.W.; Scarman, A.L.; Hooper, J.D.; Tonks, I.D.; Kay, G.F.; Antalis, T.M. The Glycosylphosphatidylinositol-Anchored Serine Protease PRSS21 (Testisin) Imparts Murine Epididymal Sperm Cell Maturation and Fertilizing Ability1. Biol. Reprod. 2009, 81, 921–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapiro, R.; Kostetskii, I.; Olds-Clarke, P.; Gerton, G.L.; Radice, G.L.; Strauss III, J.F. Male Infertility, Impaired Sperm Motility, and Hydrocephalus in Mice Deficient in Sperm-Associated Antigen 6. Mol. Cell. Biol. 2002, 22, 6298–6305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, L.; Li, W.; Huang, Q.; Yuan, S.; Li, Y.; Liu, J.; Zhang, S.; Pin, G.; Song, S.; et al. The Sperm-Associated Antigen 6 Interactome and Its Role in Spermatogenesis. Reproduction 2019, 158, 183–199. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Lujambio, I. The Hook1 Gene Is Non-Functional in the Abnormal Spermatozoon Head Shape (Azh) Mutant Mouse. Hum. Mol. Genet. 2002, 11, 1647–1658. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhu, Y.; Zhu, Z.; Zhi, E.; Lu, K.; Wang, X.; Liu, F.; Li, Z.; Xia, W. Detection of Heterozygous Mutation in Hook Microtubule-Tethering Protein 1 in Three Patients with Decapitated and Decaudated Spermatozoa Syndrome. J. Med. Genet. 2018, 3, 150–157. [Google Scholar] [CrossRef]
- Zhou, J.; Du, Y.-R.; Qin, W.-H.; Hu, Y.-G.; Huang, Y.-N.; Bao, L.; Han, D.; Mansouri, A.; Xu, G.-L. RIM-BP3 Is a Manchette-Associated Protein Essential for Spermiogenesis. Development 2009, 136, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Ouyang, Y.-C.; Jiang, B.; Lin, X.; Chen, J.; Dong, M.-Z.; Zhuang, X.; Yuan, S.; Sun, Q.-Y.; Han, C. Non-Canonical RNA Polyadenylation Polymerase FAM46C Is Essential for Fastening Sperm Head and Flagellum in Mice. Biol. Reprod. 2019, 100, 1673–1685. [Google Scholar] [CrossRef]
- Liška, F.; Gosele, C.; Rivkin, E.; Tres, L.; Cardoso, M.C.; Domaing, P.; Krejčí, E.; Šnajdr, P.; Lee-Kirsch, M.A.; de Rooij, D.G.; et al. Rat Hd Mutation Reveals an Essential Role of Centrobin in Spermatid Head Shaping and Assembly of the Head-Tail Coupling Apparatus1. Biol. Reprod. 2009, 81, 1196–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liška, F.; Gosele, C.; Popova, E.; Chylíková, B.; Křenová, D.; Křen, V.; Bader, M.; Tres, L.L.; Hubner, N.; Kierszenbaum, A.L. Overexpression of Full-Length Centrobin Rescues Limb Malformation but Not Male Fertility of the Hypodactylous (Hd) Rats. PLoS ONE 2013, 8, e60859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Wei, X.; Sha, Y.; Li, N.; Yan, X.; Cheng, L.; Qiao, D.; Zhou, W.; Wu, R.; Liu, Q.; et al. Loss-of-Function Mutation in TSGA10 Causes Acephalic Spermatozoa Phenotype in Human. Mol. Genet. Genom. Med. 2020, 8, e1284. [Google Scholar] [CrossRef]
- Luo, G.; Hou, M.; Wang, B.; Liu, Z.; Liu, W.; Han, T.; Zhang, D.; Zhou, X.; Jia, W.; Tan, Y.; et al. Tsga10 Is Essential for Arrangement of Mitochondrial Sheath and Male Fertility in Mice. Andrology 2021, 9, 368–375. [Google Scholar] [CrossRef]
- Mahsa Mobahat, B.B. TSGA10 Is a Centrosomal Protein, Interacts with ODF2 and Localizes to Basal Body. J. Cell Sci. 2015. [Google Scholar] [CrossRef]
- Nozawa, Y.I.; Yao, E.; Gacayan, R.; Xu, S.-M.; Chuang, P.-T. Mammalian Fused Is Essential for Sperm Head Shaping and Periaxonemal Structure Formation during Spermatogenesis. Dev. Biol. 2014, 388, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Jiang, M.; Yuan, Y.; Bi, Y.; Zheng, B.; Guo, X.; Huang, X.; Zhou, Z.; Sha, J. ADP-Ribosylation Factor-like 3, a Manchette-Associated Protein, Is Essential for Mouse Spermiogenesis. Mol. Hum. Reprod. 2013, 19, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wolgemuth, D.J. BET Protein BRDT Complexes With HDAC1, PRMT5, and TRIM28 and Functions in Transcriptional Repression During Spermatogenesis. J. Cell. Biochem. 2016, 117, 1429–1438. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Sha, Y.; Wang, X.; Li, P.; Wang, J.; Kee, K.; Wang, B. Whole-Exome Sequencing Identified a Homozygous BRDT Mutation in a Patient with Acephalic Spermatozoa. Oncotarget 2017, 8, 19914–19922. [Google Scholar] [CrossRef]
- Sha, Y.; Wang, X.; Yuan, J.; Zhu, X.; Su, Z.; Zhang, X.; Xu, X.; Wei, X. Loss-of-Function Mutations in Centrosomal Protein 112 Is Associated with Human Acephalic Spermatozoa Phenotype. Clin. Genet. 2020, 97, 321–328. [Google Scholar] [CrossRef]
- Bedford, J.M.; Calvin, H.I. Changes in -S-S- Linked Structures of the Sperm Tail during Epididymal Maturation, with Comparative Observations in Sub-Mammalian Species. J. Exp. Zool. 1974, 187, 181–203. [Google Scholar] [CrossRef]
- Young, R.J.; Cooper, G.W. Dissociation of Intermolecular Linkages of the Sperm Head and Tail by Primary Amines, Aldehydes, Sulphydryl Reagents and Detergents. Reproduction 1983, 69, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Calvin, H.I.; Hwang, F.H.; Wohlrab, H. Localization of Zinc in a Dense Fiber-Connecting Piece Fraction of Rat Sperm Tails Analogous Chemically to Hair Keratin. Biol. Reprod. 1975, 13, 228–239. [Google Scholar] [CrossRef] [Green Version]
- Björndahl, L.; Kvist, U. Importance of Zinc for Human Sperm Head-Tail Connection. Acta Physiol. Scand. 1982, 116, 51–55. [Google Scholar] [CrossRef]
- Burfeind, P.; Hoyer-Fender, S. Sequence and Developmental Expression of a mRNA Encoding a Putative Protein of Rat Sperm Outer Dense Fibers. Dev. Biol. 1991, 148, 195–204. [Google Scholar] [CrossRef]
- Gastmann, O.; Burfeind, P.; Günther, E.; Hameister, H.; Szpirer, C.; Hoyer-Fender, S. Sequence, Expression, and Chromosomal Assignment of a Human Sperm Outer Dense Fiber Gene. Mol. Reprod. Dev. 1993, 36, 407–418. [Google Scholar] [CrossRef]
- Morales, C.R.; Oko, R.; Clermont, Y. Molecular Cloning and Developmental Expression of an mRNA Encoding the 27 kDa Outer Dense Fiber Protein of Rat Spermatozoa. Mol. Reprod. Dev. 1994, 37, 229–240. [Google Scholar] [CrossRef]
- Starr, D.A.; Fridolfsson, H.N. Interactions Between Nuclei and the Cytoskeleton Are Mediated by SUN-KASH Nuclear-Envelope Bridges. Annu. Rev. Cell Dev. Biol. 2010, 26, 421–444. [Google Scholar] [CrossRef] [Green Version]
- Göb, E.; Schmitt, J.; Benavente, R.; Alsheimer, M. Mammalian Sperm Head Formation Involves Different Polarization of Two Novel LINC Complexes. PLoS ONE 2010, 5, e12072. [Google Scholar] [CrossRef] [Green Version]
- Frohnert, C.; Schweizer, S.; Hoyer-Fender, S. SPAG4L/SPAG4L-2 Are Testis-Specific SUN Domain Proteins Restricted to the Apical Nuclear Envelope of Round Spermatids Facing the Acrosome. Mol. Hum. Reprod. 2011, 17, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Kmonickova, V.; Frolikova, M.; Steger, K.; Komrskova, K. The Role of the LINC Complex in Sperm Development and Function. Int. J. Mol. Sci. 2020, 21, E9058. [Google Scholar] [CrossRef] [PubMed]
- Manfrevola, F.; Guillou, F.; Fasano, S.; Pierantoni, R.; Chianese, R. LINCking the Nuclear Envelope to Sperm Architecture. Genes 2021, 12, 658. [Google Scholar] [CrossRef] [PubMed]
- Lehti, M.S.; Sironen, A. Formation and Function of the Manchette and Flagellum during Spermatogenesis. Reproduction 2016, 151, R43–R54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olenick, M.A.; Holzbaur, E.L.F. Dynein Activators and Adaptors at a Glance. J. Cell Sci. 2019, 132, jcs227132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szebenyi, G.; Hall, B.; Yu, R.; Hashim, A.I.; Krämer, H. Hook2 Localizes to the Centrosome, Binds Directly to Centriolin/CEP110 and Contributes to Centrosomal Function. Traffic 2007, 8, 32–46. [Google Scholar] [CrossRef]
- Manandhar, G.; Simerly, C.; Salisbury, J.L.; Schatten, G. Centriole and Centrin Degeneration during Mouse Spermiogenesis. Cell Motil. Cytoskelet. 1999, 43, 137–144. [Google Scholar] [CrossRef]
- Manandhar, G.; Schatten, H.; Sutovsky, P. Centrosome Reduction During Gametogenesis and Its Significance. Biol. Reprod. 2005, 72, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Avasthi, P.; Scheel, J.F.; Ying, G.; Frederick, J.M.; Baehr, W.; Wolfrum, U. Germline Deletion of Cetn1 Causes Infertility in Male Mice. J. Cell Sci. 2013, 126, 3204–3213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firat-Karalar, E.N.; Sante, J.; Elliott, S.; Stearns, T. Proteomic Analysis of Mammalian Sperm Cells Identifies New Components of the Centrosome. J. Cell Sci. 2014, 127, 4128–4133. [Google Scholar] [CrossRef] [Green Version]
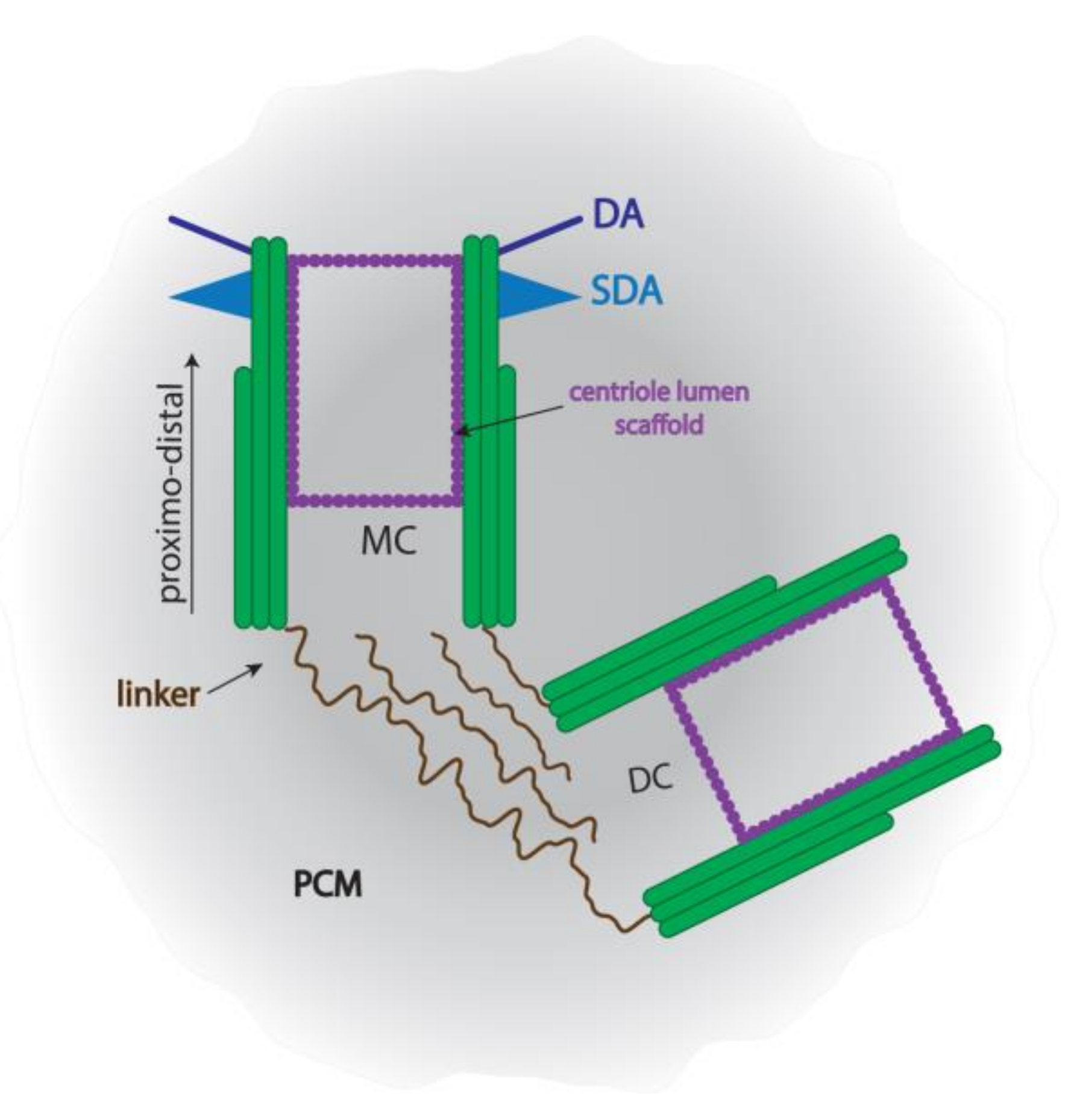
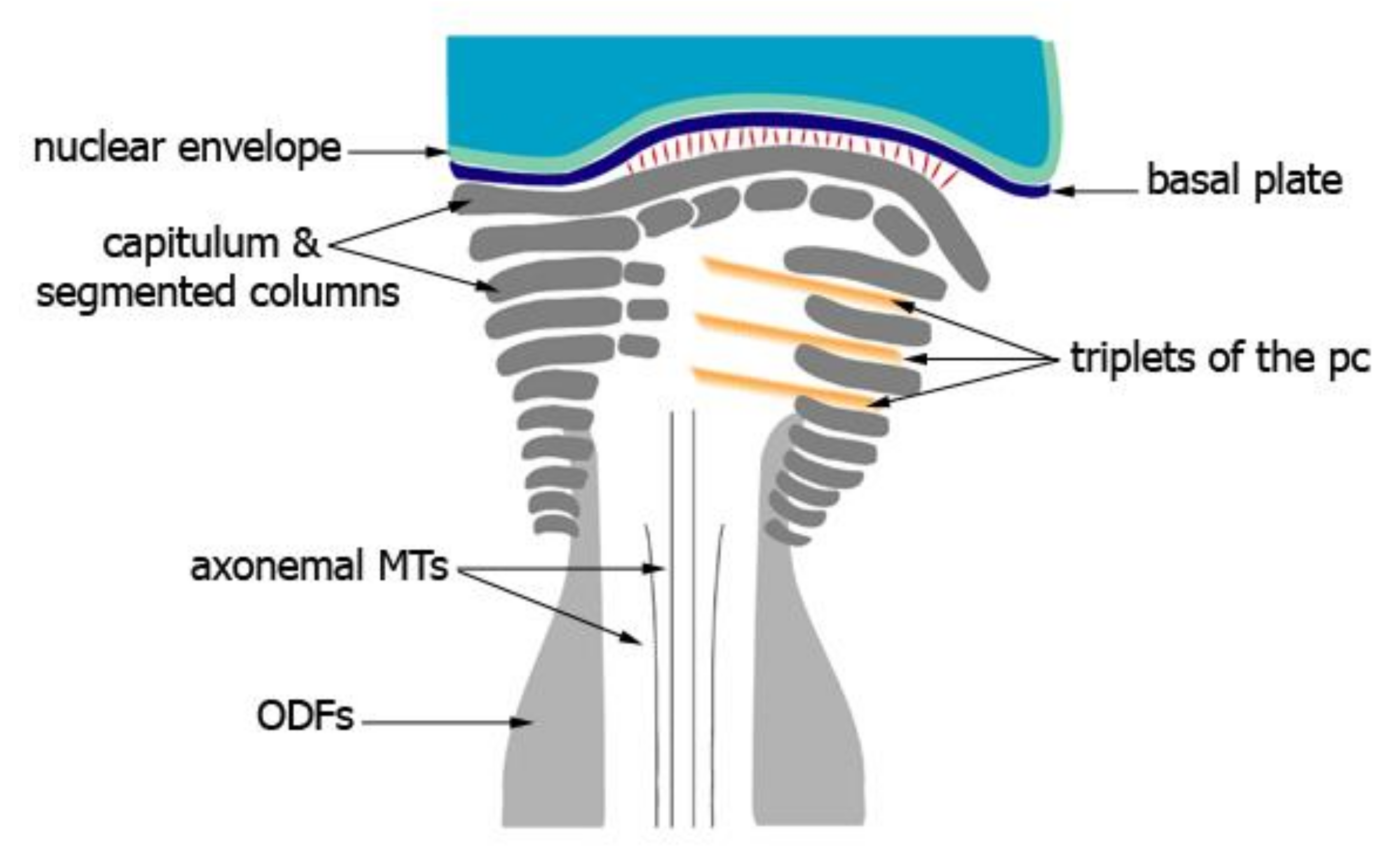

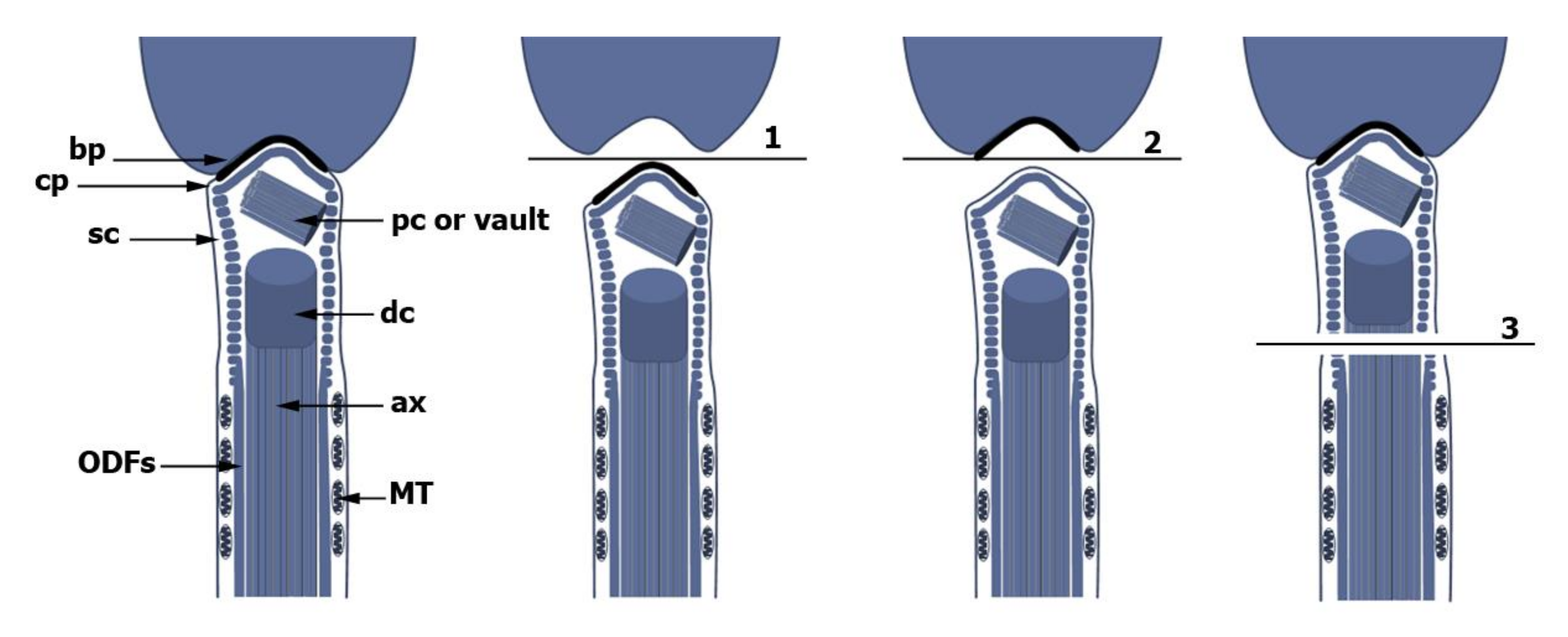

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Protein | Function |
|---|---|---|
| PCM Formation | SAS4/CPAP | Centriole assembly |
| CEP135 | Centriole assembly | |
| CEP295 | PCM assembly | |
| CEP152/asterless/Asl | PLK4 recruitment | |
| CEP192 | PLK4 recruitment, PCM assembly | |
| NEDD1 | Adapter of CEP192, binds to γ-TuRCs | |
| CDK5RAP2/CEP215 | PCM, binds to γ-TuRCs | |
| PCNT/pericentrin | PCM, binds to γ-TuRCs | |
| CEP63 | Binds CEP152 and CEP57 | |
| CEP57 | Anchors CEP63/CEP152 complex to the centriole | |
| PCM-1 | Centriolar satellites | |
| Centriole Lumen | POC1B | |
| Scaffold | POC5 | |
| CETN/centrin | ||
| FAM161A | ||
| WDR90 | ||
| Centriole Duplication | CEP192 | PLK4 recruitment |
| CEP152 | PLK4 recruitment, centriole length control | |
| PLK4 | ||
| STIL | Procentriole formation, cartwheel | |
| SAS6 | Procentriole formation, cartwheel | |
| CEP135 | Procentriole formation | |
| CPAP/CENPJ/SAS4 | Procentriole formation, centriole length control | |
| Procentriole elongation | ||
| CNTROB/centrobin | Centriole length control | |
| CEP120/CCDC100 | Centriole length control | |
| SPICE1/CCDC52 | Centriole length control | |
| POC1A, POC1B | Centriole length control | |
| POC5/C50rf37 | Centriole length control | |
| CEP295 | Centriole length control | |
| CP110 | Centriole length control | |
| CEP97 | Regulation of centriole duplication | |
| CEP76 | Centriole duplication | |
| CEP72 | ||
| CEP63 | ||
| CEP57/translokin | ||
| CCDC57 | ||
| WDR62 | Centriole duplication, centriolar satellites | |
| CEP131/AZI1 | Centriole length | |
| RTTN, PPP1R35, C2CD3 | Centriole stability | |
| ε-Tubulin, ό-tubuline, TEDC1, TEDC2 | ||
| RTTN/rotatin | ||
| Linker at the Proximal Ends of MC and DC | Rootletin | |
| C-NAP1/CEP250 | ||
| CNTLN/centlein | ||
| CEP68 | ||
| CG-NAP | ||
| LRRC45 | ||
| β-Catenin | ||
| Distal Centriole Ends | OFD1 | Centriole stabilization |
| CP110 | ||
| CEP97 | ||
| CEP78 | ||
| CETN | ||
| CEP19 | Distal ends of mother centriole | |
| CEP350 | Distal ends of mother centriole | |
| C2CD3 | Distal ends of mother centriole | |
| CEP43, CBY1/Chibby | Distal ends of mother centriole, centriolar satellites | |
| Daughter Centriole | CNTROB | |
| CEP120 | ||
| STIL | ||
| Distal Appendages | CEP164 | Ciliogenesis |
| of Mother Centriole | CEP89/CEP123/CCDC123 | |
| CEP83/CCDC41 | ||
| FBF1 | ||
| SCLT1 | ||
| C2CD3 | ||
| OFD1 | ||
| Subdistal Appendages of Mother Centriole | ODF2/cenexin | MT anchoring |
| TCHP/trichoplein | ||
| NIN/ninein | ||
| CEP170/FAM68A | ||
| CEP128 | ||
| CEP110/Centriolin/CEP1/CNTRL | ||
| CCDC120 | ||
| CCDC68 | ||
| CCDC61/hVFL3 | ||
| Nde1 | ||
| ε-Tubulin/TUBE1 | ||
| CC2D2A |
| Protein | Centrosomal Localization and Function | Localization in Spermatozoa | Phenotype of Knockout Animals | Mutations in Humans | References |
|---|---|---|---|---|---|
| ODF1 | Centrosome in NIH3T3 cells. | Manchette, HTCA, and flagellum/ODFs. ODFs, segmented columns, capitulum, and basal plate by IEM. Interacts with ODF2, CCDC42. | Odf1 ko by HR. Male mice are infertile with disorganized sperm tails and sperm decapitation. Weakened head to tail linkage in the heterozygous condition. | Easy sperm decapitation in infertile men with a reduction in ODF1 protein. | [144,145,146,147,148] |
| ODF2 | PCM, mother centriole, SDAs. Essential for primary cilia formation; centriole cohesion. Interacts with ODF1, CCDC42, CEP128, ß-catenin, trichoplein. | Flagellum/ODFs. ODFs, segmented columns, capitulum, and basal plate by IEM. | Chimaeric male mice of high percentage chimaerism, generated of Odf2 XL169 ES cells, in which the ß-neo gene-trap cassette is inserted into the Odf2 gene, are infertile. Infertility due to bent tails, missing or thinning of individual ODFs, absence of one or more axonemal MT doublets. | [101,102,103,112,149,150,151,152,153] | |
| ODF3/PMFBP1 | Flagellum/ODFs, HTCA. Cooperates with SUN5 and SPATA6. | Crispr/Cas-mediated ko. Male mice are infertile. Acephalic spermatozoa. Basal plate attached to the tail, not the nucleus. | Mutations reported in consanguineous families with ASS. | [154,155,156,157] | |
| CCDC42 | Centrosome and the basal body in somatic cells. | Manchette, connecting piece and tail. Interacts with ODF1 and ODF2. | Ccdc42 ko mice by HR (null allele). Male sterility. Elongated spermatids with abnormally shaped heads, dislocation of the HTCA from the implantation fossa, and multiplicity of HTCAs. No MT axonemes observed from the HTCA. | [148,158] | |
| SUN4/SPAG4 | SUN-domain-containing protein, testis-specific, nuclear envelope. | Male infertility in Sun4-deficient mice. Defects in sperm head formation. Required for tight head-to-tail linkage in sperm, but HTCA is still present. | [159,160,161,162] | ||
| SUN5/ SPAG4L/4L-2 | SUN-domain-containing protein, testis-specific, nuclear envelope. | Interacts with nesprin3. | TALEN-generated Sun5 ko mice by targeting exon 4). Male infertility with decapitated spermatozoa. Basal plate associated with the capitulum. | Approximately 33-47% of patients with ASS have mutations in SUN5. | [163,164,165,166,167] |
| OAZ-t/OAZ3 | Specifically expressed in spermatids. | Homozygous mutant males are infertile due to easy sperm decapitation. Basal plate remains associated with the nucleus. | [168] | ||
| Speriolin (Sprn, Spatc1, Spata15) | Concentrated at the centrosome of spermatocytes and spermatids. Localized in the neck region: surrounds the intact proximal centriole in human sperm, and the periphery of the dis-ordered distal centriole in mouse sperm. | Not reported | Not reported | [169,170] | |
| Speriolin-like protein (Spatc1l, C21orf56) | Expression starts in the round spermatid stage in mice. Localized at the neck region in testicular sperm of mice. Associates with the regulatory subunit of cAMP-dependent protein kinase A (PKA). Interacts with capping protein muscle Z-line beta (CAPZB). | Crispr/Cas-mediated ko mice. Male sterility. Sperm decapitation. Separation between the basal plate and the capitulum. | [171,172] | ||
| SPATA6/SRF1/HASH | Striated columns, capitulum | Ko mice are male sterile. Acephalic spermatozoa, impaired development of the connecting piece (by TEM). pc present but segmented columns, capitulum and mitochondrial sheath are lacking. Disturbance of the tail structure (ODFs, MT). | [173] | ||
| PRSS21 /testisin | Targeted disruption of the coding sequence by HR. PRSS21 deficient spermatozoa show decreased motility, angulated and curled tails, fragile necks and easily decapitated spermatozoa. | [174] | |||
| SPAG6/PF16/CFAP194 | Component of the central apparatus of the 9+2 axoneme. | Spag6 ko by HR. Spag6-deficient testes showed abnormal spermatogenesis with abnormalities in male germ cell morphology. Disorganized flagellar structures and frequent loss of the sperm head. Spag6-deficient mice have further cilia-related disorders (i.a. hydrocephalus). | [175,176] | ||
| HOOK1 | Centrosome in NIH3T3 cells. | Predominantly expressed in haploid male germ cells. Interacts with CCDC81, and ODF2. | HOOK1 mutation causative for the azh phenotype in mice, with head and tail abnormalities, and sperm decapitation. | Whole genome sequencing of patients with decapitated and decaudated spermatozoa revealed mutations in HOOK1. | [177,178] |
| RIM-BP3 | RIM-BPs as adaptors in the process of vesicle fusion and release. | Manchette associated, interacts with HOOK1. Expressed almost exclusively in the testis. | Targeted deletion in mice revealed abnormal sperm heads and tail detachment. | [179] | |
| FAM46C | Specifically located to the manchette of mouse spermatids. | Crispr/Cas-mediated ko mice. Male sterility with headless spermatozoa. In step13-16 spermatids, abnormal connecting pieces with incomplete segmented columns and capitulum. | [180] | ||
| Centrobin/ CNTROB/NIP2 | Centriole duplication and stability | Capitulum, dc and pc. Localizes to the manchette, centrosome, and marginal ring of the acroplaxome. | Truncated CNTROB protein by insertion of a retrovirus in intron 10 of the gene in the hd rat (hypodactylous). Disruption of the HTCA and sperm decapitation. A full- length cDNA in testis did rescue limb malformation but not decapitation. CNTROB might be required for the stabilisation of the attachment of the sperm head to the flagellum. | [89,181,182] | |
| TSGA10 (CEP4L, Cancer/testis antigen 79, CT79, SPGF26) | Crispr/Cas-mediated ko mice. Heterozygous male mice are infertile with reduced sperm count and disordered mitochondrial sheath formation. | Whole exome sequencing revealed mutations in ASS patients. | [157,183,184,185] | ||
| Fu/fused | Serine-threonine kinase | Interacts with ODF1, and KIF27. | Conditional inactivation caused sperm decapitation, i.a. | [186] | |
| ADP-ribosylation factor-like 3 (ARL3) | Ras-related small GTP-binding protein. | Manchette. | siRNA injection into testis provoked abnormal head shape, lasso-like coiled tails, and sperm decapitation. | [187] | |
| BRDT (cancer/testis antigen 9, RING3-like protein, BRD6, SPGF21) | BRDT (Bromo-domain testis associated) involved in gene repression. | BRDT expression restricted to male germ cells, specifically to pachytene/diplotene spermatocytes and early spermatids. | Loss of the first bromodomain of BRDT by targeted mutagenesis resulted in sterility and abnormalities in spermiogenesis. | Mutations in male patients with ASS. | [188,189] |
| CEP112/ CCDC46 | Not reported. | Mutations found in male patients with ASS by whole exome sequencing. | [190] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tapia Contreras, C.; Hoyer-Fender, S. The Transformation of the Centrosome into the Basal Body: Similarities and Dissimilarities between Somatic and Male Germ Cells and Their Relevance for Male Fertility. Cells 2021, 10, 2266. https://doi.org/10.3390/cells10092266
Tapia Contreras C, Hoyer-Fender S. The Transformation of the Centrosome into the Basal Body: Similarities and Dissimilarities between Somatic and Male Germ Cells and Their Relevance for Male Fertility. Cells. 2021; 10(9):2266. https://doi.org/10.3390/cells10092266
Chicago/Turabian StyleTapia Contreras, Constanza, and Sigrid Hoyer-Fender. 2021. "The Transformation of the Centrosome into the Basal Body: Similarities and Dissimilarities between Somatic and Male Germ Cells and Their Relevance for Male Fertility" Cells 10, no. 9: 2266. https://doi.org/10.3390/cells10092266